
Alterations in the Fecal Microbiome and Metabolome of Horses with Antimicrobial-Associated Diarrhea Compared to Antibiotic-Treated and Non-Treated Healthy Case Controls

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Microbiome
2.2.1. Sample Preparation
2.2.2. Statistical Analysis of Microbiome Data
2.3. Metabolome
2.3.1. Sample Preparation
2.3.2. Metabolomic Analysis
2.3.3. Statistical Analysis of Metabolomic Data
3. Results
3.1. Subjects
3.2. Microbiome
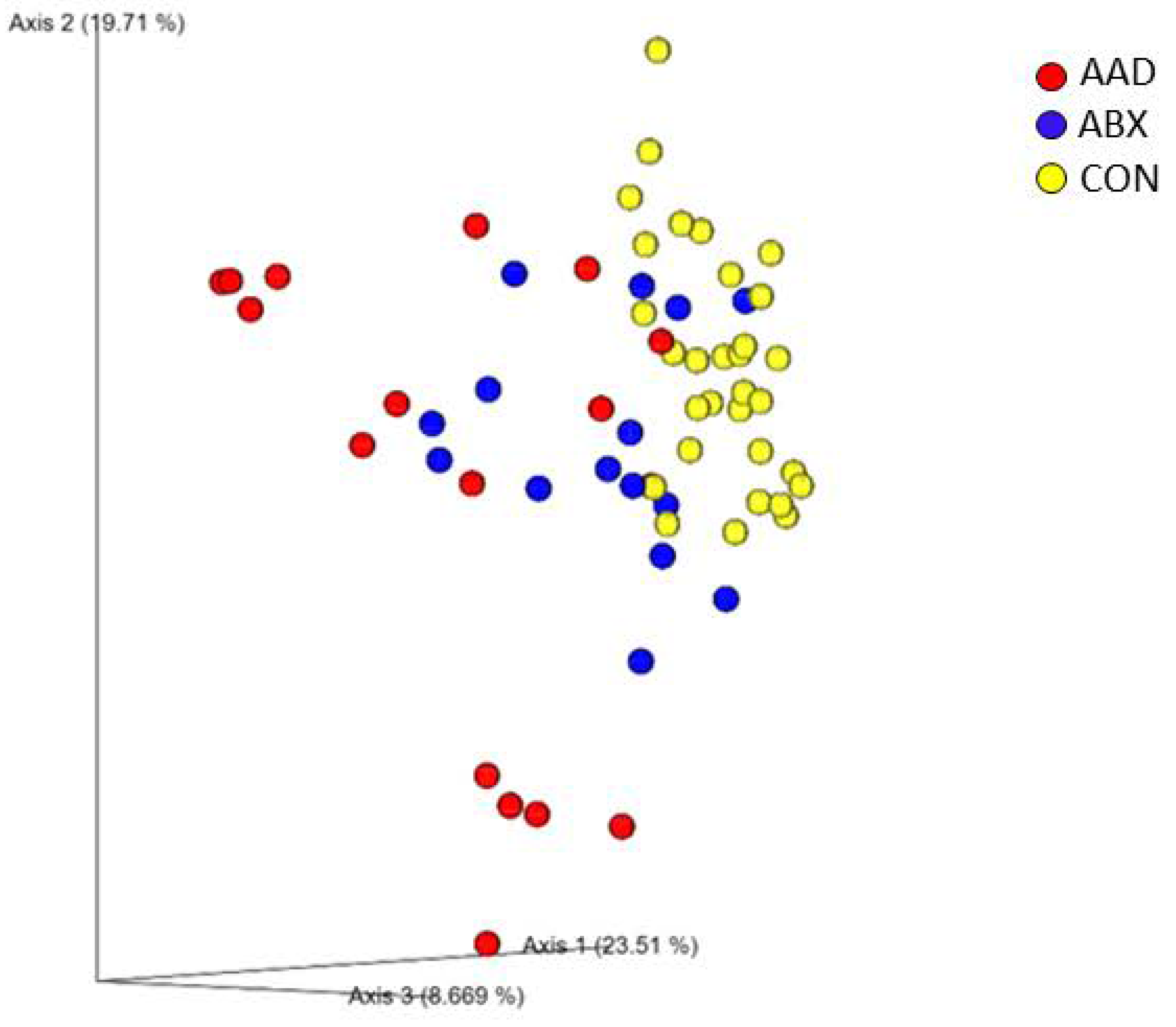
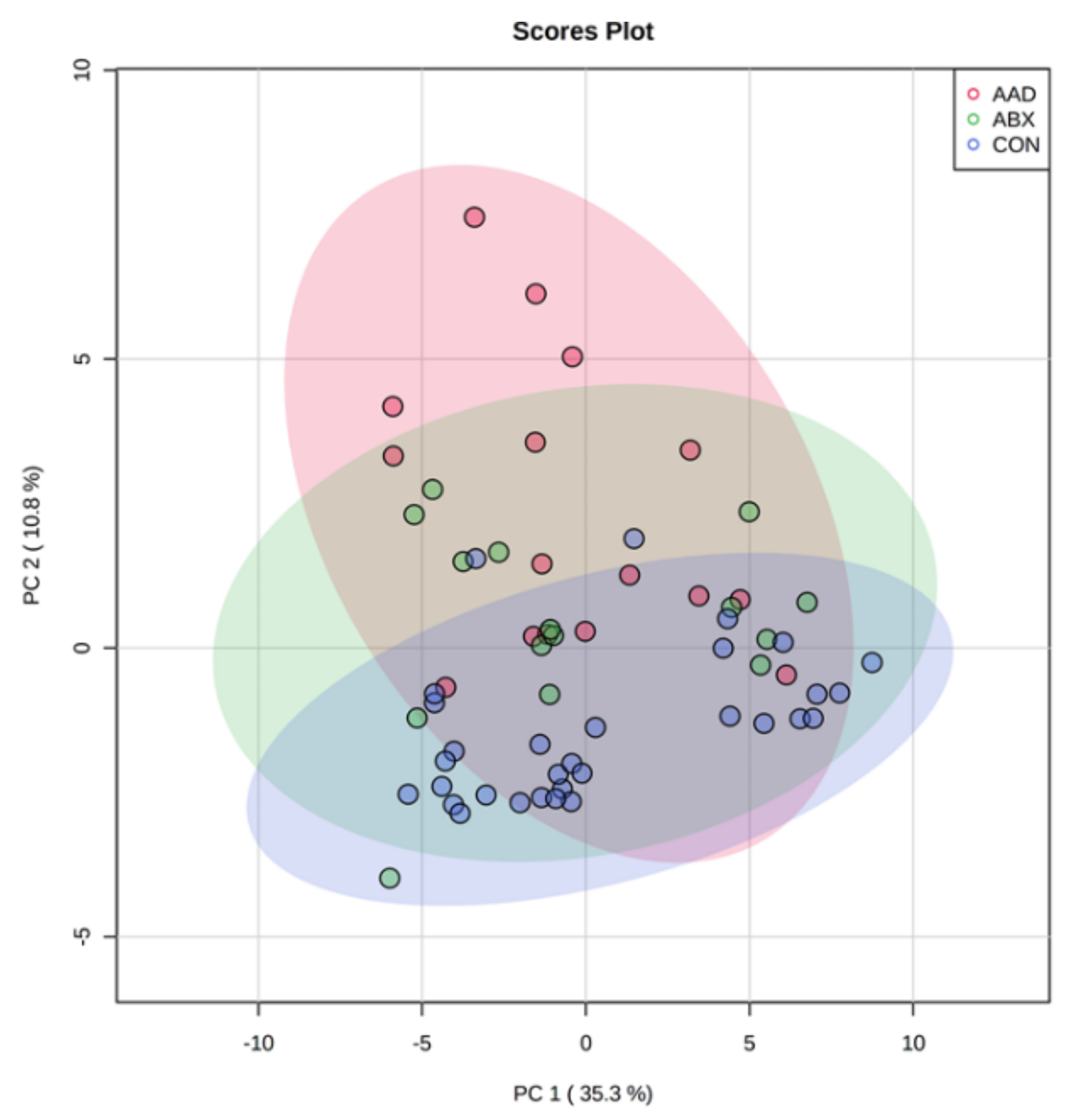
3.2.1. Microbial Community Composition (Beta Diversity)
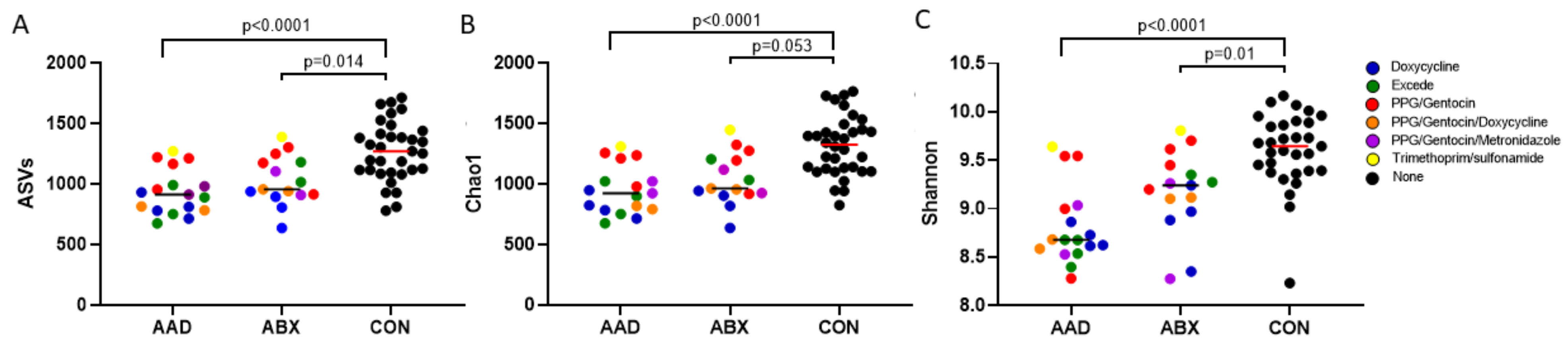
3.2.2. Species Richness and Evenness (Alpha Diversity)
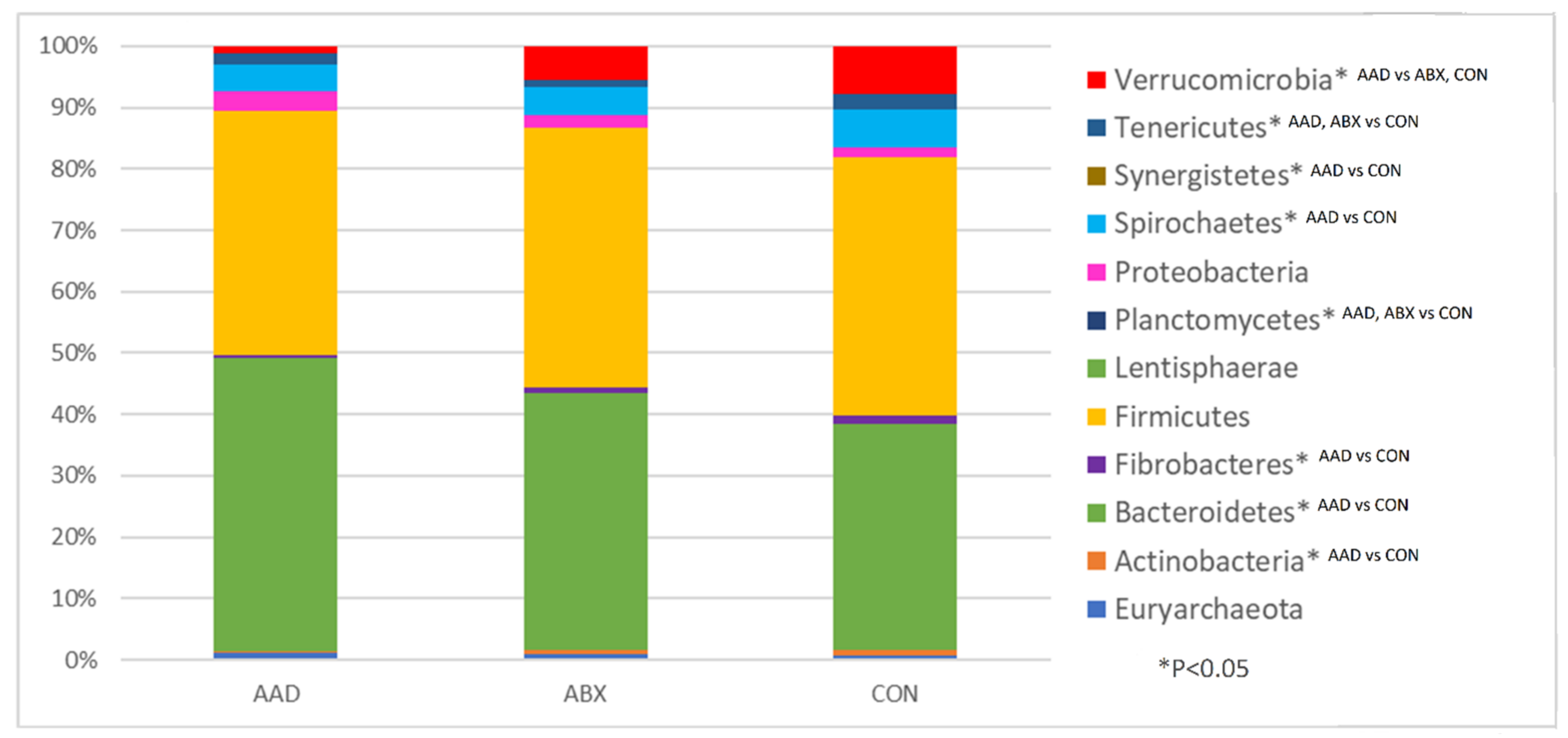
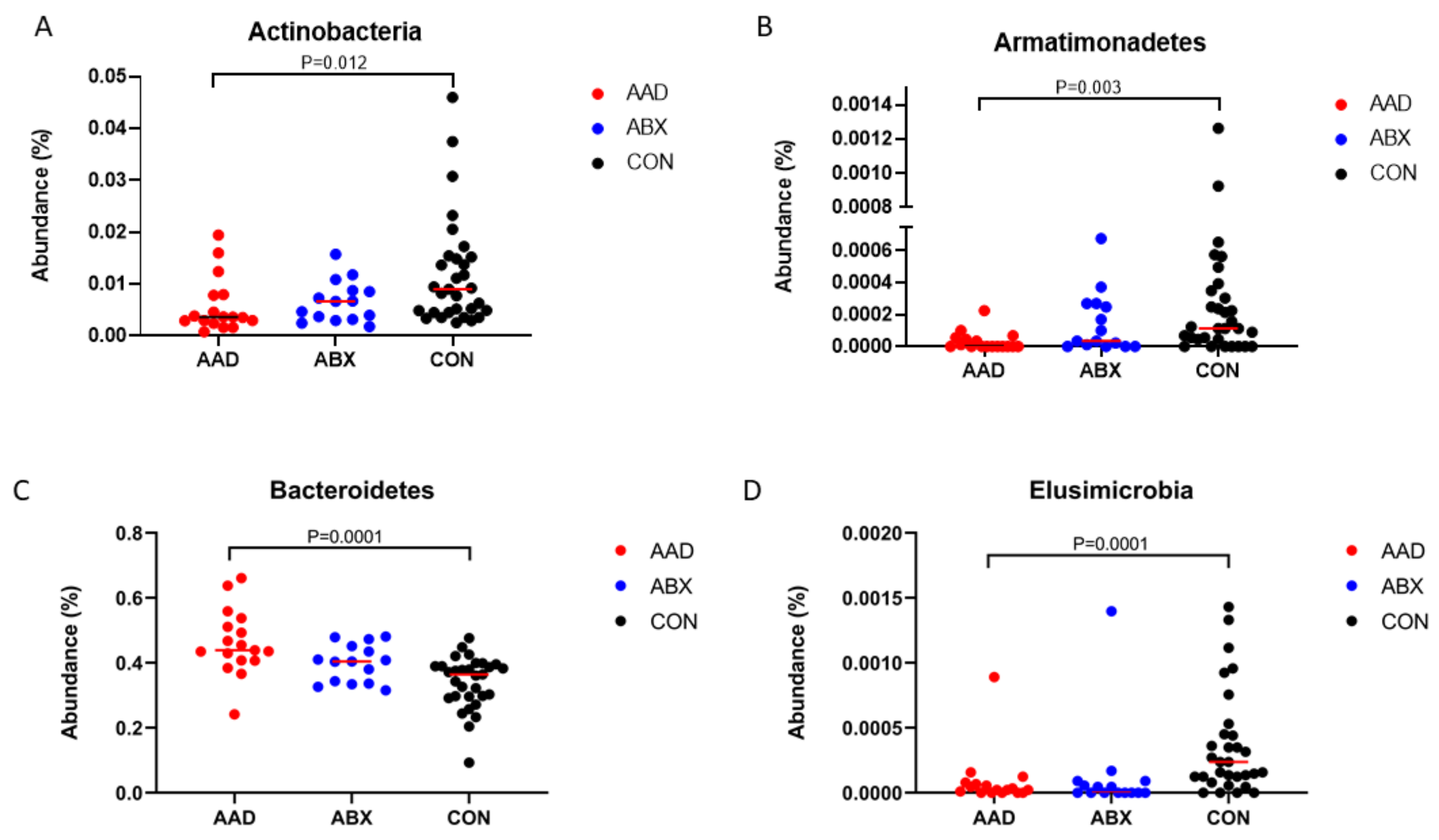
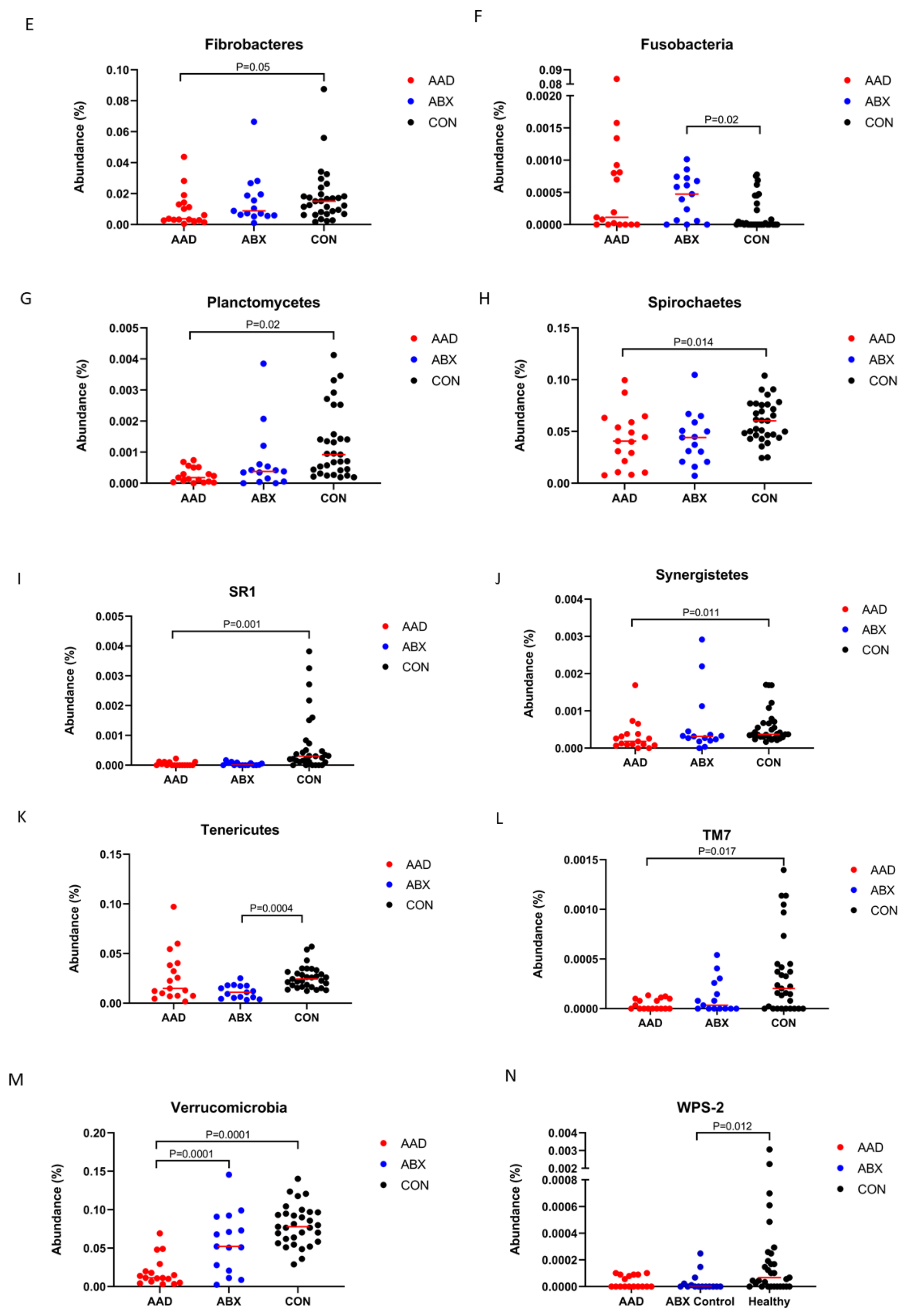
3.2.3. Taxonomy
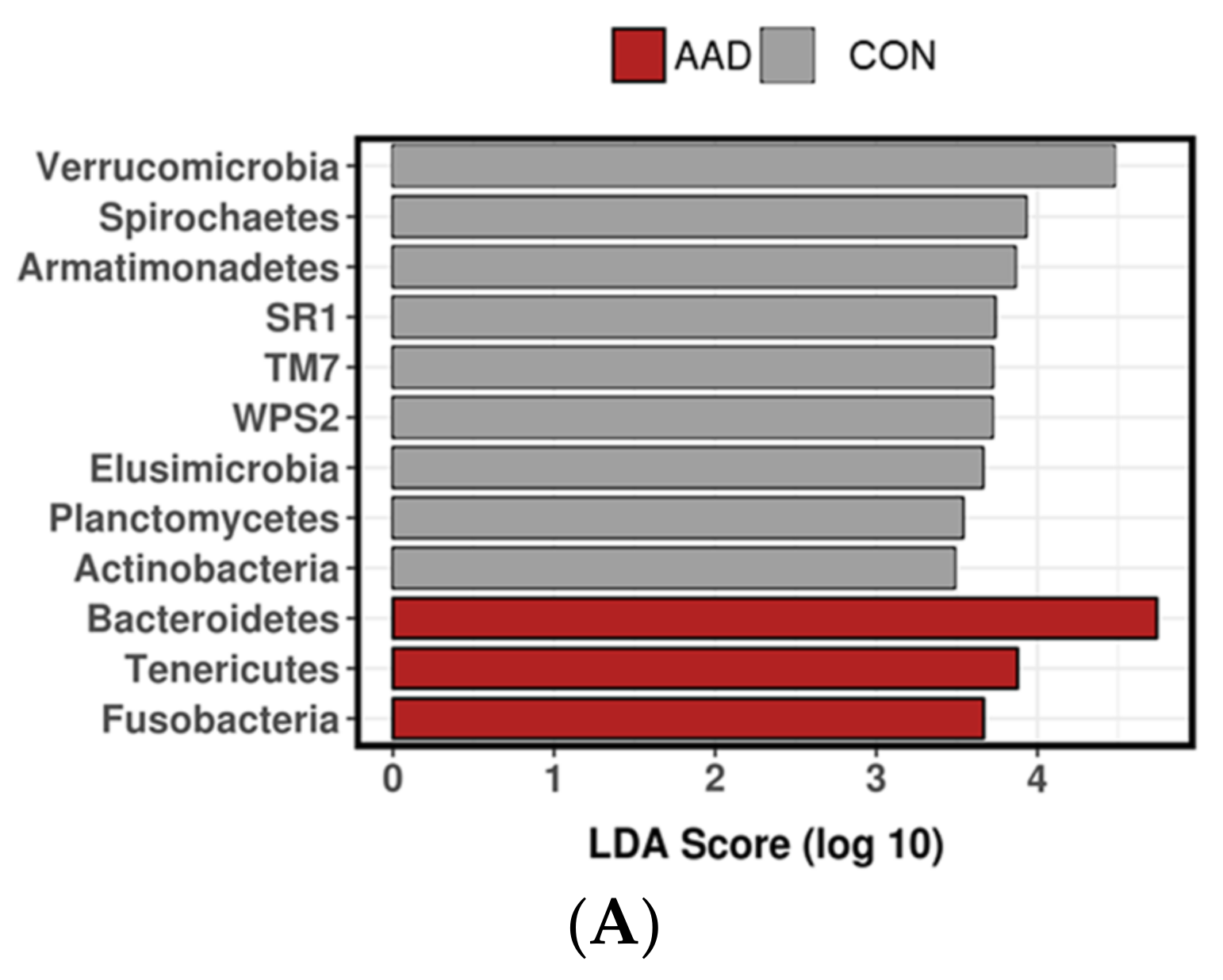
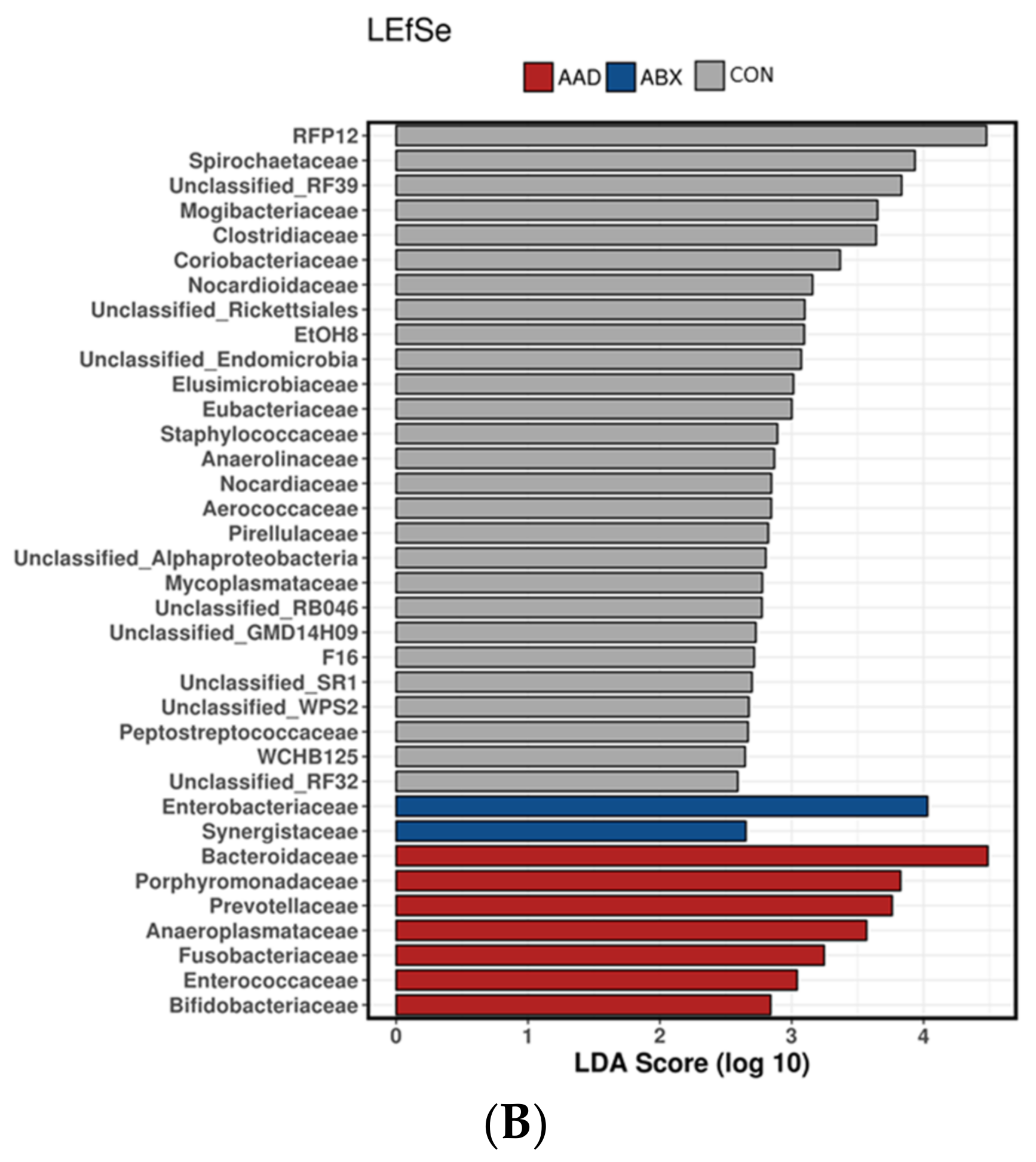
3.2.4. Linear Discriminant Analysis Effect Size (LEfSe)
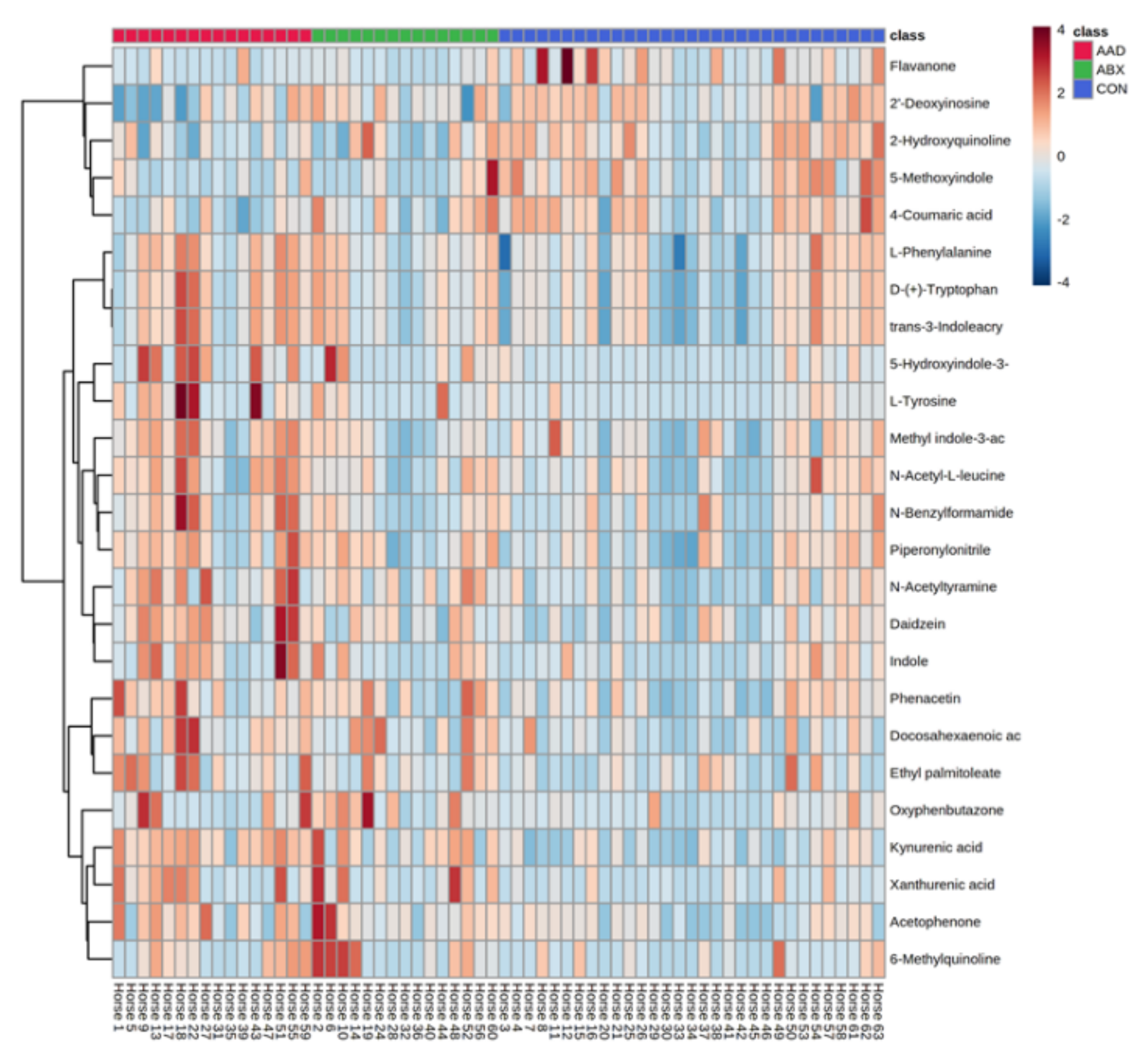
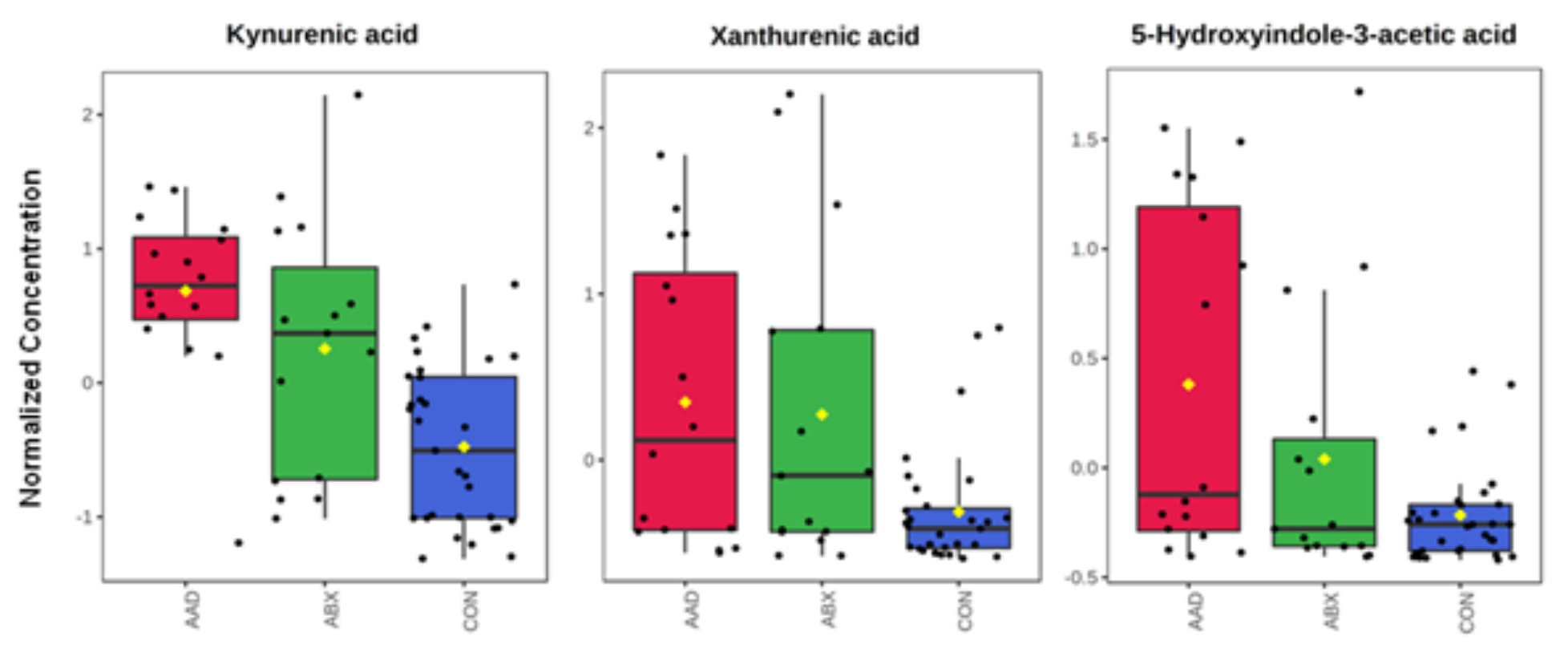
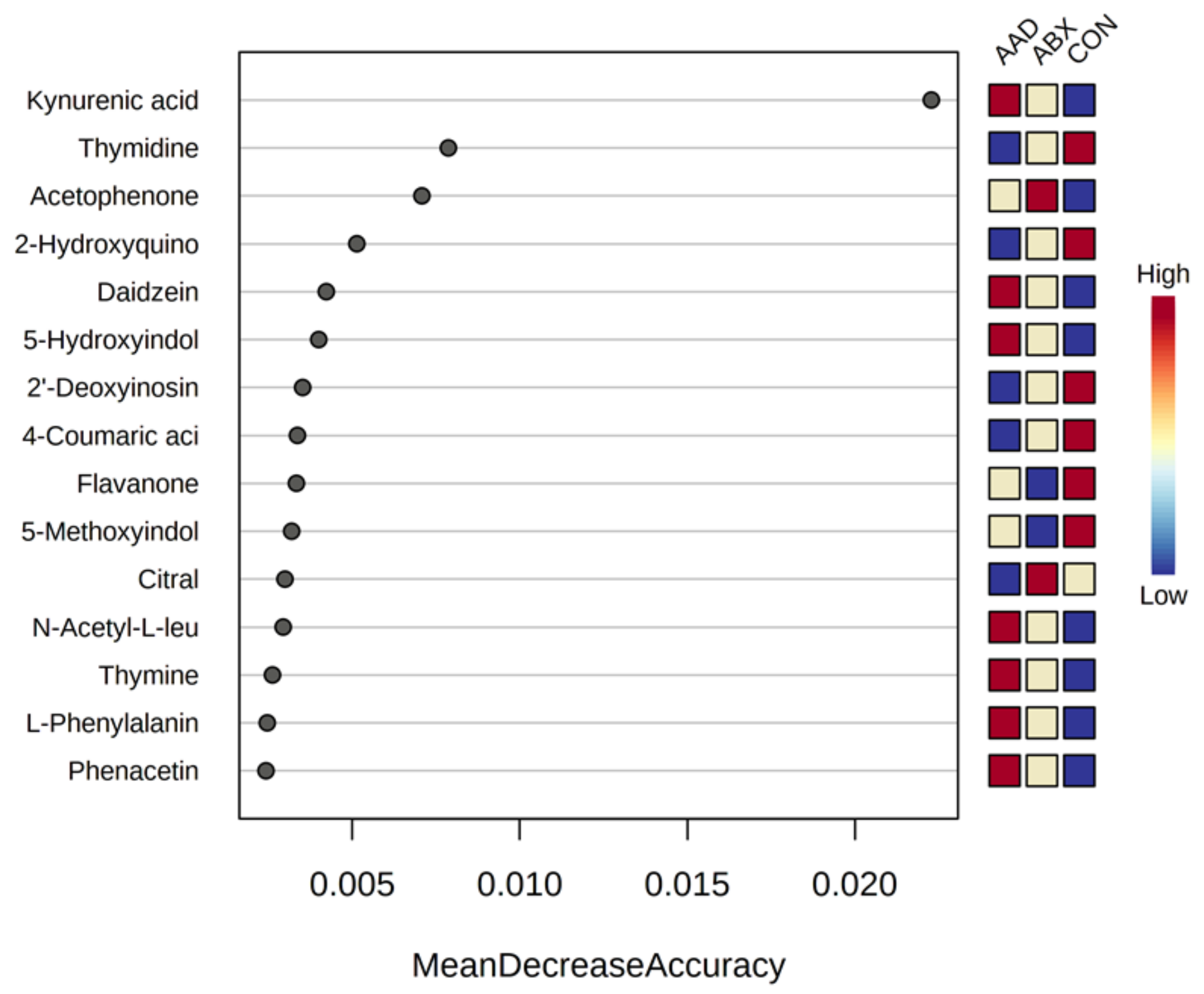
3.3. Metabolomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- McGorum, B.C.; Pirie, R.S. Antimicrobial associated diarrhoea in the horse. Part 1: Overview, pathogenesis and risk factors. Equine Vet. Educ. 2009, 21, 610–616. [Google Scholar] [CrossRef]
- Weese, J.; Baptiste, K.E.; Baverud, V.; Toutain, P.L. Guidelines of antimicrobial use in horses. In Guide to Antimicrobial use in Animals, 1st ed.; Guardabassi, L., Jensen, L.B., Kruse, H., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2006; pp. 161–182. [Google Scholar]
- Papich, M.G. Antimicrobial therapy for gastrointestinal diseases. Vet. Clin. Equine Pract. 2003, 19, 645–663. [Google Scholar] [CrossRef]
- Andersson, G.; Ekman, L.; Månsson, I.; Persson, S.; Rubarth, S.; Tufvesson, G. Lethal complications following administration of oxytetracycline in the horse. Nord. Vet. Med. 1971, 23, 9–22. [Google Scholar] [PubMed]
- Barr, B.S.; Waldridge, B.M.; Morresey, P.R.; Reed, S.M.; Clark, C.; Belgrave, R.; Donecker, J.M.; Weigel, D.J. Antimicrobial-associated diarrhoea in three equine referral practices. Equine Vet. J. 2013, 45, 154–158. [Google Scholar] [CrossRef]
- White, G.; Prior, S.D. Comparative effects of oral administration of trimethoprim/sulphadiazine or oxytetracycline on the faecal flora of horses. Vet. Rec. 1982, 111, 316–318. [Google Scholar] [CrossRef]
- Gustaffson, A.; Baverud, V.; Gunnarsson, A.; Rantzien, M.H.; Lindholm, A.; Franklin, A. The association of erythromycin ethylsuccinate with acute colitis in horses in Sweden. Equine Vet. J. 1997, 29, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Raisbeck, M.F.; Holt, G.R.; Osweiler, G.D. Lincomycin-associated colitis in horses. J. Am. Vet. Med. Assoc. 1981, 179, 362–363. [Google Scholar]
- Staempfli, H.; Prescott, J.; Carman, R.; McCutcheon, L. Use of Bacitracin in the prevention and treatment of experimentally-induced idiopathic colitis in horses. Can. J. Vet. Res. 1992, 56, 233–236. [Google Scholar]
- Weese, J. Anti-Microbial Associated Diarrhea in 40 Horses: 1997–1999. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2000. [Google Scholar]
- Hollis, A.R.; Wilkins, P.A. Current controversies in equine antimicrobial therapy. Equine Vet. Educ. 2009, 21, 216–224. [Google Scholar] [CrossRef]
- Wilson, D.A.; MacFadden, K.E.; Green, E.M.; Crabill, M.; Frankeny, R.L.; Thorne, J.G. Case Control and Historical Cohort Study of Diarrhea Associated with Administration of Trimethoprim-Potentiated Sulphonamides to Horses and Ponies. J. Vet. Intern. Med. 1996, 10, 258–264. [Google Scholar] [CrossRef]
- Baverud, V.; Gustaffsson, A.; Franklin, A.; Lindholm, A.; Gunnarsson, A. Clostridium difficile associated with acute colitis in mature horses treated with antibiotics. Equine Vet. J. 1997, 29, 279–284. [Google Scholar] [CrossRef]
- MCKellare, Q.; Varma, K.J. Pharmacokinetics and tolerance of florfenicol in Equidae. Equine Vet. J. 1996, 28, 209–213. [Google Scholar] [CrossRef]
- McGorum, B.; Dixon, P.; Smith, D. Use of metronidazole in equine idiopathic toxaemic colitis. Vet. Rec. 1998, 142, 635–638. [Google Scholar] [CrossRef]
- Arnold, C.E.; Isaiah, A.; Pilla, R.; Lidbury, J.; Coverdale, J.S.; Callaway, T.R.; Lawhon, S.D.; Steiner, J.; Suchodolski, J.S. The cecal and fecal microbiomes and metabolomes of horses before and after metronidazole administration. PLoS ONE 2020, 15, e0232905. [Google Scholar] [CrossRef]
- Cohen, N.D.; Woods, A.M. Characteristics and risk factors for failure of horses with acute diarrhea to survive: 122 cases (1990–1996). J. Am. Vet. Med. Assoc. 1999, 214, 382–390. [Google Scholar]
- Costa, M.C.; Arroyo, L.G.; Allen-Vercoe, E.; Stampfli, H.R.; Kim, P.T.; Sturgeon, A.; Weese, J.S. Comparison of the fecal microbiota of healthy horses and horses with colitis by high throughput sequencing of the V3-V5 region of the 16S rRNA gene. PLoS ONE 2012, 7, e41484. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, C.; Taminiau, B.; Brévers, B.; Avesani, V.; Van Broeck, J.; Leroux, A.; Gallot, M.; Bruwier, A.; Amory, H.; Delmée, M.; et al. Faecal microbiota characterisation of horses using 16 rdna barcoded pyrosequencing, and carriage rate of clostridium difficile at hospital admission. BMC Microbiol. 2015, 15, e181. [Google Scholar] [CrossRef] [Green Version]
- McKinney, C.A.; Oliveira, B.C.M.; Bedenice, D.; Paradis, M.R.; Mazan, M.; Sage, S.; Sanchez, A.; Widmer, G. The fecal microbiota of healthy donor horses and geriatric recipients undergoing fecal microbial transplantation for the treatment of diarrhea. PLoS ONE 2020, 15, e0230148. [Google Scholar] [CrossRef]
- Costa, M.C.; Stampfli, H.R.; Arroyo, L.G.; Allen-Vercoe, E.; Gomes, R.G.; Weese, J.S. Changes in the equine fecal microbiota associated with the use of systemic antimicrobial drugs. BMC Vet. Res. 2015, 11, e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liepman, R.S. Alterations in the Fecal Microbiome of Healthy Horses in Response to Antibiotic Treatment. Master’s Thesis, The Ohio State University, Columbus, OH, USA, 2015. [Google Scholar]
- Arnold, C.; Pilla, R.; Chaffin, M.; Steiner, J.; Suchodolski, J. The fecal microbiome of horses with antimicrobial associated diarrhea or Salmonella compared to healthy horses. In Proceedings of the 65th Meeting of the American Association of Equine Practitioners, Denver, CO, USA, 11 December 2019. [Google Scholar]
- Arnold, C. Alterations in the fecal microbiome of horses with antimicrobial associated diarrhea compared to antibiotic-treated and non-treated healthy case controls. In Proceedings of the 66th Annual Meeting of the American Association for Equine Practitioners, Las Vegas, NV, USA, 5–9 December 2020. [Google Scholar]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGorum, B.C.; Pirie, R.S. Antimicrobial associated diarrhoea in the horse. Part 2: Which antimicrobials are associated with AAD in the horse? Equine Vet. Educ. 2010, 22, 43–50. [Google Scholar] [CrossRef]
- Dethlefsen, L.; Relman, D.A. Incomplete recovery and individualized responses of the human distal gut microbiota to repeated antibiotic perturbation. Proc. Natl. Acad. Sci. USA 2011, 108, 4554–4561. [Google Scholar] [CrossRef] [Green Version]
- Mullish, B.H.; Williams, H.R. Clostridium difficile infection and antibiotic-associated diarrhoea. Clin. Med. 2018, 18, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Willing, B.P.; Russell, S.L.; Finlay, B.B. Shifting the balance: Antibiotic effects on host–microbiota mutualism. Nat. Rev. Microbiol. 2011, 9, 233–243. [Google Scholar] [CrossRef]
- Wipperman, M.F.; Fitzgerald, D.W.; Juste, M.A.J.; Taur, Y.; Namasivayam, S.; Sher, A.; Bean, J.M.; Bucci, V.; Glickman, M.S. Antibiotic treatment for Tuberculosis induces a profound dysbiosis of the microbiome that persists long after therapy is completed. Sci. Rep. 2017, 7, e10767. [Google Scholar] [CrossRef]
- Dougal, K.; de la Fuente, G.; Harris, P.A.; Girdwood, S.E.; Pinloche, E.; Geor, R.J.; Nielsen, B.D.; Schott, H.C., 2nd; Elzinga, S.; Newbold, C.J. Characterisation of the faecal bacterial community in adult and elderly horses fed a high fibre, high oil or high starch diet using 454 pyrosequencing. PLoS ONE 2014, 9, e87424. [Google Scholar] [CrossRef] [Green Version]
- Massacci, F.R.; Clark, A.; Ruet, A.; Lansade, L.; Costa, M.; Mach, N. Inter-breed diversity and temporal dynamics of the faecal microbiota in healthy horses. J. Anim. Breed. Genet. 2020, 137, 103–120. [Google Scholar] [CrossRef]
- Iizumi, T.; Battaglia, T.; Ruiz, V.; Perez Perez, G.I. Gut Microbiome and Antibiotics. Arch. Med. Res. 2017, 48, 727–734. [Google Scholar] [CrossRef]
- Korpela, K.; Salonen, A.; Virta, L.J.; Kekkonen, R.A.; Forslund, K.; Bork, P.; De Vos, W.M. Intestinal microbiome is related to lifetime antibiotic use in Finnish pre-school children. Nat. Commun. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Jernberg, C.; Andersson, A.F.; Sjölund-Karlsson, M.; Jansson, J.K.; Engstrand, L. Short-Term Antibiotic Treatment Has Differing Long-Term Impacts on the Human Throat and Gut Microbiome. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [Green Version]
- Vrieze, A.; Out, C.; Fuentes, S.; Jonker, L.; Reuling, I.; Kootte, R.S.; van Nood, E.; Holleman, F.; Knaapen, M.; Romijn, J.A. Impact of oral vancomycin on gut microbiota, bile acid metabolism, and insulin sensitivity. J. Hepatol. 2014, 60, 824–831. [Google Scholar] [CrossRef]
- Rashid, M.-U.; Zaura, E.; Buijs, M.J.; Keijser, B.J.; Crielaard, W.; Nord, C.E.; Weintraub, A. Determining the long-term effect of antibiotic administration on the human normal intestinal microbiota using culture and pyrosequencing methods. Clin. Infect. Dis. 2015, 60, S77–S84. [Google Scholar] [CrossRef] [Green Version]
- Stewardson, A.J.; Gaia, N.; Francois, P. Collateral damage from oral ciprofloxacin versus nitrofurantoin in outpatients with urinary tract infections: A culture-free analysis of gut microbiota. Clin. Microbiol. Infect. 2015, 21, 344. [Google Scholar] [CrossRef] [Green Version]
- Harlow, B.E.; Lawrence, L.M.; Flythe, M.D. Diarrhea-associated pathogens, lactobacilli and cellulolytic bacteria in equine feces: Responses to antibiotic challenge. Vet. Microbiol. 2013, 166, 225–232. [Google Scholar] [CrossRef]
- Harlow, B.E. Changes to the Equine Hindgut Micrflora in Response to Antibiotic Challenge. Master’s Thesis, University of Kentucky, Lexington, KY, USA, 2012. [Google Scholar]
- Gronvold, A.M.; L’Abee-Lund, T.M.; Strand, E.; Sorum, H.; Yannarell, A.C.; Mackie, R.I. Fecal microbiota of horses in the clinical setting: Potential effects of penicillin and general anesthesia. Vet. Microbiol. 2010, 145, 366–372. [Google Scholar] [CrossRef]
- Macchione, I.; Lopetuso, L.; Ianiro, G.; Napoli, M.; Gibiino, G.; Rizzatti, G.; Petito, V.; Gasbarrini, A.; Scaldaferri, F. Akkermansia muciniphila: Key player in metabolic and gastrointestinal disorders. Eur. Rev. Med. Pharmacol. Sci 2019, 23, 8075–8083. [Google Scholar]
- Lee-Sarwar, K.; Lasky-Su, J.; Kelly, R.; Litonjua, A.; Weiss, S. Metabolome–Microbiome Crosstalk and Human Disease. Metabolites 2020, 10, 181. [Google Scholar] [CrossRef]
- Guard, B.C.; Barr, J.W.; Reddivari, L.; Klemashevich, C.; Jayaraman, A.; Steiner, J.M.; Vanamala, J.; Suchodolski, J.S. Characterization of Microbial Dysbiosis and Metabolomic Changes in Dogs with Acute Diarrhea. PLoS ONE 2015, 10, e0127259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamoto, Y.; Otoni, C.C.; Steelman, S.M.; Büyükleblebici, O.; Steiner, J.M.; Jergens, A.E.; Suchodolski, J.S. Alteration of the fecal microbiota and serum metabolite profiles in dogs with idiopathic inflammatory bowel disease. Gut Microbes 2015, 6, 33–47. [Google Scholar] [CrossRef] [Green Version]
- Kolho, K.L.; Pessia, A.; Jaakkola, T.; de Vos, W.M.; Velagapudi, V. Faecal and Serum Metabolomics in Paediatric Inflammatory Bowel Disease. J. Crohns Colitis 2017, 11, 321–334. [Google Scholar] [CrossRef]
- Gupta, N.K.; Thaker, A.I.; Kanuri, N.; Riehl, T.E.; Rowley, C.W.; Stenson, W.F.; Ciorba, M.A. Serum Analysis of Tryptophan Catabolism Pathway: Correlation With Crohn’s Disease Activity. Inflamm. Bowel Dis. 2011, 18, 1214–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, S.I.; Murray, K.J.; Rendahl, A.K.; Geor, R.J.; Schultz, N.E.; McCue, M.E. Metabolic perturbations in Welsh Ponies with insulin dysregulation, obesity, and laminitis. J. Vet. Intern. Med. 2018, 32, 1215–1233. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, e3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setchell, K.D.R.; Clerici, C. Equol: History, Chemistry, and Formation. J. Nutr. 2010, 140, 1355S–1362S. [Google Scholar] [CrossRef] [Green Version]
- Saraf, M.K.; Jeng, Y.-J.; Watson, C.S. R-equol, a synthetic metabolite of the dietary estrogen daidzein, modulates the nongenomic estrogenic effects of 17β-estradiol in pituitary tumor cells. Endocr. Disruptors 2016, 4, e1226697. [Google Scholar] [CrossRef]
- Rafii, F. The Role of Colonic Bacteria in the Metabolism of the Natural Isoflavone Daidzin to Equol. Metabolites 2015, 5, 56–73. [Google Scholar] [CrossRef] [Green Version]
- Jansson, J.; Willing, B.; Lucio, M.; Fekete, A.; Dicksved, J.; Halfvarson, J.; Tysk, C.; Schmitt-Kopplin, P. Metabolomics Reveals Metabolic Biomarkers of Crohn’s Diseae. PLoS ONE 2009, 4, e6386. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Horse ID | Breed | Sex | Age | Diet Type | Group | Antibiotic | Days on ABX | Reason for ABX |
|---|---|---|---|---|---|---|---|---|
| Horse 104 | Quarter Horse | Mare | 1 | D | AAD | Doxycycline | 4 | Acute facial fracture |
| Horse 147 | Quarter Horse | Mare | 1 | D | ABX | Doxycycline | 2 | Over-riding dorsal spinous processes |
| Horse 58 | Tennessee Walker | Mare | 3 | D | CON | None | 0 | Healthy control |
| Horse 52 | Quarter Horse | Mare | 2 | D | CON | None | 0 | Healthy control |
| Horse 148 | Quarter Horse | Gelding | 9 | C | AAD | Doxycycline | 7 | Pemanent tracheostomy |
| Horse 149 | Quarter Horse | Gelding | 12 | C | ABX | Doxycycline | 8 | Wound revision |
| Horse 150 | Mix | Gelding | 8 | C | CON | None | 0 | Healthy control |
| Horse 45 | Warmblood | Gelding | 7 | C | CON | None | 0 | Healthy control |
| Horse 151 | Quarter Horse | Gelding | 6 | B | AAD | Doxycycline | 5 | Tie-forward |
| Horse 152 | Quarter Horse | Gelding | 15 | B | ABX | Doxycycline | 7 | Tenoscopy |
| Horse 36 | Quarter Horse | Gelding | 12 | B | CON | None | 0 | Healthy control |
| Horse 48 | Mix | Gelding | 7 | B | CON | None | 0 | Healthy control |
| Horse 108 | Quarter Horse | Stallion | 1 | C | AAD | Excede | 10 | Pneumonia |
| Horse 153 | Miniature Donkey | Stallion | 1 | C | AAD | Excede | 7 | Pneumonia |
| Horse 154 | Thoroughbred | Gelding | 12 | C | ABX | Excede | 5 | Dental/sinus disease |
| Horse 42 | Warmblood | Gelding | 6 | C | CON | None | 0 | Healthy control |
| Horse 46 | Quarter Horse | Gelding | 2 | C | CON | None | 0 | Healthy control |
| Horse 102 | Quarter Horse | Gelding | 9 | D | AAD | Excede | 3 | Pneumonia |
| Horse 155 | Paint | Stallion | 1 | D | AAD | Excede | 2 | Pneumonia |
| Horse 156 | Quarter Horse | Mare | 19 | D | ABX | Excede | 4 | Oocyte aspiration |
| Horse 61 | Tennessee Walking Horse | Stallion | 8 | D | CON | None | 0 | Healthy control |
| Horse 63 | Draft cross | Gelding | 9 | D | CON | None | 0 | Healthy control |
| Horse 111 | Paint | Mare | 11 | D | AAD | Penicillin, Gentocin | 4 | Mass removal |
| Horse 159 | Quarter Horse | Mare | 8 | D | ABX | Penicillin, Gentocin | 3 | Fetlock arthrodesis |
| Horse 55 | Warmblood | Mare | 8 | D | CON | None | 0 | Healthy control |
| Horse 60 | Quarter Horse | Mare | 14 | D | CON | None | 0 | Healthy control |
| Horse 106 | Quarter Horse | Stallion | 5 | C | AAD | Penicillin, Gentocin | 4 | Acute facial fracture |
| Horse 161 | Warmblood | Mare | 12 | C | ABX | Penicillin, Gentocin | 4 | Pastern arthrodesis |
| Horse 162 | Quarter Horse | Stallion | 3 | C | CON | None | 0 | Healthy control |
| Horse 163 | Quarter Horse | Stallion | 6 | C | CON | None | 0 | Healthy control |
| Horse 164 | Warmblood | Gelding | 1 | C | AAD | Penicillin, Gentocin | 1 | Cryptorchid |
| Horse 165 | Quarter Horse | Gelding | 2 | C | ABX | Penicillin, Gentocin | 1 | Stifle arthroscopy |
| Horse 166 | Quarter Horse | Gelding | 1 | C | CON | None | 0 | Healthy control |
| Horse 28 | Warmblood | Gelding | 2 | C | CON | None | 0 | Healthy control |
| Horse 103 | Quarter Horse | Stallion | 1 | D | AAD | Penicillin, Gentocin | 3 | Inferior check ligament desmotomy |
| Horse167 | Quarter Horse | Stallion | 1 | D | ABX | Penicillin, Gentocin | 3 | Cryptorchid |
| Horse 61 | Quarter Horse | Stallion | 3 | D | CON | None | 0 | Healthy control |
| Horse 168 | Draft | Stallion | 1 | D | CON | None | 0 | Healthy control |
| Horse 116 | Mixed | Gelding | 1 | B | AAD | Penicillin, Gentocin, Doxycycline | 7 | Pneumonia |
| Horse 169 | Saddlebred | Stallion | 1 | B | ABX | Penicillin, Gentocin, Doxycycline | 7 | Radial fracture |
| Horse 170 | Quarter Horse | Gelding | 7 | B | CON | None | 0 | Healthy control |
| Horse 171 | Tennessee Walker | Stallion | 6 | B | CON | None | 0 | Healthy control |
| Horse 105 | Warmblood | Gelding | 12 | C | AAD | Penicillin, Gentocin, Doxycycline | 7 | Fasciotomy/neurectomy |
| Horse 172 | Paint | Gelding | 10 | C | ABX | Penicillin, Gentocin, Doxycycline | 7 | Ulnar fracture |
| Horse 40 | Quarter Horse | Gelding | 8 | C | CON | None | 0 | Healthy control |
| Horse 178 | Warmblood | Gelding | 16 | C | CON | None | 0 | Healthy control |
| Horse 113 | Quarter Horse | Mare | 12 | B | AAD | Penicillin, Gentocin, Metronidazole | 3 | Pneumonia |
| Horse 173 | Warmblod | Gelding | 19 | B | ABX | Penicillin, Gentocin, Metronidazole | 4 | Pneumonia |
| Horse 30 | Quarter Horse | Mare | 13 | B | CON | None | 0 | Healthy control |
| Horse 25 | Quarter Horse | Mare | 12 | B | CON | None | 0 | Healthy control |
| Horse 107 | Thoroughbred | Mare | 25 | D | AAD | Penicillin, Gentocin, Metronidazole | 4 | Pneumonia |
| Horse 174 | Criollo | Mare | 19 | D | ABX | Penicillin, Gentocin, Metronidazole | 4 | Acute facial trauma |
| Horse 175 | Quarter Horse | Mare | 24 | D | CON | None | 0 | Healthy control |
| Horse 59 | Thoroughbred | Mare | 12 | D | CON | None | 0 | Healthy control |
| Horse 176 | Miniature Horse | Gelding | 26 | D | AAD | TMS | 7 | Dental/sinus disease |
| Horse 177 | Miniature Horse | Gelding | 23 | D | ABX | TMS | 14 | Dental/sinus disease |
| Horse 62 | Appoloosa | Gelding | 21 | D | CON | None | 0 | Healthy control |
| Horse 63 | Thoroughbred | Gelding | 24 | D | CON | None | 0 | Healthy control |
| Horse 56 | Quarter Horse | Gelding | 19 | D | CON | None | 0 | Healthy control |
| Group | Unweighted | Weighted | ||
|---|---|---|---|---|
| R | p-Value | R | p-Value | |
| Overall | 0.398 | 0.001 | 0.391 | 0.001 |
| AAD vs. ABX | 0.063 | 0.078 | 0.121 | 0.0012 |
| AAD vs. CON | 0.547 | 0.001 | 0.568 | 0.001 |
| ABX vs. CON | 0.37 | 0.001 | 0.3 | 0.001 |
| Bacterial Phyla | AAD | ABX | CON | AAD vs. ABX vs. CON | ||||
|---|---|---|---|---|---|---|---|---|
| Median | Range | Median | Range | Median | Range | p-Value | Q-Value | |
| Euryarchaeota | 0.98 | 0.1–2.12 | 0.84 | 0.07–1.98 | 0.78 | 0.02–1.43 | 0.4535 | 0.4535 |
| Unknown | 0 | 0–0.04 | 0.01 | 0–0.03 | 0 | 0–0.03 | 0.25 | 0.2632 |
| Actinobacteria | 0.36 a | 0.08–1.94 | 0.66 a,b | 0.18–1.57 | 0.9 b | 0.25–4.6 | 0.0115 | 0.0192 |
| Armatimonadetes | 0 a | 0–0.02 | 0 a,b | 0–0.07 | 0.01 b | 0–0.13 | 0.0032 | 0.008 |
| Bacteroidetes | 43.88 a | 24.16–66.11 | 40.43 a,b | 31.59–48.07 | 36.46 b | 9.29–47.6 | 0.0001 | 0.0005 |
| Elusimicrobia | 0 a | 0–0.09 | 0 a | 0–0.14 | 0.02 b | 0–0.14 | 0.0001 | 0.0005 |
| Fibrobacteres | 0.38 a | 0.06–4.38 | 0.89 a,b | 0.1–6.65 | 1.51 b | 0.18–8.75 | 0.0492 | 0.0656 |
| Firmicutes | 36.45 | 21.78–54.83 | 40.8 | 28.59–58.12 | 41.68 | 29.81–68.78 | 0.2058 | 0.2287 |
| Fusobacteria | 0.01 a,b | 0–8.36 | 0.05 a | 0–0.1 | 0 b | 0–0.08 | 0.0214 | 0.0306 |
| Lentisphaerae | 0.02 | 0–0.37 | 0.07 | 0.01–0.31 | 0.05 | 0–0.61 | 0.1373 | 0.1615 |
| Planctomycetes | 0.02 a | 0–0.07 | 0.04 a | 0–0.39 | 0.09 b | 0.02–0.41 | 0.0002 | 0.0008 |
| Proteobacteria | 2.93 | 0.88–14.32 | 1.97 | 0.28–25.99 | 1.59 | 0.37–10 | 0.0695 | 0.0860 |
| SR1 | 0 a | 0–0.02 | 0 a | 0–0.02 | 0.03 b | 0–0.38 | 0.0001 | 0.0005 |
| Spirochaetes | 4.07 a | 0.77–9.96 | 4.41 a,b | 0.72–10.46 | 6.03 b | 2.44–10.39 | 0.0137 | 0.0211 |
| Synergistetes | 0.02 a | 0–0.17 | 0.03 a,b | 0–0.29 | 0.04 b | 0.02–0.17 | 0.0114 | 0.0192 |
| TM7 | 0 a | 0–0.01 | 0 a,b | 0–0.05 | 0.02 b | 0–0.14 | 0.0075 | 0.01667 |
| Tenericutes | 1.5 a,b | 0.18–9.72 | 1.11 a | 0.34–2.52 | 2.44 b | 1.24–5.7 | 0.0004 | 0.00133 |
| Verrucomicrobia | 1.15 a | 0.31–6.93 | 5.23 b | 0.24–14.55 | 7.78 b | 2.9–14.02 | 0.0001 | 0.0005 |
| WPS-2 | 0 a,b | 0–0.01 | 0 a | 0–0.02 | 0.01 b | 0–0.31 | 0.0109 | 0.01917 |
| Group | Metabolite | p-Value | FDR | Perturbation to CON | Source | KEGG Pathway | Specific Pathway |
|---|---|---|---|---|---|---|---|
| AAD vs. CON | N-acetyltyramine | 0.0017 | 0.0262 | Increased in AAD | Endogenous | Amino acid metabolism | Precursor for L-tyrosine |
| 5-hydroxyindole-3-acetic acid | 0.0022 | 0.0290 | Increase in AAD | Gut microbiota | Amino acid metabolism | Tyrptophan, indole pathway | |
| AAD, ABX vs. CON | Kynurenic acid | 0.0000 | 0.0004 | Increased in AAD, ABX | Gut microbiota | Amino acid metabolism | Tryptophan, kynurenine pathway |
| Docosahexaenoic acid | 0.0015 | 0.0262 | Increased in AAD, ABX | Dietary | Lipid metabolism | Omega-3 essential fatty acid | |
| Xanthurenic acid | 0.0033 | 0.0348 | Increased in AAD, ABX | Gut microbiota | Amino acid metabolism | Tryptophan, kynurenine pathway | |
| AAD vs. ABX, CON | Daidzein | 0.0009 | 0.0215 | Increased in AAD | Gut microbiota | Biosynthesis of secondary metabolites | Isoflavone metabolite, estrogenic compound |
| L-tyrosine | 0.0030 | 0.0348 | Increase in AAD | Dietary | Amino acid metabolism | Precursors for dopamine, norepinephrine, and epinephrine |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arnold, C.; Pilla, R.; Chaffin, K.; Lidbury, J.; Steiner, J.; Suchodolski, J. Alterations in the Fecal Microbiome and Metabolome of Horses with Antimicrobial-Associated Diarrhea Compared to Antibiotic-Treated and Non-Treated Healthy Case Controls. Animals 2021, 11, 1807. https://doi.org/10.3390/ani11061807
Arnold C, Pilla R, Chaffin K, Lidbury J, Steiner J, Suchodolski J. Alterations in the Fecal Microbiome and Metabolome of Horses with Antimicrobial-Associated Diarrhea Compared to Antibiotic-Treated and Non-Treated Healthy Case Controls. Animals. 2021; 11(6):1807. https://doi.org/10.3390/ani11061807
Chicago/Turabian StyleArnold, Carolyn, Rachel Pilla, Keith Chaffin, Jonathan Lidbury, Joerg Steiner, and Jan Suchodolski. 2021. "Alterations in the Fecal Microbiome and Metabolome of Horses with Antimicrobial-Associated Diarrhea Compared to Antibiotic-Treated and Non-Treated Healthy Case Controls" Animals 11, no. 6: 1807. https://doi.org/10.3390/ani11061807