
Lipofection-Mediated Introduction of CRISPR/Cas9 System into Porcine Oocytes and Embryos

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Oocyte Collection and In Vitro Maturation
2.3. IVF and Embryo Culture
2.4. Lipofection-Mediated Introduction of CRISPR/Cas9 System
2.5. Analysis of the Targeted Gene in Embryos
2.6. Experimental Design
2.6.1. Experiment 1: Lipofection-Mediated Introduction of CRISPR/Cas9 System into Oocytes during IVF
2.6.2. Experiment 2: Lipofection-Mediated Introduction of CRISPR/Cas9 System into Embryos
2.6.3. Experiment 3: Comparison of Lipofection-Mediated Gene-Editing Efficiency among Embryos at Different Cleavage Stages
2.7. Statistical Analyses
3. Results
3.1. Experiment 1
3.2. Experiment 2
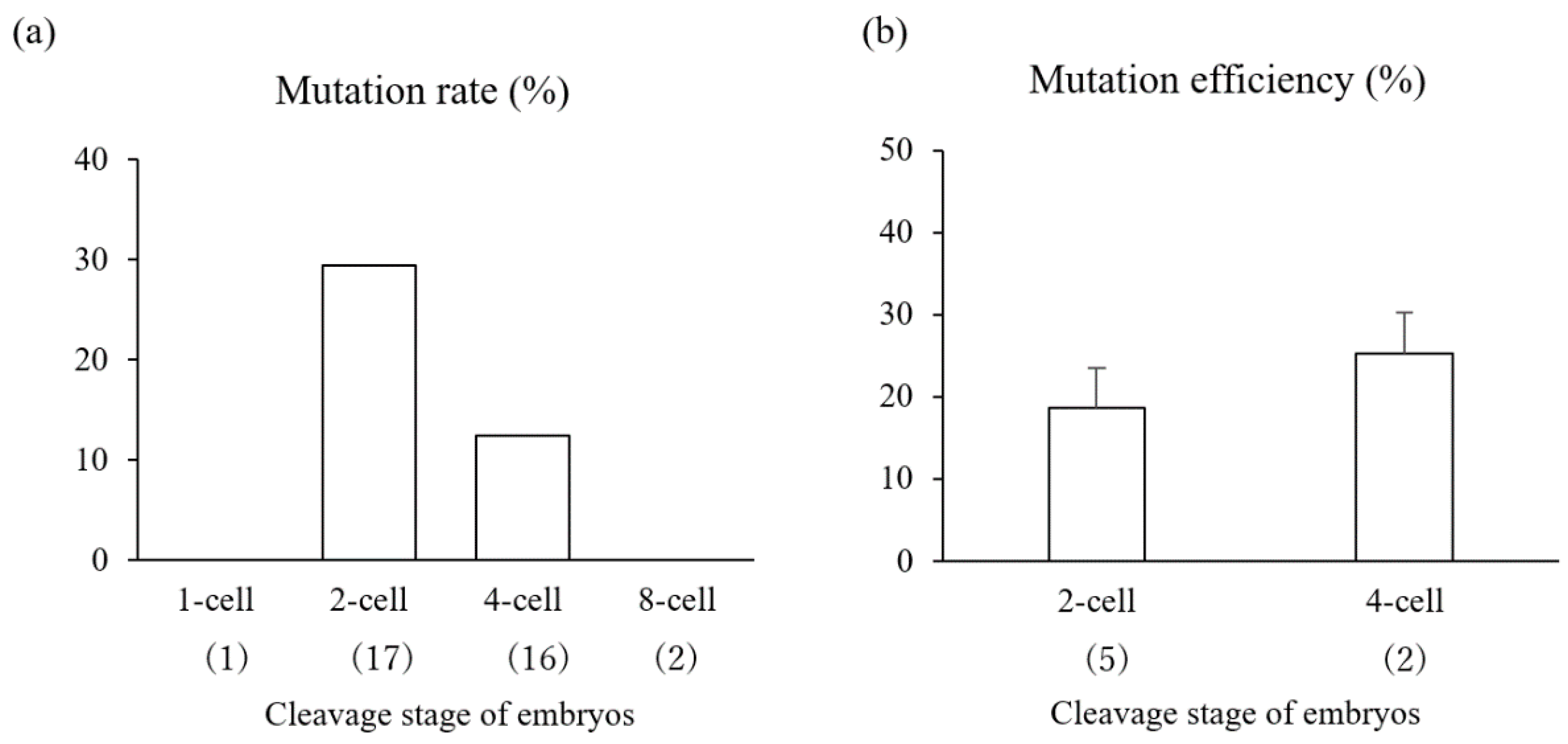
3.3. Experiment 3
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, H.H.; Zhao, H.; Qing, Y.B.; Pan, W.R.; Jia, B.Y.; Zhao, H.Y.; Huang, X.X.; Wei, H.J. Porcine Zygote Injection with Cas9/sgRNA Results in DMD-Modified Pig with Muscle Dystrophy. Int. J. Mol. Sci. 2016, 17, 1668. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Du, Y.; Shen, B.; Zhou, X.; Li, J.; Liu, Y.; Wang, J.; Zhou, J.; Hu, B.; Kang, N.; et al. Efficient generation of gene-modified pigs via injection of zygote with Cas9/sgRNA. Sci. Rep. 2015, 5, 8256. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Shen, L.; Chen, J.; Liu, X.; Tan, T.; Hu, Y.; Bai, X.; Li, Y.; Tian, K.; Li, N.; et al. Deletion of CD163 Exon 7 Confers Resistance to Highly Pathogenic Porcine Reproductive and Respiratory Viruses on Pigs. Int. J. Biol. Sci. 2019, 15, 1993–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanihara, F.; Takemoto, T.; Kitagawa, E.; Rao, S.; Do, L.T.; Onishi, A.; Yamashita, Y.; Kosugi, C.; Suzuki, H.; Sembon, S.; et al. Somatic cell reprogramming-free generation of genetically modified pigs. Sci. Adv. 2016, 2, e1600803. [Google Scholar] [CrossRef] [Green Version]
- Abe, A.; Miyanohara, A.; Friedmann, T. Polybrene increases the efficiency of gene transfer by lipofection. Gene Ther. 1998, 5, 708–711. [Google Scholar] [CrossRef] [Green Version]
- Felgner, P.L.; Gadek, T.R.; Holm, M.; Roman, R.; Chan, H.W.; Wenz, M.; Northrop, J.P.; Ringold, G.M.; Danielsen, M. Lipofection: A highly efficient, lipid-mediated DNA-transfection procedure. Proc. Natl. Acad. Sci. USA 1987, 84, 7413–7417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, A.; Lakey, J.; Murray, A.; Moore, R. Gene therapy: A lipofection approach for gene transfer into primary endothelial cells. Cell Transplant. 2002, 11, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Zeitlin, P.L.; Guggino, W.B.; Craig, R.W. Gene transfer by lipofection in rabbit and human secretory epithelial cells. Pflügers Arch. 1989, 415, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Lascombe, I.; Mougin, P.; Vuillermoz, C.; Adessi, G.; Jouvenot, M. Gene transfer into subcultured endometrial cells using lipofection. Biotechniques 1996, 20, 88–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochelen, D.; Eclancher, F.; Kupferberg, A.; Privat, A.; Mersel, M. 7β-hydroxycholesterol and 7β-hydroxycholesteryl-3-esters reduce the extent of reactive gliosis caused by an electrolytic lesion in rat brain. Neuroscience 1992, 51, 827–834. [Google Scholar] [CrossRef]
- Yu, A.; Lee, Y.; Eng, L. Inhibition of GFAP synthesis by antisense RNA in astrocytes. J. Neurosci. Res. 1991, 30, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Yang, N.S.; Jiao, S.; Sun, J.; Cheng, L.; Wolff, J.; Duncan, I. Efficient and sustained transgene expression in mature rat oligodendrocytes in primary culture. J. Neurosci. Res. 1996, 43, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Brazolot, C.; Petitte, J.; Etches, R.; Gibbins, A.V. Efficient transfection of chicken cells by lipofection, and introduction of transfected blastodermal cells into the embryo. Mol. Reprod. Dev. 1991, 30, 304–312. [Google Scholar] [CrossRef]
- Holt, C.E.; Garlick, N.; Cornel, E. Lipofection of cDNAs in the embryonic vertebrate central nervous system. Neuron 1990, 4, 203–214. [Google Scholar] [CrossRef]
- McLenachan, S.; Sarsero, J.P.; Ioannou, P.A. Flow-cytometric analysis of mouse embryonic stem cell lipofection using small and large DNA constructs. Genomics 2007, 89, 708–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuris, J.A.; Thompson, D.B.; Shu, Y.; Guilinger, J.P.; Bessen, J.L.; Hu, J.H.; Maeder, M.L.; Joung, J.K.; Chen, Z.Y.; Liu, D.R. Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo. Nat. Biotechnol. 2015, 33, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.V.; Tanihara, F.; Do, L.; Sato, Y.; Taniguchi, M.; Takagi, M.; Van Nguyen, T.; Otoi, T. Chlorogenic acid supplementation during in vitro maturation improves maturation, fertilization and developmental competence of porcine oocytes. Reprod. Domest. Anim. 2017, 52, 969–975. [Google Scholar] [CrossRef]
- Brinkman, E.K.; Kousholt, A.N.; Harmsen, T.; Leemans, C.; Chen, T.; Jonkers, J.; van Steensel, B. Easy quantification of template-directed CRISPR/Cas9 editing. Nucleic Acids Res. 2018, 46, e58. [Google Scholar] [CrossRef] [PubMed]
- Tanihara, F.; Hirata, M.; Thi Nguyen, N.; Anh Le, Q.; Hirano, T.; Otoi, T. Generation of viable PDX1 gene-edited founder pigs as providers of nonmosaics. Mol. Reprod. Dev. 2020, 87, 471–481. [Google Scholar] [CrossRef]
- Tanihara, F.; Hirata, M.; Nguyen, N.T.; Sawamoto, O.; Kikuchi, T.; Doi, M.; Otoi, T. Efficient generation of GGTA1-deficient pigs by electroporation of the CRISPR/Cas9 system into in vitro-fertilized zygotes. BMC Biotechnol. 2020, 20, 40. [Google Scholar] [CrossRef]
- Akita, H.; Ito, R.; Khalil, I.; Futaki, S.; Harashima, H. Quantitative three-dimensional analysis of the intracellular trafficking of plasmid DNA transfected by a nonviral gene delivery system using confocal laser scanning microscopy. Mol. Ther. 2004, 9, 443–451. [Google Scholar] [CrossRef]
- Cardarelli, F.; Digiacomo, L.; Marchini, C.; Amici, A.; Salomone, F.; Fiume, G.; Rossetta, A.; Gratton, E.; Pozzi, D.; Caracciolo, G. The intracellular trafficking mechanism of Lipofectamine-based transfection reagents and its implication for gene delivery. Sci. Rep. 2016, 6, 25879. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Sunshine, J.C.; Green, J.J. Differential polymer structure tunes mechanism of cellular uptake and transfection routes of poly (β-amino ester) polyplexes in human breast cancer cells. Bioconj. Chem. 2014, 25, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maneiro, E.; Ron-Corzo, A.; Julve, J.; Goyanes, V.J. Surface area/volume ratio and growth equation of the human early embryo. Int. J. Dev. Biol. 2002, 35, 139–143. [Google Scholar]
- Pereira, R.; Marques, C. Animal oocyte and embryo cryopreservation. Cell Tissue Bank. 2008, 9, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Kosuke, M.; Koriyama, M.; Inada, E.; Saitoh, I.; Ohtsuka, M.; Nakamura, S.; Sakurai, T.; Watanabe, S.; Miyoshi, K. Timing of CRISPR/Cas9-related mRNA microinjection after activation as an important factor affecting genome editing efficiency in porcine oocytes. Theriogenology 2018, 108, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Tao, L.; Yang, M.; Wang, X.; Zhang, Z.; Wu, Z.; Tian, J.; An, L.; Wang, S. Efficient biallelic mutation in porcine parthenotes using a CRISPR-Cas9 system. Biochem. Biophys. Res. Commun. 2016, 476, 225–229. [Google Scholar] [CrossRef]
- Gu, B.; Posfai, E.; Rossant, J. Efficient generation of targeted large insertions by microinjection into two-cell-stage mouse embryos. Nat. Biotechnol. 2018, 36, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Plaza Reyes, A.; Lanner, F. Time Matters: Gene Editing at the Mouse 2-Cell Embryo Stage Boosts Knockin Efficiency. Cell Stem Cell 2018, 23, 155–157. [Google Scholar] [CrossRef] [Green Version]
- Hyttel, P.; Laurincik, J.; Rosenkranz, C.; Rath, D.; Niemann, H.; Ochs, R.L.; Schellander, K. Nucleolar proteins and ultrastructure in preimplantation porcine embryos developed in vivo. Biol. Reprod. 2000, 63, 1848–1856. [Google Scholar] [CrossRef] [Green Version]
- Ostrup, O.; Olbricht, G.; Ostrup, E.; Hyttel, P.; Collas, P.; Cabot, R. RNA profiles of porcine embryos during genome activation reveal complex metabolic switch sensitive to in vitro conditions. PLoS ONE 2013, 8, e61547. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Tu, L.C.; Naseri, A.; Huisman, M.; Zhang, S.; Grunwald, D.; Pederson, T. CRISPR-Cas9 nuclear dynamics and target recognition in living cells. J. Cell Biol. 2016, 214, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Burkard, C.; Lillico, S.G.; Reid, E.; Jackson, B.; Mileham, A.J.; Ait-Ali, T.; Whitelaw, C.B.A.; Archibald, A.L. Precision engineering for PRRSV resistance in pigs: Macrophages from genome edited pigs lacking CD163 SRCR5 domain are fully resistant to both PRRSV genotypes while maintaining biological function. PLoS Pathog. 2017, 13, e1006206. [Google Scholar] [CrossRef]
- Vera-Rodriguez, M.; Rubio, C. Assessing the true incidence of mosaicism in preimplantation embryos. Fertil. Steril. 2017, 107, 1107–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilarino, M.; Suchy, F.P.; Rashid, S.T.; Lindsay, H.; Reyes, J.; McNabb, B.R.; van der Meulen, T.; Huising, M.O.; Nakauchi, H.; Ross, P.J. Mosaicism diminishes the value of pre-implantation embryo biopsies for detecting CRISPR/Cas9 induced mutations in sheep. Transgenic Res. 2018, 27, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-H.; Abeydeera, L.R.; Han, Y.-M.; Prather, R.S.; Day, B.N. Morphologic evaluation and actin filament distribution in porcine embryos produced in vitro and in vivo. Biol. Reprod. 1999, 60, 1020–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, Q.A.; Tanihara, F.; Wittayarat, M.; Namula, Z.; Sato, Y.; Lin, Q.; Takebayashi, K.; Hirata, M.; Otoi, T. Comparison of the effects of introducing the CRISPR/Cas9 system by microinjection and electroporation into porcine embryos at different stages. BMC Res. Notes 2021, 14, 7. [Google Scholar] [CrossRef]
- Tanihara, F.; Hirata, M.; Nguyen, N.T.; Le, Q.A.; Hirano, T.; Otoi, T. Effects of concentration of CRISPR/Cas9 components on genetic mosaicism in cytoplasmic microinjected porcine embryos. J. Reprod. Dev. 2019, 65, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishio, K.; Tanihara, F.; Nguyen, T.V.; Kunihara, T.; Nii, M.; Hirata, M.; Takemoto, T.; Otoi, T. Effects of voltage strength during electroporation on the development and quality of in vitro-produced porcine embryos. Reprod. Domest. Anim. 2018, 53, 313–318. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| gRNA †† | No. of Embryos Examined | No. (Mean ± SEM) of Embryos Developed to Blastocysts | No. of Blastocysts Examined | No. (Mean) of Blastocysts ††† | Mutation Efficiency (Mean ± SEM) †††† | ||
|---|---|---|---|---|---|---|---|
| WT | Biallelic | Mosaic | |||||
| Control | 202 | 24 (11.9 ± 0.4) | - | - | - | - | - |
| PDX1 | 395 | 44 (9.5 ± 2.6) | 35 | 32 (91.4) | 0 (0) | 3 (8.6) | 35.8 ±10.5 |
| GGTA1 | 272 | 39 (13.8 ± 6.0) | 39 | 33 (84.6) | 0 (0) | 6 (15.4) | 17.8 ± 3.0 |
| Post-IVF Exposure Period †† | No. of Embryos Examined | No. (Mean ± SEM) of Embryos Developed to Blastocysts | No. of Blastocysts Examined | No. (Mean) of Blastocysts ††† | Mutation Efficiency (Mean ± SEM) †††† | ||
|---|---|---|---|---|---|---|---|
| WT | Biallelic | Mosaic | |||||
| Control | 210 | 45 (21.4 ± 2.2) a | - | - | - | - | - |
| 24–29 h | 250 | 21 (8.1 ± 1.5) b | 18 | 8 (44.4) | 0 (0) | 10 (55.6) | 45.3 ± 9.9 |
| 29–34 h | 250 | 20 (8.1 ± 0.9) b | 14 | 6 (42.9) | 1 (7.1) | 7 (50.0) | 40.8 ± 10.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirata, M.; Wittayarat, M.; Namula, Z.; Anh Le, Q.; Lin, Q.; Takebayashi, K.; Thongkittidilok, C.; Tanihara, F.; Otoi, T. Lipofection-Mediated Introduction of CRISPR/Cas9 System into Porcine Oocytes and Embryos. Animals 2021, 11, 578. https://doi.org/10.3390/ani11020578
Hirata M, Wittayarat M, Namula Z, Anh Le Q, Lin Q, Takebayashi K, Thongkittidilok C, Tanihara F, Otoi T. Lipofection-Mediated Introduction of CRISPR/Cas9 System into Porcine Oocytes and Embryos. Animals. 2021; 11(2):578. https://doi.org/10.3390/ani11020578
Chicago/Turabian StyleHirata, Maki, Manita Wittayarat, Zhao Namula, Quynh Anh Le, Qingyi Lin, Koki Takebayashi, Chommanart Thongkittidilok, Fuminori Tanihara, and Takeshige Otoi. 2021. "Lipofection-Mediated Introduction of CRISPR/Cas9 System into Porcine Oocytes and Embryos" Animals 11, no. 2: 578. https://doi.org/10.3390/ani11020578