Molecular Identification and mRNA Expression Profiles of Galectin-9 Gene in Red Sea Bream (Pagrus major) Infected with Pathogens

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification and Characterization of the Galectin-9 Gene
2.2. Experimental Fish and Microorganisms
2.3. Tissue Collection and Processing
2.4. Quantitative Real-Time PCR
3. Results
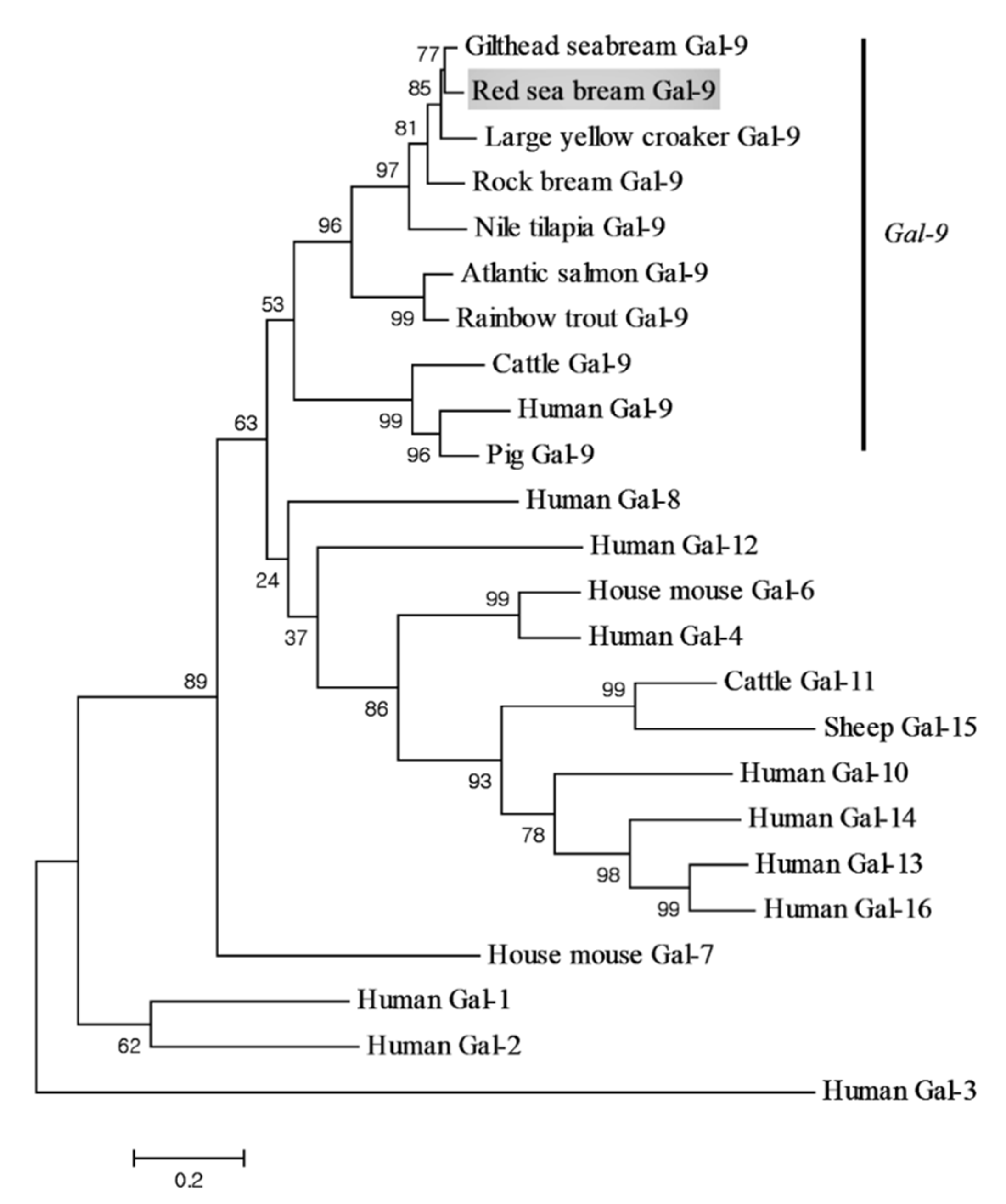
3.1. Identification and Characterization of PmGal-9 Sequence
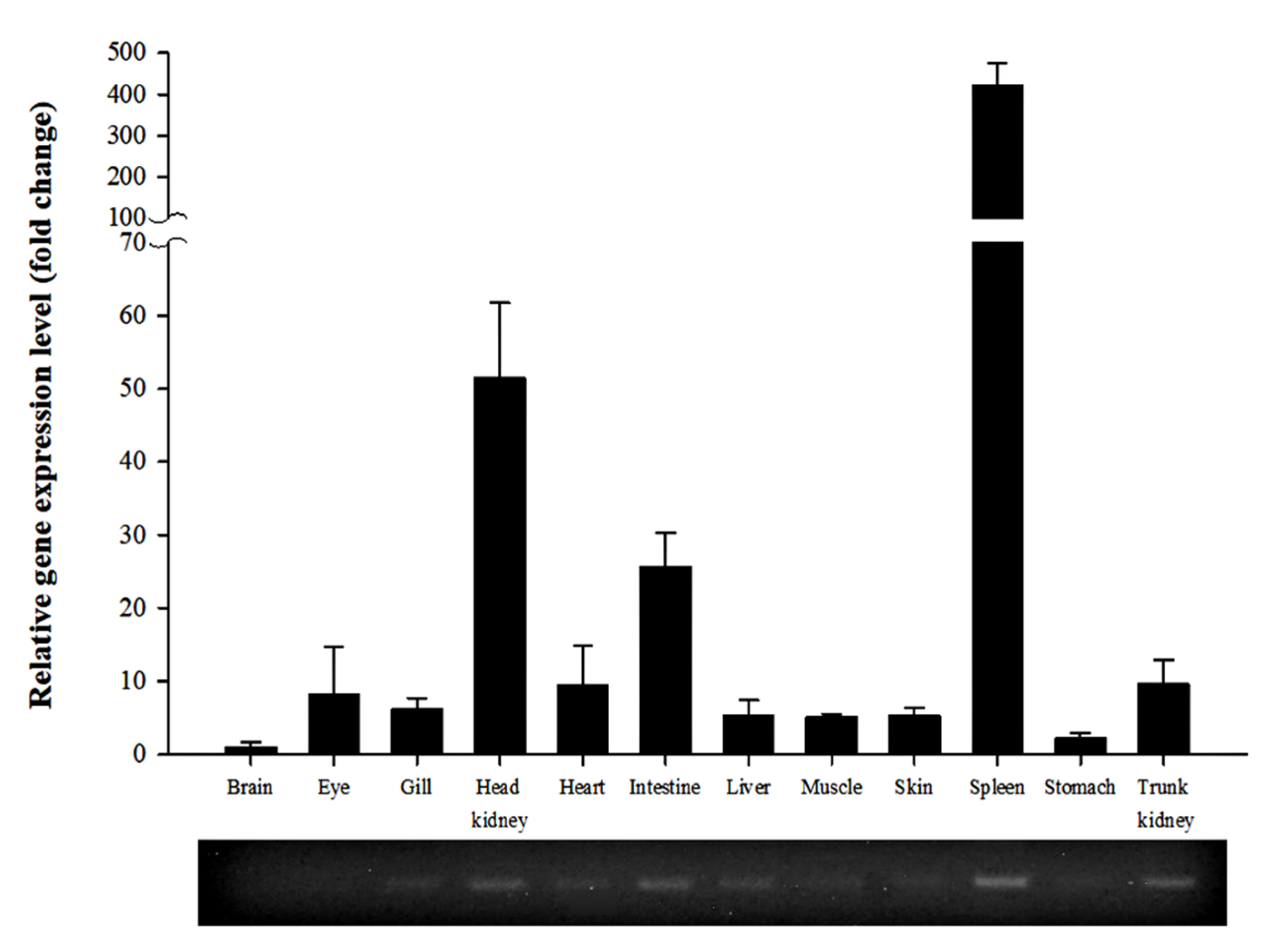
3.2. Expression of PmGal-9 mRNA in Various Tissues
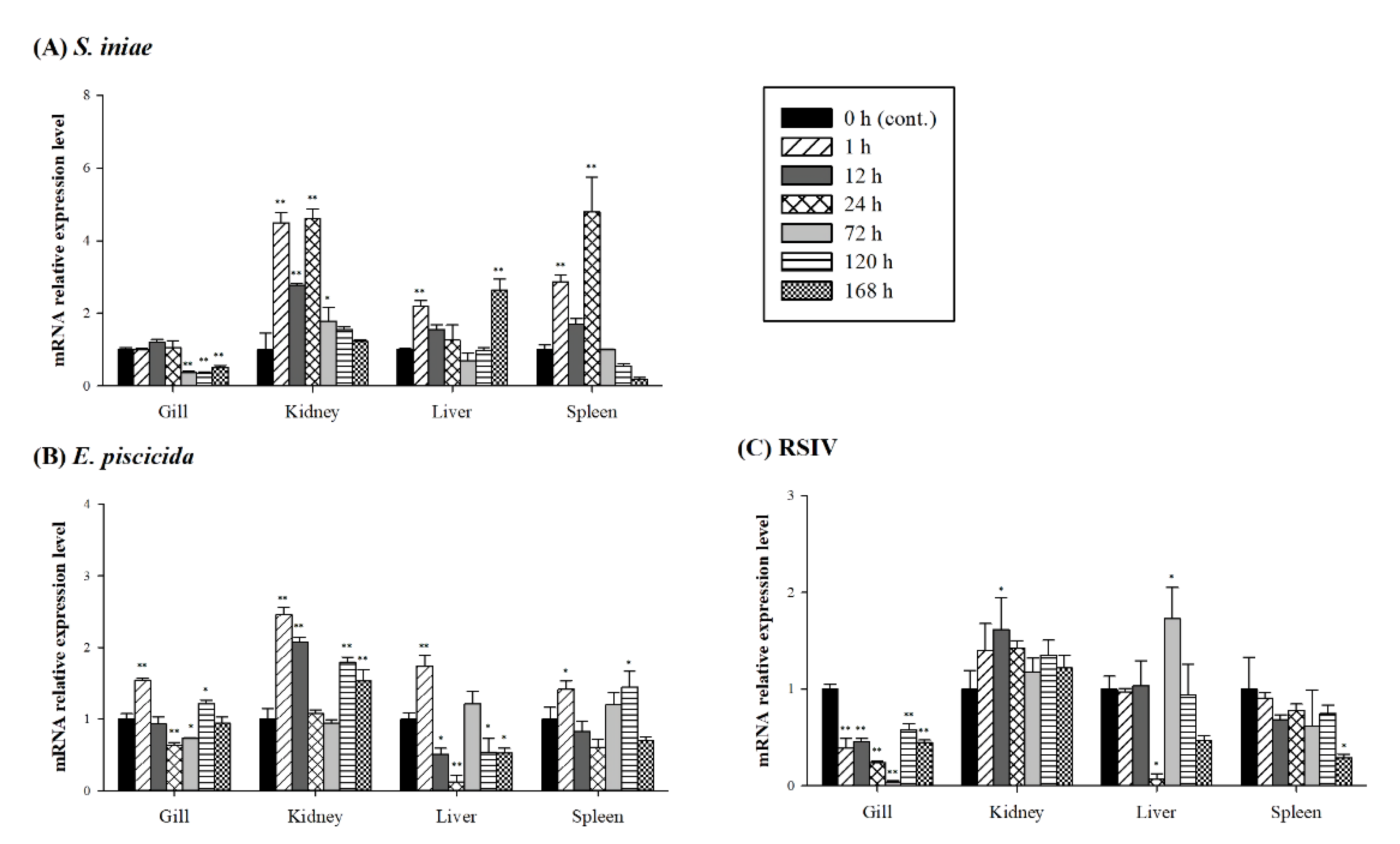
3.3. Expression of PmGal-9 mRNA after Pathogen Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barondes, S.H.; Cooper, D.N.; Gitt, M.A.; Leffler, H. Galectins. Structure and function of a large family of animal lectins. J. Biol. Chem. 1994, 269, 20807–20810. [Google Scholar] [PubMed]
- Mehul, B.; Bawumia, S.; Martin, S.R.; Hughes, R.C. Structure of baby hamster kidney carbohy-drate-binding protein CBP30, an S-type animal lectin. J. Biol. Chem. 1994, 269, 18250–18258. [Google Scholar] [PubMed]
- Vasta, G.R. Roles of galectins in infection. Nat. Rev. Genet. 2009, 7, 424–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, R.C. Galectins as modulators of cell adhesion. Biochimie 2001, 83, 667–676. [Google Scholar] [CrossRef]
- Boscher, C.; Zheng, Y.Z.; Lakshminarayan, R.; Johannes, L.; Dennis, J.W.; Foster, L.J.; Nabi, I.R. Galectin-3 Protein Regulates Mobility of N-cadherin and GM1 Ganglioside at cell-cell junctions of mammary carcinoma cells. J. Biol. Chem. 2012, 287, 32940–32952. [Google Scholar] [CrossRef] [Green Version]
- Johannes, L.; Jacob, R.; Leffler, H. Galectins at a glance. J. Cell Sci. 2018, 131, jcs208884. [Google Scholar] [CrossRef] [Green Version]
- Hirabayashi, J.; Kasai, K.-I. The family of metazoan metal-independent β-galactoside-binding lectins: Structure, function and molecular evolution. Glycobiology 1993, 3, 297–304. [Google Scholar] [CrossRef]
- Wada, J.; Kanwar, Y.S. Identification and characterization of galectin-9, a novel β-galactoside-binding mammalian lectin. J. Biol. Chem. 1997, 272, 6078–6086. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Anderson, A.C.; Schubart, A.; Xiong, H.; Imitola, J.; Khoury, S.J.; Zheng, X.X.; Strom, T.B.; Kuchroo, V.K. The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity. Nat. Immunol. 2005, 6, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, N.; Kozuru, H.; Koga, T.; Abiru, S.; Yamasaki, K.; Komori, A.; Fujita, Y.; Tenmoku, J.; Asano, T.; Sato, S.; et al. Galectin-9 in autoimmune hepatitis. Medicine 2019, 98, e16924. [Google Scholar] [CrossRef]
- Panda, S.K.; Facchinetti, V.; Voynova, E.; Hanabuchi, S.; Karnell, J.L.; Hanna, R.N.; Kolbeck, R.; Sanjuan, M.A.; Ettinger, R.; Liu, Y.-J. Galectin-9 inhibits TLR7-mediated autoimmunity in murine lupus models. J. Clin. Investig. 2018, 128, 1873–1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, A.; Kontani, K.; Kihara, M.; Nishi, N.; Yokomise, H.; Hirashima, M. Galectin-9, a novel prognostic factor with antimetastatic potential in breast cancer. Breast J. 2006, 12, S196–S200. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.-H.; Luo, S.F.; Wang, M.-Y.; Ho, L.-J. translational implication of galectin-9 in the pathogenesis and treatment of viral infection. Int. J. Mol. Sci. 2017, 18, 2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Mohsen, M.; Chavez, L.; Tandon, R.; Chew, G.M.; Deng, X.; Danesh, A.; Keating, S.; Lanteri, M.; Samuels, M.L.; Hoh, R.; et al. Human galectin-9 is a potent mediator of hiv transcription and reactivation. PLoS Pathog. 2016, 12, e1005677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.-Y.; Dong, J.-H.; Chen, Y.-W.; Wang, X.-Q.; Li, C.-H.; Wang, J.; Wang, G.-Q.; Li, H.-L.; Wang, X.-D. Galectin-9 Acts as a prognostic factor with antimetastatic potential in hepatocellular carcinoma. Asian Pac. J. Cancer Prev. 2012, 13, 2503–2509. [Google Scholar] [CrossRef]
- Irie, A.; Yamauchi, A.; Kontani, K.; Kihara, M.; Liu, D.; Shirato, Y.; Seki, M.; Nishi, N.; Nakamura, T.; Yokomise, H.; et al. Galectin-9 as a prognostic factor with antimetastatic potential in breast cancer. Clin. Cancer Res. 2005, 11, 2962–2968. [Google Scholar] [CrossRef] [Green Version]
- O’Farrell, C.; Vaghefi, N.; Cantonnet, M.; Buteau, B.; Boudinot, P.; Benmansour, A. Survey of transcript expression in rainbow trout leukocytes reveals a major contribution of interferon-responsive genes in the early response to a rhabdovirus infection. J. Virol. 2002, 76, 8040–8049. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.J.; Kim, W.-J.; Kim, H.S.; Lee, Y.J.; Kim, C.H.; Nam, B.-H.; Kim, Y.-O.; Kim, D.-G.; Lee, S.-J.; Lim, S.-G.; et al. Molecular characterization of a tandem-repeat galectin-9 (RuGlec9) from Korean rose bitterling (Rhodeus uyekii). Fish Shellfish Immunol. 2012, 32, 939–944. [Google Scholar] [CrossRef]
- Wang, Y.; Ke, F.; Ma, J.; Zhou, S. A tandem-repeat galectin-9 involved in immune response of yellow catfish, Pelteobagrus fulvidraco, against Aeromonas hydrophila. Fish Shellfish Immunol. 2016, 51, 153–160. [Google Scholar] [CrossRef]
- Zhang, D.L.; Lv, C.H.; Yu, D.H.; Wang, Z. Characterization and functional analysis of a tandem-repeat galectin-9 in large yellow croaker Larimichthys crocea. Fish Shellfish Immunol. 2016, 52, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, Z.; Krishnan, R.; Prasad, K.P.; Bedekar, M.K.; Pavan-Kumar, A. Molecular cloning, characterization and expression profiling of galectin-9 gene from Labeo rohita (Hamilton, 1822). Fish Shellfish Immunol. 2018, 76, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Huang, Y.; Li, Y.; Wang, Z.; Tang, J.; Wang, B.; Lu, Y.; Cai, J.; Jian, J. Characterization of a tandem-repeat galectin-9 from Nile tilapia (Oreochromis niloticus) involved in the immune response against bacterial infection. Fish Shellfish Immunol. 2019, 92, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Jung, S.J.; Choi, T.J.; Kim, H.R.; Rajendran, K.V.; Oh, M.J. PCR amplification and sequence analysis of irido-like virus infecting fish in Korea. J. Fish Dis. 2002, 25, 121–124. [Google Scholar] [CrossRef]
- Shin, G.-H.; Shin, Y.; Jung, M.; Hong, J.-M.; Lee, S.; Subramaniyam, S.; Noh, E.-S.; Shin, E.-H.; Park, E.-H.; Park, J.Y.; et al. First draft genome for red sea bream of family Sparidae. Front. Genet. 2018, 9, 643. [Google Scholar] [CrossRef] [Green Version]
- Rabinovich, G.A. Galectins: An evolutionarily conserved family of animal lectins with multifunctional properties; a trip from the gene to clinical therapy. Cell Death Differ. 1999, 6, 711–721. [Google Scholar] [CrossRef]
- Nagae, M.; Nishi, N.; Murata, T.; Usui, T.; Nakamura, T.; Wakatsuki, S.; Kato, R. Crystal structure of the galectin-9 n-terminal carbohydrate recognition domain from mus musculus reveals the basic mechanism of carbohydrate recognition. J. Biol. Chem. 2006, 281, 35884–35893. [Google Scholar] [CrossRef] [Green Version]
- Bi, S.; Earl, L.A.; Jacobs, L.; Baum, L.G. Structural features of galectin-9 and galectin-1 that determine distinct t cell death pathways. J. Biol. Chem. 2008, 283, 12248–12258. [Google Scholar] [CrossRef] [Green Version]
- Nishi, N.; Itoh, A.; Fujiyama, A.; Yoshida, N.; Araya, S.-I.; Hirashima, M.; Shoji, H.; Nakamura, T. Development of highly stable galectins: Truncation of the linker peptide confers protease-resistance on tandem-repeat type galectins. FEBS Lett. 2005, 579, 2058–2064. [Google Scholar] [CrossRef] [Green Version]
- Labrie, M.; Vladoiu, M.; Leclerc, B.G.; Grosset, A.-A.; Gaboury, L.; Stagg, J.; St-Pierre, Y. A mutation in the carbohydrate recognition domain drives a phenotypic switch in the role of galectin-7 in prostate cancer. PLoS ONE 2015, 10, e0131307. [Google Scholar] [CrossRef]
- Hwang, E.Y.; Jeong, M.S.; Park, S.K.; Ha, S.C.; Yu, H.S.; Jang, S.B. Structural basis for carbohydrate recognition and anti-inflammatory modulation by gastrointestinal nematode parasite Toxascaris leonina galectin. J. Biol. Chem. 2016, 291, 25326–25338. [Google Scholar] [CrossRef] [Green Version]
- Wada, J.; Ota, K.; Kumar, A.I.; Wallner, E.; Kanwar, Y.S. Developmental regulation, expression, and apoptotic potential of galectin-9, a beta-galactoside binding lectin. J. Clin. Investig. 1997, 99, 2452–2461. [Google Scholar] [CrossRef] [PubMed]
- Perillo, N.L.; Marcus, M.E.; Baum, L.G. Galectins: Versatile modulators of cell adhesion, cell proliferation, and cell death. J. Mol. Med. 1998, 76, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Spitzenberger, F.; Graessler, J.; Schroeder, H.-E. Molecular and functional characterization of galectin 9 mRNA isoforms in porcine and human cells and tissues. Biochimie 2001, 83, 851–862. [Google Scholar] [CrossRef]
- Matsumoto, R.; Matsumoto, H.; Seki, M.; Hata, M.; Asano, Y.; Kanegasaki, S.; Stevens, R.L.; Hirashima, M. Human ecalectin, a variant of human galectin-9, is a novel eosinophil chemoattractant produced by T lymphocytes. J. Biol. Chem. 1998, 273, 16976–16984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Fueyo, A.; Tian, J.; Picarella, D.; Domenig, C.; Zheng, X.X.; Sabatos, C.A.; Manlongat, N.; Bender, O.; Kamradt, T.; Kuchroo, V.K.; et al. Tim-3 inhibits T helper type 1-mediated auto- and alloimmune responses and promotes immunological tolerance. Nat. Immunol. 2003, 4, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Cano, L.Q.; Tagit, O.; Dolen, Y.; Van Duffelen, A.; Dieltjes, S.; Buschow, S.I.; Niki, T.; Hirashima, M.; Joosten, B.; van den Dries, K.; et al. Intracellular galectin-9 controls dendritic cell function by maintaining plasma membrane rigidity. iScience 2019, 22, 240–255. [Google Scholar] [CrossRef] [Green Version]
- Inagawa, H.; Kuroda, A.; Nishizawa, T.; Honda, T.; Ototake, M.; Yokomizo, Y.; Nakanishi, T.; Soma, G.-I. Cloning and characterisation of tandem-repeat type galectin in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2001, 11, 217–231. [Google Scholar] [CrossRef]
- Wang, F.; He, W.; Zhou, H.; Yuan, J.; Wu, K.; Xu, L.; Chen, Z.K. The Tim-3 ligand galectin-9 negatively regulates CD8+ alloreactive T cell and prolongs survival of skin graft. Cell. Immunol. 2007, 250, 68–74. [Google Scholar] [CrossRef]
- Vega-Carrascal, I.; Bergin, D.A.; McElvaney, O.J.; McCarthy, C.; Banville, N.; Pohl, K.; Hirashima, M.; Kuchroo, V.K.; Reeves, E.P.; McElvaney, N.G. Galectin-9 Signaling through TIM-3 Is involved in neutrophil-mediated gram-negative bacterial killing: An effect abrogated within the cystic fibrosis lung. J. Immunol. 2014, 192, 2418–2431. [Google Scholar] [CrossRef]
- Steichen, A.L.; Simonson, T.J.; Salmon, S.L.; Metzger, D.W.; Mishra, B.B.; Sharma, J. Alarmin Function of Galectin-9 in murine respiratory tularemia. PLoS ONE 2015, 10, e0123573. [Google Scholar] [CrossRef]
- Inouye, K.; Yamano, K.; Maeno, Y.; Nakajima, K.; Matsuoka, M.; Wada, Y.; Sorimachi, M. Iridovirus infection of cultured red sea bream, Pagrus major. Fish Pathol. 1992, 27, 19–27. [Google Scholar] [CrossRef]
- Chou, H.-Y.; Hsu, C.-C.; Peng, T.-Y. Isolation and characterization of a pathogenic iridovirus from cultured grouper (Epinephelus sp.) in Taiwan. Fish Pathol. 1998, 33, 201–206. [Google Scholar] [CrossRef]
- Sudthongkong, C.; Miyata, M.; Miyazaki, T. Viral DNA sequences of genes encoding the ATPase and the major capsid protein of tropical iridovirus isolates which are pathogenic to fishes in Japan, South China Sea and Southeast Asian countries. Arch. Virol. 2002, 147, 2089–2109. [Google Scholar] [CrossRef] [PubMed]
- Girisha, S.K.; Puneeth, T.G.; Nithin, M.S.; Naveen-Kumar, B.T.; Ajay, S.K.; Vinay, T.N.; Suresh, T.; Venugopal, M.N.; Ramesh, K. S Red sea bream iridovirus disease (RSIVD) outbreak in Asian seabass (Lates calcarifer) cultured in open estuarine cages along the west coast of India: First report. Aquaculture 2020, 520, 734712. [Google Scholar] [CrossRef]
- Alcendor, D.J.; Knobel, S.M.; Desai, P.; Zhu, W.Q.; Vigil, H.E.; Hayward, G.S. KSHV downregulation of galectin-3 in Kaposi’s sarcoma. Glycobiology 2009, 20, 521–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo, K.A.; Fermino, M.L.; Andrade, C.D.C.; Riul, T.B.; Alves, R.T.; Muller, V.D.M.; Russo, R.R.; Stowell, S.R.; Cummings, R.D.; Aquino, V.H.; et al. Galectin-1 exerts inhibitory effects during denv-1 infection. PLoS ONE 2014, 9, e112474. [Google Scholar] [CrossRef] [Green Version]
- Uluca, U.; Sen, V.; Ece, A.; Tan, I.; Karabel, D.; Aktar, F.; Karabel, M.; Balık, H.; Güneş, A. Serum galectin-3 levels in children with chronic hepatitis B infection and inactive hepatitis B carriers. Med. Sci. Monit. 2015, 21, 1376–1380. [Google Scholar] [CrossRef] [Green Version]
- Shimmoto, H.; Kawai, K.; Ikawa, T.; Oshima, S.-I. Protection of red sea bream Pagrus major against red sea bream iridovirus infection by vaccination with a recombinant viral protein. Microbiol. Immunol. 2010, 54, 135–142. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Species | Protein Sequence Length | GenBank Accession No. | Protein Similarity (%) |
|---|---|---|---|---|
| Rock bream | Oplegnathus fasciatus | 347 | ANN46244 | 85.6 |
| Gilthead sea bream | Sparus aurata | 361 | XP_030261557 | 85.0 |
| Large yellow croaker | Larimichthys crocea | 343 | XP_010754381 | 79.0 |
| Nile tilapia | Oreochromis niloticus | 324 | XP_003458375 | 72.9 |
| Rainbow trout | Oncorhynchus mykiss | 341 | ACO08221 | 65.0 |
| Atlantic salmon | Salmo salar | 344 | ACI67584 | 64.1 |
| Pig | Sus scrofa | 355 | NP_999097 | 45.4 |
| Cattle | Bos taurus | 355 | NP_001034266 | 44.0 |
| Human | Homo sapiens | 355 | CAB93851 | 43.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, K.-M.; Joo, M.-S.; Cho, D.-H.; Woo, W.-S.; Kang, G.; Heo, M.J.; Kim, D.-H.; Park, C.-I. Molecular Identification and mRNA Expression Profiles of Galectin-9 Gene in Red Sea Bream (Pagrus major) Infected with Pathogens. Animals 2021, 11, 139. https://doi.org/10.3390/ani11010139
Choi K-M, Joo M-S, Cho D-H, Woo W-S, Kang G, Heo MJ, Kim D-H, Park C-I. Molecular Identification and mRNA Expression Profiles of Galectin-9 Gene in Red Sea Bream (Pagrus major) Infected with Pathogens. Animals. 2021; 11(1):139. https://doi.org/10.3390/ani11010139
Chicago/Turabian StyleChoi, Kwang-Min, Min-Soo Joo, Dong-Hee Cho, Won-Sik Woo, Gyoungsik Kang, Min Jin Heo, Do-Hyung Kim, and Chan-Il Park. 2021. "Molecular Identification and mRNA Expression Profiles of Galectin-9 Gene in Red Sea Bream (Pagrus major) Infected with Pathogens" Animals 11, no. 1: 139. https://doi.org/10.3390/ani11010139