Growth Performance, Serum Biochemical Indices, Duodenal Histomorphology, and Cecal Microbiota of Broiler Chickens Fed on Diets Supplemented with Cinnamon Bark Powder at Prestarter and Starter Phases

 ,
,  ,
,  , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Proximate Analysis of Cinnamon Bark Meal
2.2. Phytochemical Analysis of Cinnamon Bark Meal
2.3. Bird Husbandry, Dietary Treatments, and Experimental Design
2.4. Growth Performance Indices
2.5. Samplings
2.6. Serum Biochemical Indices
2.7. Morphometric Analysis of Duodenum
2.8. Cecal Microflora
2.9. Statistical Analysis
3. Results
3.1. Proximate Analysis and Phytochemicals of Cinnamon Bark Meal
3.2. Growth Performance Indices
3.3. Serum Biochemical Indices
3.4. Organ Development
3.5. Morphometric Analysis of Duodenum
3.6. Cecal Microflora
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Leeson, S. Predictions for Commercial Poultry Nutrition. J. Appl. Poult. Res. 2008, 17, 315–322. [Google Scholar] [CrossRef]
- European Medicines Agency. Sales of Veterinary Antimicrobial Agents in 30 European Countries in 2015; European Medicines Agency: Amsterdam, The Netherlands, 2017; p. 178. [Google Scholar]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [PubMed]
- Hughes, P.; Heritage, J. Antibiotic growth-promoters in food animals. FAO Anim. Prod. Health 2004, 160, 129–152. [Google Scholar]
- Han, T.; Zhang, Q.; Liu, N.; Wang, J.; Li, Y.; Huang, X.; Liu, J.; Wang, J.; Qu, Z.; Qi, K. Changes in antibiotic resistance of Escherichia coli during the broiler feeding cycle. Poult. Sci. 2020, 99, 6983–6989. [Google Scholar] [CrossRef]
- Schokker, D.; Jansman, A.; Veninga, G.; De Bruin, N.; Vastenhouw, S.A.; De Bree, F.M.; Bossers, A.; Rebel, J.M.J.; Smits, M.A. Perturbation of microbiota in one-day old broiler chickens with antibiotic for 24 hours negatively affects intestinal immune development. BMC Genom. 2017, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.K.; Kumar, P.; Saxena, M.J. Feed Additives in Animal Health. In Nutraceuticals in Veterinary Medicine; Springe: Berlin, Germany, 2019; pp. 345–362. [Google Scholar]
- Diarra, M.S.; Malouin, F. Antibiotics in Canadian poultry productions and anticipated alternatives. Front. Microbiol. 2014, 5, 282. [Google Scholar] [CrossRef] [Green Version]
- Toghyani, M.; Toghyani, M.; Gheisari, A.; Ghalamkari, G.; Eghbalsaied, S. Evaluation of cinnamon and garlic as antibiotic growth promoter substitutions on performance, immune responses, serum biochemical and haematological parameters in broiler chicks. Livest. Sci. 2011, 138, 167–173. [Google Scholar] [CrossRef]
- Abdel-Wareth, A.A.A.; Kehraus, S.; Südekum, K.-H. Peppermint and its respective active component in diets of broiler chickens: Growth performance, viability, economics, meat physicochemical properties, and carcass characteristics. Poult. Sci. 2019, 98, 3850–3859. [Google Scholar] [CrossRef]
- Christensen, V.L. Development during the First Seven Days Post-hatching. Avian Biol. Res. 2009, 2, 27–33. [Google Scholar] [CrossRef]
- Lilburn, M.S.; Loeffler, S. Early intestinal growth and development in poultry. Poult. Sci. 2015, 94, 1569–1576. [Google Scholar] [CrossRef]
- Mahdavi, R.; Karlovich, O.A.; Ficinin, V.I. Effects of supplying time of prestarter on subsequent broiler performance, carcass yield and intestinal morphometry. Russ. Agric. Sci. 2017, 43, 326–331. [Google Scholar] [CrossRef]
- Popoola, I.O.; Oshibanjo, D.O.; Popoola, O.R.; Okuneye, T.A.; Ilaboya, I.I.; Iyayi, E.A. Effect of Dietary Electrolyte Balance on Water Intake, Litter Moisture and Production of Broiler chicks at Pre-Starter and Starter Phases. Open J. Anim. Sci. 2019, 9, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Knezevic, J.; Starchl, C.; Berisha, A.T.; Amrein, K. Thyroid-Gut-Axis: How Does the Microbiota Influence Thyroid Function? Nutrients 2020, 12, 1769. [Google Scholar] [CrossRef]
- Kogut, M.H. The effect of microbiome modulation on the intestinal health of poultry. Anim. Feed. Sci. Technol. 2019, 250, 32–40. [Google Scholar] [CrossRef]
- Vangalapati, M.; Satya, S.; Prakash, S.; Avanigadda, S. A review on pharmacological activities and clinical effects of cinnamon species. Res. J. Pharm. Biol. Chem. Sci. 2012, 3, 653–663. [Google Scholar]
- Rao, P.V.; Gan, S.H. Cinnamon: A Multifaceted Medicinal Plant. Evid. Based Complement. Altern. Med. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Fauvelle, C.; Lambotin, M.; Heydmann, L.; Prakash, E.; Bhaskaran, S.; Vishwaraman, M.; Moog, C.; Baumert, T.F. A cinnamon-derived procyanidin type A compound inhibits hepatitis C virus cell entry. Hepatol. Int. 2017, 11, 440–445. [Google Scholar] [CrossRef]
- Noshirvani, N.; Ghanbarzadeh, B.; Gardrat, C.; Rezaei, M.R.; Hashemi, M.; Le Coz, C.; Coma, V. Cinnamon and ginger essential oils to improve antifungal, physical and mechanical properties of chitosan-carboxymethyl cellulose films. Food Hydrocoll. 2017, 70, 36–45. [Google Scholar] [CrossRef]
- Sharma, U.K.; Sharma, A.K.; Pandey, A.K. Medicinal attributes of major phenylpropanoids present in cinnamon. BMC Complement. Altern. Med. 2016, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Wang, T.; Chen, L.; Yu, B.-W.; Jia, Q.; Chen, K.-X.; Fan, H.-M.; Li, Y.-M.; Wang, H.-Y. Trimer procyanidin oligomers contribute to the protective effects of cinnamon extracts on pancreatic β-cells in vitro. Acta Pharmacol. Sin. 2016, 37, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Benoit, J.; Oerting, H.; Schroeder, K.; Goemann, C.; Diaz, E. Fragrance Composition. U.S. Patent 20,160,032,217, 4 February 2016. [Google Scholar]
- Kumar, M.; Kumar, V.; Roy, D.; Kushwaha, R.; Vaswani, S. Application of herbal feed additives in animal nutrition: A Review. Int. J. Livest. Res. 2014, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Faghani, M.; Rahimian, Y.; Rafiee, A.; Namjoo, A.R. Effect of garlic and cinnamon in comparison to virginiamycin on performance and some haematological parameters in broiler chicks. Res. Opin. Anim. Vet. Sci. 2014, 4, 504–507. [Google Scholar]
- Sang-Oh, P.; Chae-Min, R.; Byung-Sung, P.; Jong, H. The meat quality and growth performance in broiler chickens fed diet with cinnamon powder. J. Environ. Biol. 2013, 34, 127. [Google Scholar] [PubMed]
- Hussein, T.K.; Hwaidi, E.H.; Mohammad, A.H. The effects of cinnamon powder feeding on some blood aspects and performance of broiler chicken. Kufa J. Vet. Sci. 2016, 6, 118–122. [Google Scholar]
- AOAC-International. Official Methods of Analysis; AOAC International: Gaithersburg, MD, USA, 2006. [Google Scholar]
- Oryschak, M.; Korver, D.; Zuidhof, M.; Beltranena, E. Nutritive value of single-screw extruded and nonextruded triticale distillers dried grains with solubles, with and without an enzyme complex, for broilers. Poult. Sci. 2010, 89, 1411–1423. [Google Scholar] [CrossRef] [PubMed]
- Holst, D.O. Holst Filtration Apparatus for Van Soest Detergent Fiber Analyses. J. Assoc. Off. Anal. Chem. 1973, 56, 1352–1356. [Google Scholar] [CrossRef]
- Humeera, N.; Kamili, A.N.; Bandh, S.A.; Amin, S.-U.; Lone, B.A.; Gousia, N. Antimicrobial and antioxidant activities of alcoholic extracts of Rumex dentatus L. Microb. Pathog. 2013, 57, 17–20. [Google Scholar] [CrossRef]
- Almazroui, M.; Islam, M.N.; Athar, H.; Jones, P.D.; Rahman, M.A. Recent climate change in the Arabian Peninsula: Annual rainfall and temperature analysis of Saudi Arabia for 1978–2009. Int. J. Clim. 2012, 32, 953–966. [Google Scholar] [CrossRef]
- Alhotan, R.A.; Vedenov, D.V.; Pesti, G.M. Estimation of the maximum safe level of feed ingredients by spline or broken-line nonlinear regression models. Poult. Sci. 2017, 96, 904–913. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Nutrient Requirements of Poultry; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Rostagno, H.; Albino, L.; Donzele, J.; Gomes, P.; Oliveira, R.; Lopes, D.; Ferreira, A.; Barreto, S.T.; Euclides, R. Brazilian Tables for Poultry and Swine: Composition of Feedstuffs and Nutritional Requirements, 3rd ed.; UFV: Viçosa, Brazil, 2011. [Google Scholar]
- Aptekmann, K.P.; Artoni, S.; Stefanini, M.A.; Orsi, M.A. Morphometric Analysis of the Intestine of Domestic Quails (Coturnix coturnix japonica) Treated with Different Levels of Dietary Calcium. Anat. Histol. Embryol. 2001, 30, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Hirose, H.; Onizuka, A.; Hayashi, M.; Futamura, N.; Kawamura, Y.; Ezaki, T. Quantitative Study of Changes in Intestinal Morphology and Mucus Gel on Total Parenteral Nutrition in Rats. J. Surg. Res. 2000, 94, 99–106. [Google Scholar] [CrossRef]
- SAS. SAS Institute/OR 9.3 User’s Guide: Mathematical Programming Examples; SAS Institute: Cary, NC, USA, 2012. [Google Scholar]
- Elbadawy, M.; Aboubakr, M. Efficacy of colimoxŪ (a new combination of amoxicillin and colistin) in the control of experimentally induced necrotic enteritis in broiler chickens. Int. J. Pharmacol. Toxicol. 2017, 5, 51–56. [Google Scholar] [CrossRef]
- Bedford, M. Removal of antibiotic growth promoters from poultry diets: Implications and strategies to minimise subsequent problems. Worlds Poult. Sci. J. 2000, 56, 347–365. [Google Scholar] [CrossRef]
- Moreira, G.C.; Dias, F.D.S. Mixture design and Doehlert matrix for optimization of the ultrasonic assisted extraction of caffeic acid, rutin, catechin and trans-cinnamic acid in Physalis angulata L. and determination by HPLC DAD. Microchem. J. 2018, 141, 247–252. [Google Scholar] [CrossRef]
- Shan, Y.; Jin, X.; Cheng, Y.; Yan, W. Simultaneous determination of chlorogenic acids in green coffee bean extracts with effective relative response factors. Int. J. Food Prop. 2017, 20, 2028–2040. [Google Scholar] [CrossRef]
- Praseptiangga, D.; Invicta, S.E.; Khasanah, L.U. Sensory and physicochemical characteristics of dark chocolate bar with addition of cinnamon (Cinnamomum burmannii) bark oleoresin microcapsule. J. Food Sci. Technol. 2019, 56, 4323–4332. [Google Scholar] [CrossRef] [PubMed]
- Hameed, I.; Al-Tameme, H.J.; Mohammed, G. Evaluation of Antifungal and Antibacterial Activity and Analysis of Bioactive Phytochemical Compounds of Cinnamomum Zeylanicum (Cinnamon Bark) using Gas Chromatography-Mass Spectrometry. Orient. J. Chem. 2016, 32, 1769–1788. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Safdar, M.; Khan, M.M.A.; Khattak, K.N.; Anderson, R.A. Cinnamon improves glucose and lipids of people with type 2 diabetes. Diabetes Care 2003, 26, 3215–3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhavaniramya, S.; Vishnupriya, S.; Al-Aboody, M.S.; Vijayakumar, R.; Baskaran, D. Role of essential oils in food safety: Antimicrobial and antioxidant applications. Grain Oil Sci. Technol. 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Favaretto, J.A.; Alba, D.F.; Marchiori, M.S.; Marcon, H.J.; Souza, C.F.; Baldissera, M.D.; Bianchi, A.E.; Zanluchi, M.; Klein, B.; Wagner, R.; et al. Supplementation with a blend based on micro-encapsulated carvacrol, thymol, and cinnamaldehyde in lambs feed inhibits immune cells and improves growth performance. Livest. Sci. 2020, 240, 104144. [Google Scholar] [CrossRef]
- Al-Kassie, G.A. Influence of two plant extracts derived from thyme and cinnamon on broiler performance. Pak. Vet. J. 2009, 29, 169–173. [Google Scholar]
- Khafaji, S. Study the Effect of Ceylon Cinnamon (Cinnamomumzeylanicum) Powder on Some Physiological Parameters in Broiler Chicks. J. Global Pharma. Technol. 2018, 10, 236–242. [Google Scholar]
- Lee, K.-W.; Everts, H.; Kappert, H.; Frehner, M.; Losa, R.; Beynen, A. Effects of dietary essential oil components on growth performance, digestive enzymes and lipid metabolism in female broiler chickens. Br. Poult. Sci. 2003, 44, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Amirali, S.; Karimi, A.; Jahromi, S.P.; Azizi, T.; Daneshmand, A. Effects of cinnamon, thyme and turmeric infusions on the performance and immune response in of 1- to 21-day-old male broilers. Braz. J. Poult. Sci. 2012, 14, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Oso, A.; Suganthi, R.U.; Reddy, G.M.; Malik, P.; Thirumalaisamy, G.; Awachat, V.; Selvaraju, S.; Arangasamy, A.; Bhatta, R. Effect of dietary supplementation with phytogenic blend on growth performance, apparent ileal digestibility of nutrients, intestinal morphology, and cecal microflora of broiler chickens. Poult. Sci. 2019, 98, 4755–4766. [Google Scholar] [CrossRef]
- Ahmadi, E.; Shahri, M.M. The Antioxidant and Anticoagulant Effects of Coumarin and Quercetin from Cinnamon Methanolic Extract, and the Assessment of Cinnamon Powder Effect on Plasma Parameters in Diabetes, and the Disinfectant Activity in Diabetic Patients. J. Herb. Med. 2020, 4, 103–110. [Google Scholar]
- Khezeli, T.; Daneshfar, A.; Sahraei, R. A green ultrasonic-assisted liquid–liquid microextraction based on deep eutectic solvent for the HPLC-UV determination of ferulic, caffeic and cinnamic acid from olive, almond, sesame and cinnamon oil. Talanta 2016, 150, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Geyra, A.; Uni, Z.; Sklan, D. The effect of fasting at different ages on growth and tissue dynamics in the small intestine of the young chick. Br. J. Nutr. 2001, 86, 53–61. [Google Scholar] [CrossRef]
- Li, J.; Sun, X.; Wan, J. Effects of copper-bearing montmorillonite on growth performance and nutrient digestibility of broilers. China Feed. 2016, 2016, 7. [Google Scholar]
- Chowlu, H.; Vidyarthi, V.; Zuyie, R.; Maiti, C. Effect of Dietary Supplementation of Cinnamon on the Performance of Broiler Chicken. Livest. Res. Int. 2019, 7, 83–87. [Google Scholar]
- Thornton, K.J. Triennial growth symposium: The nutrition of muscle growth: Impacts of nutrition on the proliferation and differentiation of satellite cells in livestock species. J. Anim. Sci. 2019, 97, 2258–2269. [Google Scholar] [CrossRef] [PubMed]
- Tejeda, O.; Calderon, A.; Arana, J.; Meloche, K.; Starkey, J. Broiler chicken myofiber morphometrics and myogenic stem cell population heterogeneity. Poult. Sci. 2019, 98, 4123–4130. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wu, C.; Li, K.; Gui, G.; Zhang, G.; Yang, H. Association of growth rate with hormone levels and myogenic gene expression profile in broilers. J. Anim. Sci. Biotechnol. 2017, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, G.K.; Coelho, J.R.; Jensen, D.R. Body temperature regulation and oxygen consumption in young chicks fed thyroid hormone. Can. J. Zool. 1991, 69, 1842–1847. [Google Scholar] [CrossRef]
- Sun, Q.; Yang, H.; Tang, P.; Liu, J.; Wang, W.; Li, H. Interactions of cinnamaldehyde and its metabolite cinnamic acid with human serum albumin and interference of other food additives. Food Chem. 2018, 243, 74–81. [Google Scholar] [CrossRef]
- Mimoto, M.S.; Refetoff, S. Clinical recognition and evaluation of patients with inherited serum thyroid hormone-binding protein mutations. J. Endocrinol. Investig. 2020, 43, 31–41. [Google Scholar] [CrossRef]
- Mahendhar, R.; Shahbaz, A.; Riaz, M.; Aninyei, M.; Reich, D.M.; Sachmechi, I. Effect of Albumin Polymorphism on Thyroid Hormones: A Case Report and Literature Review. Cureus 2018, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Cuevas-Ramos, G.; Petit, C.R.; Marcq, I.; Boury, M.; Oswald, E.; Nougayrède, J.-P. Escherichia coli induces DNA damage in vivo and triggers genomic instability in mammalian cells. Proc. Natl. Acad. Sci. USA 2010, 107, 11537–11542. [Google Scholar] [CrossRef] [Green Version]
- Brancaccio, P.; Lippi, G.; Maffulli, N. Biochemical markers of muscular damage. Clin. Chem. Lab. Med. 2010, 48, 757–767. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, T.; Cui, X.; Li, S.; Zhao, X.; Zhong, X. Hepatoprotective Effects of a Chinese Herbal Formula, Longyin Decoction, on Carbon-Tetrachloride-Induced Liver Injury in Chickens. Evid. Based Complement. Altern. Med. 2013, 2013, 1–9. [Google Scholar] [CrossRef]
- Choct, M. Managing gut health through nutrition. Br. Poult. Sci. 2009, 50, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, K.; Cloft, S.; Wong, E.A. Changes with age in density of goblet cells in the small intestine of broiler chicks. Poult. Sci. 2020, 99, 2342–2348. [Google Scholar] [CrossRef] [PubMed]
- Hutsko, S.L.; Meizlisch, K.; Wick, M.; Lilburn, M.S. Early intestinal development and mucin transcription in the young poult with probiotic and mannan oligosaccharide prebiotic supplementation. Poult. Sci. 2016, 95, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Windisch, W.; Rohrer, E.; Schedle, K. Phytogenic Feed Additives to Young Piglets and Poultry: Mechanisms and Application; Nottingham University Press: Nottingham, UK, 2009. [Google Scholar]
- Chowdhury, S.; Mandal, G.P.; Patra, A.K.; Kumar, P.; Samanta, I.; Pradhan, S.; Samanta, A.K. Different essential oils in diets of broiler chickens: 2. Gut microbes and morphology, immune response, and some blood profile and antioxidant enzymes. Anim. Feed. Sci. Technol. 2018, 236, 39–47. [Google Scholar] [CrossRef]
- Roura, E.; Homedes, J.; Klasing, K.C. Prevention of Immunologic Stress Contributes to the Growth-Permitting Ability of Dietary Antibiotics in Chicks. J. Nutr. 1992, 122, 2383–2390. [Google Scholar] [CrossRef]
- Gaskins, H.R. Host and intestinal microbiota negotiations in the context of animal growth efficiency. In Gut Efficiency; The Key Ingredient in Pig and Poultry Production; Taylor-Pickard, J.A., Spring, P., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2008; pp. 29–37. [Google Scholar]
- Hedde, R.; Lindsey, T. Virginiamycin: A nutritional tool for swine production. Agri-Practice 1986, 7, 70–72. [Google Scholar]
- Awawdeh, L. Studies on Avian Pathogenic Escherichia coli in Commercial Broiler Chicken in South East Queensland. Ph.D. Thesis, Queensland University, Brisbane, Australia, 2018; pp. 1–276. [Google Scholar]
- Fröhlich, E.; Wahl, R. Microbiota and Thyroid Interaction in Health and Disease. Trends Endocrinol. Metab. 2019, 30, 479–490. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Ingredients, % | 1–21 days |
|---|---|
| Yellow corn, 7.8% | 53.22 |
| Soybean meal, 45% | 37.85 |
| Wheat bran | 2.00 |
| Corn gluten meal | 1.40 |
| Rice bran oil | 1.50 |
| Di-calcium phosphate | 1.98 |
| Limestone | 0.90 |
| Sodium chloride (NaCl) | 0.40 |
| DL-methionine | 0.29 |
| l-lysine HCL | 0.21 |
| Choline chloride | 0.05 |
| Vitamin- mineral premix 1 | 0.20 |
| Cinnamon bark 2 | 0.00 |
| Total | 100 |
| Nutrient content (%, based on as-fed basis) 3 | |
| Crude protein | 22.80 |
| Digestible lysine | 1.26 |
| Digestible sulfur amino acids | 0.91 |
| Digestible threonine | 0.77 |
| Calcium | 0.93 |
| Non-phytate P | 0.45 |
| Metabolizable energy, kcal/kg | 2900 |
| Item | % (on a Dry Matter Basis) |
|---|---|
| Moisture (% as fed) | 10.29 |
| Dry matter (% as fed) | 89.71 |
| Crude protein | 4.43 |
| Ether extract (lipid) | 4.03 |
| Ash | 3.18 |
| Total crude fiber | 24.35 |
| 2 Nitrogen-free extract (NFE) | 53.72 |
| 3 Organic matter | 96.82 |
| Total fiber fractions: | |
| A. Acid detergent fiber | 45.75 |
| B. Neutral detergent fiber | 65.34 |
| Gross Energy (kcal/kg) | 4974.52 |
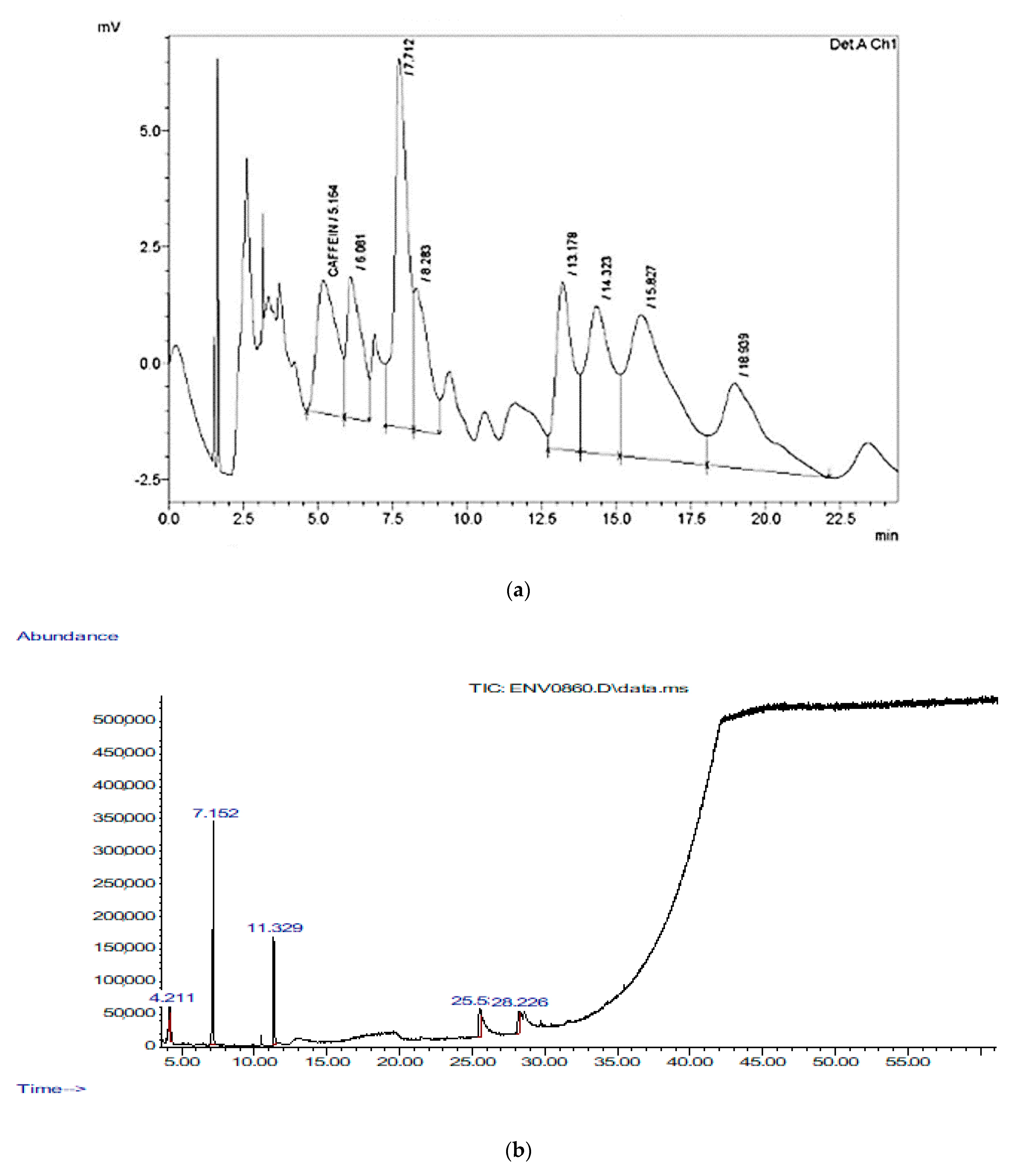
| RT(min) | Bioactive Chemical Constituents | Quality | MW (amu) | Molecular Formula |
|---|---|---|---|---|
| 4.108 | 4-Ethylbenzoic acid, 2-butyl ester | 78 | 206.131 | C6H18O3Si3 |
| 4.108 | 2-Amino-6-methylbenzoic acid; 2-Amino-4-methylbenzoic acid | 59 | 151.063 | C8H9NO2 |
| 4.211 | Oxime-, methoxy-phenyl-_ | 87 | 151.063 | C8H9NO2 |
| 4.211 | 4-Ethylbenzoic acid, hexyl ester | 50 | 234.162 | C15H22O2 |
| 7.152 | Hexamethyl cyclotrisiloxane | 56 | 222.056 | C6H18O3Si3 |
| 7.152 | Silane, 1,4-phenylenebis[trimethyl- | 50 | 222.126 | C14H26Si2 |
| 11.329 | Cinnamaldehyde, (E)- 2-Propenal, 3-phenyl- | 97 | 132.058 | C9H7ClO |
| 11.329 | Cinnamaldehyde, (E)- 3-Phenyl-2-propyn-1-ol | 91 | 132.058 | C9H8O |
| 11.329 | 2-methyl-Benzofuran | 87 | 132.058 | C9H8O |
| 11.329 | Benzene, (2-propynyloxy)- | 72 | 132.058 | C9H8O |
| 11.329 | Benzenemethanol,.alpha.-ethynyl- | 64 | 132.058 | C9H8O |
| 11.329 | Benzopyrimidine, 3,4-dihydro- or 3,4-Dihydroquinazoline | 59 | 132.069 | C8H8N2 |
| 11.329 | 2-Propenal, 3-phenyl- | 58 | 132.058 | C9H8O |
| 11.329 | 2-Propenoyl chloride, 3-phenyl-, (E)- (Cinnamoyl chloride) | 53 | 166.019 | C9H7ClO |
| 11.329 | 4-Methylcoumarine-7-cinnamate | 53 | 306.089 | C19H14O4 |
| 11.329 | 2-Nitrophenyl cinnamamide | 50 | 268.085 | C16H14N2O4 |
| 11.329 | 3-Butenoic acid, 2-oxo-4-phenyl- | 50 | 176.047 | C10H8O3 |
| 25.536 | Hexadecanoic acid, methyl ester | 99 | 270.256 | C17H34O2 |
| 25.536 | Pentadecanoic acid, 14-methyl-, methyl ester | 98 | 270.256 | C17H34O2 |
| 25.536 | Tridecanoic acid, methyl ester | 83 | 228.209 | C14H28O2 |
| 25.536 | Hexadecanoic acid, 2-methyl- | 76 | 270.256 | C17H34O2 |
| 25.536 | Nonadecanoic acid, methyl ester | 70 | 312.303 | C20H40O2 |
| 25.536 | Octadecanoic acid, methyl ester | 62 | 298.287 | C19H38O2 |
| 25.536 | Methyl tetradecanoate | 58 | 242.225 | C15H30O2 |
| 25.536 | Pentadecanoic acid, methyl ester | 58 | 256.24 | C16H32O2 |
| 25.536 | Hexadecanoic acid, 15-methyl-, methyl ester; Heptadecanoic acid, methyl ester | 58 | 284.272 | C18H36O2 |
| Age (Day) | |||||
|---|---|---|---|---|---|
| Treatment 1 | 1 | 7 | 10 | 14 | 21 |
| NC | 45.70 2 | 141.15 | 224.00 b | 375.36 b | 780.47 b |
| PC | 45.67 | 150.76 | 246.57 a | 410.87 a | 844.13 a |
| 2000 CN | 45.80 | 147.00 | 238.20 ab | 410.60 a | 850.67 a |
| 4000 CN | 45.63 | 139.26 | 225.48 b | 374.36 b | 777.53 b |
| 6000 CN | 45.67 | 141.83 | 226.48 b | 387.16 ab | 791.51 b |
| Mean CN | 45.70 | 142.70 | 230.05 | 390.71 | 806.57 |
| SEM 3 | 0.029 | 1.471 | 2.709 | 5.020 | 10.604 |
| p-value | 0.3625 | 0.1022 | 0.0399 | 0.0036 | 0.0053 |
| Parameters | Treatment 1 | ||||||
|---|---|---|---|---|---|---|---|
| Period (days) | NC | PC | 2000CN | 4000 CN | 6000 CN | SEM 3 | p-Value |
| BWG (g) | |||||||
| 1–7 | 13.63 2ab | 15.01 a | 14.46 ab | 13.38 b | 13.74 ab | 0.21 | 0.104 |
| 1–10 | 20.63 b | 23.48 a | 22.43 ab | 21.06 b | 20.98 b | 0.328 | 0.037 |
| 1–14 | 26.36 2b | 29.34 a | 29.32 a | 26.44 b | 27.37 ab | 0.415 | 0.004 |
| 1–21 | 34.24 b | 37.48 a | 37.71 a | 34.23 b | 34.97 b | 0.501 | 0.003 |
| AFI (g) | |||||||
| 1–7 | 20.58 | 21.4 | 19.7 | 20.4 | 20.08 | 0.254 | 0.196 |
| 1–10 | 20.3 | 21.13 | 19.49 | 21.2 | 19.48 | 0.365 | 0.384 |
| 1–14 | 23.84 | 24.75 | 24.28 | 24.41 | 22.95 | 0.389 | 0.539 |
| 1–21 | 36.04 | 38.32 | 36.53 | 36.9 | 35.82 | 0.474 | 0.3 |
| FCR (g/g) | |||||||
| 1–7 | 1.51 | 1.43 | 1.37 | 1.53 | 1.49 | 0.026 | 0.245 |
| 1–10 | 0.99 | 0.9 | 0.87 | 1.02 | 0.94 | 0.023 | 0.246 |
| 1–14 | 0.91 | 0.85 | 0.83 | 0.93 | 0.85 | 0.015 | 0.131 |
| 1–21 | 1.05 a | 1.03 ab | 0.97 b | 1.08 a | 1.03 ab | 0.012 | 0.027 |
| FE (g/g) | |||||||
| 1–7 | 0.66 | 0.7 | 0.74 | 0.66 | 0.69 | 0.011 | 0.18 |
| 1–10 | 1.03 | 1.12 | 1.16 | 1.02 | 1.08 | 0.023 | 0.298 |
| 1–14 | 1.11 | 1.19 | 1.23 | 1.09 | 1.19 | 0.021 | 0.134 |
| 1–21 | 0.95 b | 0.98 ab | 1.04 a | 0.93 b | 0.98 ab | 0.011 | 0.026 |
| Treatment 1 | Uric Acid | T3 | T4 | AST | ALT |
|---|---|---|---|---|---|
| NC | 2.71 2 | 3.79 | 4.92 | 197.44 | 11.35 |
| PC | 3.37 | 3.92 | 3.49 | 203.49 | 9.81 |
| 2000 CN | 2.74 | 4.09 | 3.96 | 198.49 | 12.95 |
| 4000 CN | 3.10 | 4.07 | 4.07 | 213.57 | 10.10 |
| 6000 CN | 2.90 | 3.92 | 4.86 | 235.59 | 9.96 |
| SEM 3 | 0.1707 | 0.1247 | 0.2287 | 12.273 | 0.463 |
| p-value | 0.7408 | 0.9472 | 0.6012 | 0.8042 | 0.1538 |
| Groups 1 | Cinnamon (mg/kg) | ||||||
|---|---|---|---|---|---|---|---|
| Parameters | NC | PC | 2000 | 4000 | 6000 | SEM 2 | p-Value |
| Pancreas | 0.451 3 | 0.43 | 0.4 | 0.41 | 0.44 | 0.014 | 0.838 |
| Heart | 0.68 | 0.65 | 0.66 | 0.66 | 0.72 | 0.015 | 0.58 |
| Liver | 2.85 | 2.76 | 2.87 | 3.04 | 2.88 | 0.052 | 0.583 |
| Proventriculus | 0.86 | 0.81 | 0.89 | 0.88 | 0.89 | 0.016 | 0.47 |
| Gizzard | 3.13 | 2.87 | 2.96 | 3.03 | 2.91 | 0.041 | 0.254 |
| Bursa | 0.21 | 0.26 | 0.23 | 0.2 | 0.26 | 0.0102 | 0.287 |
| Spleen | 0.06 | 0.08 | 0.06 | 0.06 | 0.08 | 0.005 | 0.401 |
| Thymus | 0.41 | 0.51 | 0.45 | 0.53 | 0.56 | 0.024 | 0.192 |
| Breast | 15.65 b | 18.37 a | 19.05 a | 18.08 a | 18.67 a | 0.329 | 0.011 |
| Legs | 16.23 | 16.37 | 16.89 | 15.94 | 16.13 | 0.21 | 0.745 |
| Groups | Cinnamon (mg/kg) | ||||||
|---|---|---|---|---|---|---|---|
| Treatment 1 | NC | PC | 2000 | 4000 | 6000 | SEM 3 | p-Value |
| VL (μm) | 552.47 2bc | 537.34 c | 648.14 a | 610.27 ab | 585.10 abc | 11.4625 | 0.0098 |
| VW (μm) | 110.93 a | 85.70 b | 81.35 b | 91.77 b | 87.02 b | 2.2766 | <0.0001 |
| VSA (mm2) | 0.19 a | 0.15 b | 0.17 ab | 0.17 ab | 0.16 b | 0.0046 | 0.0092 |
| CD (μm) | 58.52 b | 67.38 ab | 57.60 b | 69.52 ab | 72.28 a | 1.911 | 0.0393 |
| VL/CD | 9.25 b | 8.17 b | 13.48 a | 7.54 b | 8.14 b | 0.6445 | 0.0239 |
| GCD | 64.80 d | 66.27 d | 102.44 a | 79.00 c | 91.42 b | 2.4996 | <0.0001 |
| GCD/100 µm VA | 5.81 b | 6.18 b | 8.03 a | 6.53 b | 7.88 a | 0.1986 | <0.0001 |
| Treatment 1 | Lactobacillus | Aerobic | Escherichia coli | Salmonella | Lactobacillus/Escherichia coli |
|---|---|---|---|---|---|
| NC | 6.65 2 | 8.795 | 7.32 c | 3.39 | 0.92 a |
| PC | 5.34 | 9.451 | 7.82 bc | 3.48 | 0.73 ab |
| 2000 CN | 6.01 | 9.810 | 11.41 a | 3.82 | 0.53 b |
| 4000 CN | 6.17 | 9.404 | 10.70 a | 3.84 | 0.58 b |
| 6000 CN | 5.89 | 9.154 | 9.90 ab | 4.33 | 0.60 b |
| SEM 3 | 0.156 | 0.446 | 0.514 | 0.563 | 0.047 |
| p-value | 0.085 | 0.978 | 0.012 | 0.182 | 0.022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qaid, M.M.; Al-Mufarrej, S.I.; Azzam, M.M.; Al-Garadi, M.A.; Albaadani, H.H.; Alhidary, I.A.; Aljumaah, R.S. Growth Performance, Serum Biochemical Indices, Duodenal Histomorphology, and Cecal Microbiota of Broiler Chickens Fed on Diets Supplemented with Cinnamon Bark Powder at Prestarter and Starter Phases. Animals 2021, 11, 94. https://doi.org/10.3390/ani11010094
Qaid MM, Al-Mufarrej SI, Azzam MM, Al-Garadi MA, Albaadani HH, Alhidary IA, Aljumaah RS. Growth Performance, Serum Biochemical Indices, Duodenal Histomorphology, and Cecal Microbiota of Broiler Chickens Fed on Diets Supplemented with Cinnamon Bark Powder at Prestarter and Starter Phases. Animals. 2021; 11(1):94. https://doi.org/10.3390/ani11010094
Chicago/Turabian StyleQaid, Mohammed M., Saud I. Al-Mufarrej, Mahmoud M. Azzam, Maged A. Al-Garadi, Hani H. Albaadani, Ibrahim A. Alhidary, and Riyadh S. Aljumaah. 2021. "Growth Performance, Serum Biochemical Indices, Duodenal Histomorphology, and Cecal Microbiota of Broiler Chickens Fed on Diets Supplemented with Cinnamon Bark Powder at Prestarter and Starter Phases" Animals 11, no. 1: 94. https://doi.org/10.3390/ani11010094