
Citrullus colocynthis Seeds: A Potential Natural Immune Modulator Source for Broiler Reared under Chronic Heat Stress


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Compliance Statement
2.2. Preparation of CC Seeds Powder
2.3. Birds’ Management and Experimental Design
2.4. Production Performance Traits
2.5. Blood Sampling and Preparation
2.6. Stress Indicators and Antioxidant Markers
2.6.1. H/L Ratio
2.6.2. Plasma Corticosterone
2.6.3. MDA and TNF-α Levels in the PBMCs
2.6.4. TAC and SOD Levels in the PBMCs
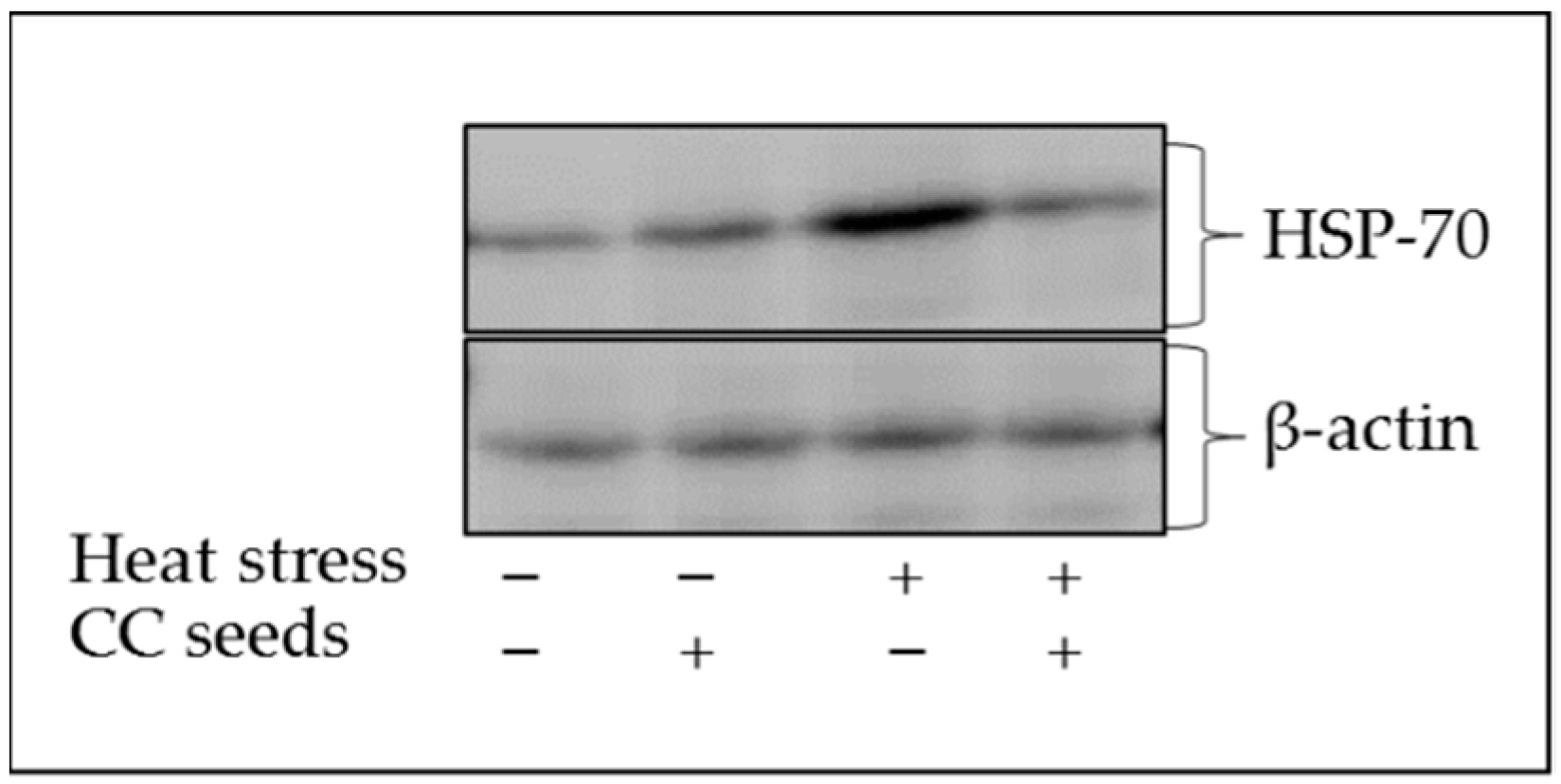
2.7. Expression of HSP-70 in PBMCs
2.8. Immunological Parameters
2.8.1. TWBCs Count
2.8.2. Toe Web Swelling
2.8.3. Antibody Titers against Sheep Red Blood Cells (Anti-SRBCs AB)
2.8.4. Lymphocyte Proliferation Index
2.9. Plasma Biochemical Assay
2.10. Statistical Analysis
3. Results
3.1. Production Performance
3.2. Stress Indicators and Antioxidant Markers
3.3. Immunological Parameters
3.4. Plasma Biochemical Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nagai, M.; Iriki, M. Changes in immune activities by heat stress. In Thermotherapy for Neoplasia, Inflammation, and Pain; Kosaka, M., Sugahara, T., Schmidt, K.L., Simon, E., Eds.; Springer: Tokyo, Japan, 2001; pp. 266–270. [Google Scholar]
- Xu, Y.; Lai, X.; Li, Z.; Zhang, X.; Luo, Q. Effect of chronic heat stress on some physiological and immunological parameters in different breed of broilers. Poult. Sci. 2018, 97, 4073–4082. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Liu, Q.; Zhang, M.; Feng, J.; Li, X.; Zhou, Y.; Wang, X. iTRAQ-based quantitative proteomics analysis of the spleen reveals innate immunity and cell death pathways associated with heat stress in broilers (Gallus gallus). J. Proteom. 2019, 196, 11–21. [Google Scholar] [CrossRef]
- Hosseini-Vashan, S.J.; Raei-Moghadam, M.S. Antioxidant and immune system status, plasma lipid, abdominal fat, and growth performance of broilers exposed to heat stress and fed diets supplemented with pomegranate pulp (Punica granatum L.). J. Appl. Anim. Res. 2019, 47, 521–531. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Yin, Q.; Xiong, Y.; Liu, D.; Hu, H. Effects of dietary fumaric acid on the growth performance, immune response, relative weight and antioxidant status of immune organs in broilers exposed to chronic heat stress. Czech J. Anim. Sci. 2020, 65, 104–113. [Google Scholar] [CrossRef]
- Awad, E.A.; Najaa, M.; Zulaikha, Z.A.; Zulkifli, I.; Soleimani, A.F. Effects of heat stress on growth performance, selected physiological and immunological parameters, caecal microflora, and meat quality in two broiler strains. Asian-Australas. J. Anim. Sci. 2020, 33, 778–787. [Google Scholar] [CrossRef]
- Olfati, A.; Mojtahedin, A.; Sadeghi, T.; Akbari, M.; Martínez-Pastor, F. Comparison of growth performance and immune responses of broiler chicks reared under heat stress, cold stress and thermoneutral conditions. Span. J. Agric. Res. 2018, 16, e0505. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Ou, J.; Luo, Z.; Kim, I.H. Effect of dietary β-1,3-glucan supplementation and heat stress on growth performance, nutrient digestibility, meat quality, organ weight, ileum microbiota, and immunity in broilers. Poult. Sci. 2020, 99, 4969–4977. [Google Scholar] [CrossRef] [PubMed]
- Jahanian, R.; Rasouli, E. Dietary chromium methionine supplementation could alleviate immunosuppressive effects of heat stress in broiler chicks1. J. Anim. Sci. 2015, 93, 3355–3363. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Yu, Q.; He, Y.; Hu, R.; Xia, S.; He, J. Dietary resveratrol supplementation inhibits heat stress-induced high-activated innate immunity and inflammatory response in spleen of yellow-feather broilers. Poult. Sci. 2019, 98, 6378–6387. [Google Scholar] [CrossRef]
- Hirakawa, R.; Nurjanah, S.; Furukawa, K.; Murai, A.; Kikusato, M.; Nochi, T.; Toyomizu, M. Heat stress causes immune abnormalities via massive damage to effect proliferation and differentiation of lymphocytes in broiler chickens. Front. Veter. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Padgett, D.A.; Glaser, R. How stress influences the immune response. Trends Immunol. 2003, 24, 444–448. [Google Scholar] [CrossRef]
- Song, J.-H.; Kim, K.-J.; Choi, S.-Y.; Koh, E.-J.; Park, J.; Lee, B.-Y. Korean ginseng extract ameliorates abnormal immune response through the regulation of inflammatory constituents in Sprague Dawley rat subjected to environmental heat stress. J. Ginseng Res. 2019, 43, 252–260. [Google Scholar] [CrossRef]
- Abdel-Latif, M.; Sakran, T.; Badawi, Y.K.; Abdel-Hady, D.S. Influence of Moringa oleifera extract, vitamin C, and sodium bicarbonate on heat stress-induced HSP70 expression and cellular immune response in rabbits. Cell Stress Chaperon. 2018, 23, 975–984. [Google Scholar] [CrossRef]
- Kamel, N.N.; Ahmed, A.M.H.; Mehaisen, G.M.K.; Mashaly, M.M.; Abass, A.O. Depression of leukocyte protein synthesis, immune function and growth performance induced by high environmental temperature in broiler chickens. Int. J. Biometeorol. 2017, 61, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Mashaly, M.M.; Hendricks, G.L.; Kalama, M.A.; Gehad, A.E.; Abbas, A.O.; Patterson, P.H. Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult. Sci. 2004, 83, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, R.; Amin, G.; Ardekani, M.R.S. A Review on Citrullus colocynthis Schrad.: From Traditional Iranian Medicine to Modern Phytotherapy. J. Altern. Complement. Med. 2012, 18, 551–554. [Google Scholar] [CrossRef]
- Hussain, A.I.; Rathore, H.; Sattar, M.Z.; Chatha, S.A.S.; Sarker, S.D.; Gilani, A.H. Citrullus colocynthis (L.) Schrad (bitter apple fruit): A review of its phytochemistry, pharmacology, traditional uses and nutritional potential. J. Ethnopharmacol. 2014, 155, 54–66. [Google Scholar] [CrossRef]
- Halla, N.; Boucherit, K.; Boucherit-Otmani, Z.; Touati, F.Z.; Rahmani, N.; Aid, I. Ammodaucus leucotrichus and Citrullus colocynthis from algerian Sahara: Ethnopharmacological application, phytochemical screening, polyphenols content and antioxidant activity of hydromethanolic extracts. J. King Saud Univ. Sci. 2019, 31, 541–548. [Google Scholar] [CrossRef]
- Ostovar, M.; Akbari, A.; Anbardar, M.H.; Iraji, A.; Salmanpour, M.; Ghoran, S.H.; Heydari, M.; Shams, M. Effects of Citrullus colocynthis L. in a rat model of diabetic neuropathy. J. Integr. Med. 2020, 18, 59–67. [Google Scholar] [CrossRef]
- Al-Hwaiti, M.S.; Alsbou, E.M.; Abu Sheikha, G.; Bakchiche, B.; Pham, T.H.; Thomas, R.H.; Bardaweel, S.K. Evaluation of the anticancer activity and fatty acids composition of “Handal” ( Citrullus colocynthis L.) seed oil, a desert plant from south Jordan. Food Sci. Nutr. 2021, 9, 282–289. [Google Scholar] [CrossRef]
- Jemai, R.; Drira, R.; Makni, M.; Fetoui, H.; Sakamoto, K. Colocynth (Citrullus colocynthis) seed extracts attenuate adipogenesis by down-regulating PPARγ/ SREBP-1c and C/EBPα in 3T3-L1 cells. Food Biosci. 2020, 33. [Google Scholar] [CrossRef]
- Marzouk, B.; Mahjoub, M.A.; Bouraoui, A.; Fenina, N.; Aouni, M.; Marzouk, Z. Anti-inflammatory and analgesic activities of a new cucurbitacin isolated from Citrullus colocynthis seeds. Med. Chem. Res. 2012, 22, 3984–3990. [Google Scholar] [CrossRef]
- Marzouk, B.; Marzouk, Z.; Fenina, N.; Bouraoui, A.; Aouni, M. Anti-inflammatory and analgesic activities of Tunisian Citrullus colocynthis Schrad. immature fruit and seed organic extracts. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 665–672. [Google Scholar]
- Marzouk, B.; Marzouk, Z.; Haloui, E.; Turki, M.; Bouraoui, A.; Aouni, M.; Fenina, N. Anti-inflammatory evaluation of immature fruit and seed aqueous extracts from several populations of Tunisian Citrullus colocynthis Schrad. Afr. J. Biotechnol. 2011, 10, 4217–4225. [Google Scholar]
- Marzouk, B.; Marzouk, Z.; Mastouri, M.; Fenina, N.; Aouni, M. Comparative evaluation of the antimicrobial activity of Citrullus colocynthis immature fruit and seed organic extracts. Afr. J. Biotechnol 2011, 10, 2130–2134. [Google Scholar]
- Sawaya, W.N.; Daghir, N.J.; Khalil, J.K. Citrullus colocynthis seeds as a potential source of protein for food and feed. J. Agric. Food Chem. 1986, 34, 285–288. [Google Scholar] [CrossRef]
- Sawaya, W.N.; Daghir, N.J.; Khan, P. Chemical characterization and edibility of the oil extracted from Citrullus colocynthis seeds. J. Food Sci. 1983, 48, 104–106. [Google Scholar] [CrossRef]
- Shafaei, H.; Solaeymanirad, J.; Mahdavi, R.; Ostad Rahimi, A.R.; Rezazadeh, H.; Argani, H.; Rashidi, M.R.; Nazemieh, H.; Delazar, A. The potentiating effects of Citrullus colocynthis extract on immune system. Med. J. Tabriz Univ. Med Sci. 2007, 29, 77–82. [Google Scholar]
- Abass, O.A.; Kamel, N.N.; Khalifa, W.H.; Gouda, G.F.; El-Manylawi, M.A.F.; Mehaisen, G.; Mashaly, M.M. Propolis supplementation attenuates the negative effects of oxidative stress induced by paraquat injection on productive performance and immune function in turkey poults. Poult. Sci. 2017, 96, 4419–4429. [Google Scholar] [CrossRef] [PubMed]
- Mehaisen, G.M.K.; Eshak, M.G.; Elkaiaty, A.M.; Atta, A.-R.M.M.; Mashaly, M.M.; Abass, A.O. Comprehensive growth performance, immune function, plasma biochemistry, gene expressions and cell death morphology responses to a daily corticosterone injection course in broiler chickens. PLoS ONE 2017, 12, e0172684. [Google Scholar] [CrossRef]
- Gehad, A.E.; Mehaisen, G.M.; Abbas, A.O.; Mashaly, M.M. The role of light program and melatonin on alleviation of inflammation induced by lipopolysaccharide injection in broiler chickens. Int. J. Poult. Sci. 2008, 7, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Loa, C.C.; Lin, T.L.; Wu, C.C.; Bryan, T.; Thacker, H.L.; Hooper, T.; Schrader, D. Humoral and cellular immune responses in turkey poults infected with turkey Coronavirus. Poult. Sci. 2001, 80, 1416–1424. [Google Scholar] [CrossRef]
- Abbas, A.O.; Alaqil, A.A.; El-Beltagi, H.S.; El-Atty, H.K.A.; Kamel, N.N. Modulating laying hens productivity and immune performance in response to oxidative stress induced by E. coli challenge using dietary propolis supplementation. Antioxidants 2020, 9, 893. [Google Scholar] [CrossRef]
- Marketon, J.; Glaser, R. Stress hormones and immune function. Cell. Immunol. 2008, 252, 16–26. [Google Scholar] [CrossRef]
- Cruz-Topete, D.; Cidlowski, J. One hormone, two actions: Anti- and pro-Inflammatory effects of glucocorticoids. Neuroimmunomodulation 2015, 22, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Sohail, M.U.; Ijaz, A.; Yousaf, M.S.; Ashraf, K.; Zaneb, H.; Aleem, M.; Rehman, H. Alleviation of cyclic heat stress in broilers by dietary supplementation of mannan-oligosaccharide and Lactobacillus-based probiotic: Dynamics of cortisol, thyroid hormones, cholesterol, C-reactive protein, and humoral immunity. Poult. Sci. 2010, 89, 1934–1938. [Google Scholar] [CrossRef]
- Habibian, M.; Ghazi, S.; Moeini, M.M.; Abdolmohammadi, A. Effects of dietary selenium and vitamin E on immune response and biological blood parameters of broilers reared under thermoneutral or heat stress conditions. Int. J. Biometeorol. 2014, 58, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Akhavan-Salamat, H.; Ghasemi, H.A. Alleviation of chronic heat stress in broilers by dietary supplementation of betaine and turmeric rhizome powder: Dynamics of performance, leukocyte profile, humoral immunity, and antioxidant status. Trop. Anim. Heal. Prod. 2016, 48, 181–188. [Google Scholar] [CrossRef]
- Lee, G.Y.; Han, S.N. The role of vitamin E in immunity. Nutrients 2018, 10, 1614. [Google Scholar] [CrossRef] [Green Version]
- Lewis, E.D.; Meydani, S.N.; Wu, D. Regulatory role of vitamin E in the immune system and inflammation. IUBMB Life 2019, 71, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Nehdi, I.A.; Sbihi, H.; Tan, C.P.; Al-Resayes, S.I. Evaluation and characterisation of Citrullus colocynthis (L.) Schrad seed oil: Comparison with Helianthus annuus (sunflower) seed oil. Food Chem. 2013, 136, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Reiter, E.; Jiang, Q.; Christen, S. Anti-inflammatory properties of α- and γ-tocopherol. Mol. Asp. Med. 2007, 28, 668–691. [Google Scholar] [CrossRef] [Green Version]
- Wasti, S.; Sah, N.; Mishra, B. Impact of heat stress on poultry health and performances, and potential mitigation strategies. Animals 2020, 10, 1266. [Google Scholar] [CrossRef]
- Hosseini-Vashan, S.J.; Golian, A.; Yaghobfar, A. Growth, immune, antioxidant, and bone responses of heat stress-exposed broilers fed diets supplemented with tomato pomace. Int. J. Biometeorol. 2015, 60, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sa, Y.; Wang, G.; Pan, X.; Zhen, Y.; Cheng, X.; Zhang, K.; Fu, L.; Wang, H.; Liu, B. The protective effects of Citrullus colocynthis on inhibiting oxidative damage and autophagy-associated cell death in Parkinson’s disease. J. Taiwan Inst. Chem. Eng. 2019, 100, 18–25. [Google Scholar] [CrossRef]
- Hussain, A.I.; Rathore, H.; Sattar, M.Z.; Chatha, S.A.S.; Ahmad, F.U.D.; Ahmad, A.; Johns, E. Phenolic profile and antioxidant activity of various extracts from Citrullus colocynthis (L.) from the Pakistani flora. Ind. Crop. Prod. 2013, 45, 416–422. [Google Scholar] [CrossRef]
- Bourhia, M.; Messaoudi, M.; Bakrim, H.; Mothana, R.A.; Sddiqui, N.A.; Almarfadi, O.M.; El Mzibri, M.; Gmouh, S.; Laglaoui, A.; Benbacer, L. Citrullus colocynthis (L.) Schrad: Chemical characterization, scavenging and cytotoxic activities. Open Chem. 2020, 18, 986–994. [Google Scholar] [CrossRef]
- Siddiqui, S.H.; Kang, D.; Park, J.; Khan, M.; Shim, K. Chronic heat stress regulates the relation between heat shock protein and immunity in broiler small intestine. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Kregel, K.C. Invited Review: Heat shock proteins: Modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physiol. 2002, 92, 2177–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitley, D.; Goldberg, S.P.; Jordan, W.D. Heat shock proteins: A review of the molecular chaperones. J. Vasc. Surg. 1999, 29, 748–751. [Google Scholar] [CrossRef] [Green Version]
- Gruys, E.; Toussaint, M.J.M.; Niewold, T.A.; Koopmans, S.J. Acute phase reaction and acute phase proteins. J. Zhejiang Univ. Sci. B 2005, 6, 1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, A.; Hussain, S. Hepato toxic or hepato protective: A review of hepatic effects of Citrullus colocynthis. J. Pharmacogn. Phytochem. 2018, 7, 1226–1233. [Google Scholar]
- Vakiloddin, S.; Fuloria, N.; Fuloria, S.; Dhanaraj, S.A.; Balaji, K.; Karupiah, S. Evidences of hepatoprotective and antioxidant effect of Citrullus colocynthis fruits in paracetamol induced hepatotoxicity. Pak. J. Pharm. Sci. 2015, 28, 951–957. [Google Scholar] [PubMed]


{kind=link}
| Ingredients | g/kg as Fed |
|---|---|
| Corn | 626 |
| Gluten meal | 20.0 |
| Soybean meal, 48% CP | 292 |
| Soya oil | 25.0 |
| Di-calcium phosphate | 16.5 |
| Limestone | 7.00 |
| Salt | 4.50 |
| Vitamin-Mineral Premix * | 5.00 |
| L-threonine | 0.50 |
| DL-Methionine | 0.80 |
| L-Lysine | 1.70 |
| Choline chloride | 0.20 |
| 3-Phytase | 0.80 |
| Nutrient content | |
| Chemical analysis | |
| Metabolizable energy (kcal/kg) | 3150 |
| Crude protein, g/kg | 202 |
| Crude fat, g/kg | 58.8 |
| Crude fiber, g/kg | 25.1 |
| Ash, g/kg | 5.63 |
| Calculated analysis | |
| Calcium, g/kg | 8.48 |
| Available Phosphorus, g/kg | 4.21 |
| DL-methionine, g/kg | 5.68 |
| L-Lysine, g/kg | 11.00 |
| Sodium | 1.40 |
| Parameter | Thermo-Neutral | Heat Stress | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Without CC | With CC | Without CC | With CC | HS | CC | HS × CC | ||
| BW, 22 d, g | 754 | 738 | 743 | 757 | 29.35 | NS | NS | NS |
| BW, 42 d, g | 2432 a | 2488 a | 1740 c | 1983 b | 71.30 | <0.0001 | NS | NS |
| BWG, g | 1678 a | 1750 a | 997 c | 1226 b | 75.16 | <0.0001 | NS | NS |
| FI, g | 3134 a | 3189 a | 2375 b | 2443 b | 131.37 | <0.0001 | NS | NS |
| FCR | 1.88 c | 1.82 c | 2.38 a | 2.00 b | 0.037 | <0.0001 | <0.0001 | 0.0005 |
| Parameter | Thermo-Neutral | Heat Stress | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Without CC | With CC | Without CC | With CC | HS | CC | HS × CC | ||
| TNF-α, pg/mL | 94.27 c | 83.95 c | 186.13 a | 151.63 b | 4.92 | <0.0001 | 0.0002 | 0.0232 |
| Corticosterone, ng/mL | 2.36 c | 1.93 c | 9.62 a | 5.80 b | 0.56 | <0.0001 | 0.0011 | 0.0067 |
| MDA, µM/mL | 1.08 c | 1.00 c | 4.07 a | 2.05 b | 0.23 | <0.0001 | 0.0002 | 0.0004 |
| H/L ratio | 0.38 c | 0.35 c | 0.90 a | 0.67 b | 0.06 | <0.0001 | 0.0361 | NS |
| SOD, U/mL | 5.47 a | 5.71 a | 3.36 c | 4.57 b | 0.28 | <0.0001 | 0.0164 | NS |
| TAC, U/mL | 8.68 b | 9.54 a | 5.11 d | 7.10 c | 0.29 | <0.0001 | <0.0001 | NS |
| Parameter | Thermo-Neutral | Heat Stress | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Without CC | With CC | Without CC | With CC | HS | CC | HS × CC | ||
| TWBC, 103/mL | 42.33 b | 56.83 a | 24.33 c | 46.50 b | 2.97 | 0.0001 | <0.0001 | NS |
| Anti-SRBCsAB, log2 | 6.50 ab | 7.33 a | 3.83 c | 5.33 b | 0.42 | <0.0001 | 0.0113 | NS |
| Tow web swelling, mm | 0.34 a | 0.36 a | 0.18 c | 0.26 b | 0.02 | <0.0001 | 0.005 | NS |
| B-lymphocytes SI | 2.40 b | 2.97 a | 0.69 c | 2.14 b | 0.15 | <0.0001 | <0.0001 | 0.0074 |
| T-lymphocytes SI | 4.93 b | 6.83 a | 2.12 c | 4.12 b | 0.30 | <0.0001 | <0.0001 | NS |
| Parameter | Thermo-Neutral | Heat Stress | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Without CC | With CC | Without CC | With CC | HS | CC | HS × CC | ||
| TP, g/dL | 3.35 c | 3.55 c | 5.74 a | 4.13 b | 0.19 | <0.0001 | 0.0014 | 0.0001 |
| Albumin, g/dL | 1.78 ab | 1.86 a | 1.20 c | 1.61 b | 0.06 | <0.0001 | 0.0007 | 0.0138 |
| Globulin, g/dL | 1.56 c | 1.68 c | 4.54 a | 2.52 b | 0.19 | <0.0001 | <0.0001 | <0.0001 |
| A/G ratio | 1.16 a | 1.22 a | 0.27 c | 0.67 b | 0.11 | <0.0001 | 0.0500 | NS |
| AST, U/mL | 84.96 c | 81.08 c | 139.37 a | 113.98 b | 3.28 | <0.0001 | 0.0002 | 0.0037 |
| ALT, U/mL | 10.98 c | 10.45 c | 23.89 a | 18.87 b | 1.09 | <0.0001 | 0.0186 | NS |
| Creatinine, mg/dL | 0.30 b | 0.31 b | 0.51 a | 0.37 b | 0.02 | <0.0001 | 0.0161 | 0.0035 |
| Urea, mg/dL | 4.95 b | 4.41 b | 6.70 a | 5.39 b | 0.32 | 0.0004 | 0.0093 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alzarah, M.I.; Althobiati, F.; Abbas, A.O.; Mehaisen, G.M.K.; Kamel, N.N. Citrullus colocynthis Seeds: A Potential Natural Immune Modulator Source for Broiler Reared under Chronic Heat Stress. Animals 2021, 11, 1951. https://doi.org/10.3390/ani11071951
Alzarah MI, Althobiati F, Abbas AO, Mehaisen GMK, Kamel NN. Citrullus colocynthis Seeds: A Potential Natural Immune Modulator Source for Broiler Reared under Chronic Heat Stress. Animals. 2021; 11(7):1951. https://doi.org/10.3390/ani11071951
Chicago/Turabian StyleAlzarah, Mohamed I., Fayez Althobiati, Ahmed O. Abbas, Gamal M. K. Mehaisen, and Nancy N. Kamel. 2021. "Citrullus colocynthis Seeds: A Potential Natural Immune Modulator Source for Broiler Reared under Chronic Heat Stress" Animals 11, no. 7: 1951. https://doi.org/10.3390/ani11071951