Morphological and Molecular Characterization of Kochinema farodai Baqri and Bohra, 2001 (Dorylaimida: Nordiidae) from California, with the First Molecular Study and an Updated Taxonomy of the Genus


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling, Extraction, and Morphological Identification
2.2. DNA Extraction, PCR, and Sequencing
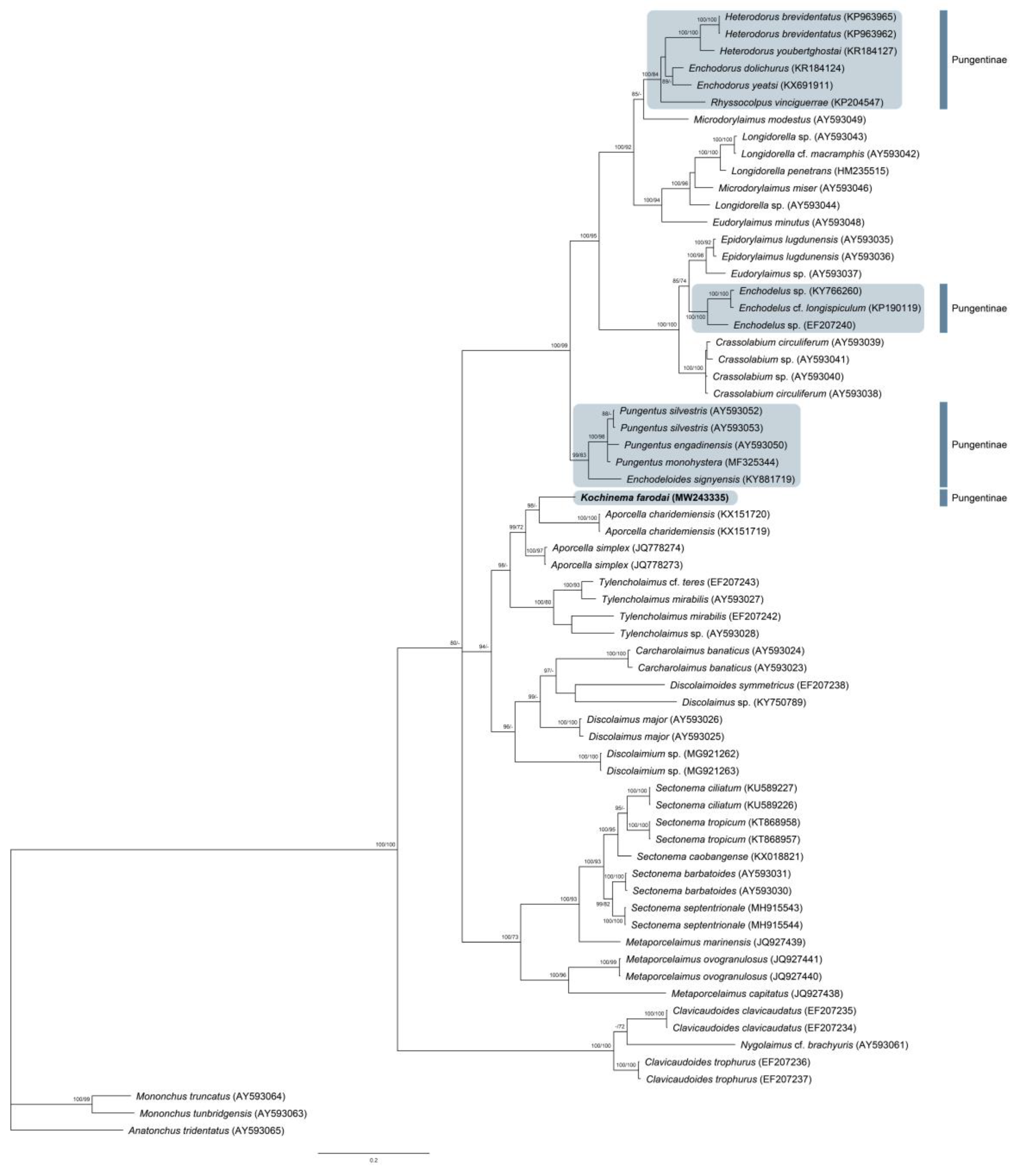
2.3. Phylogenetic Analyses
3. Results and Discussion
3.1. Systematics
3.1.1. Material Examined
3.1.2. Morphometrics
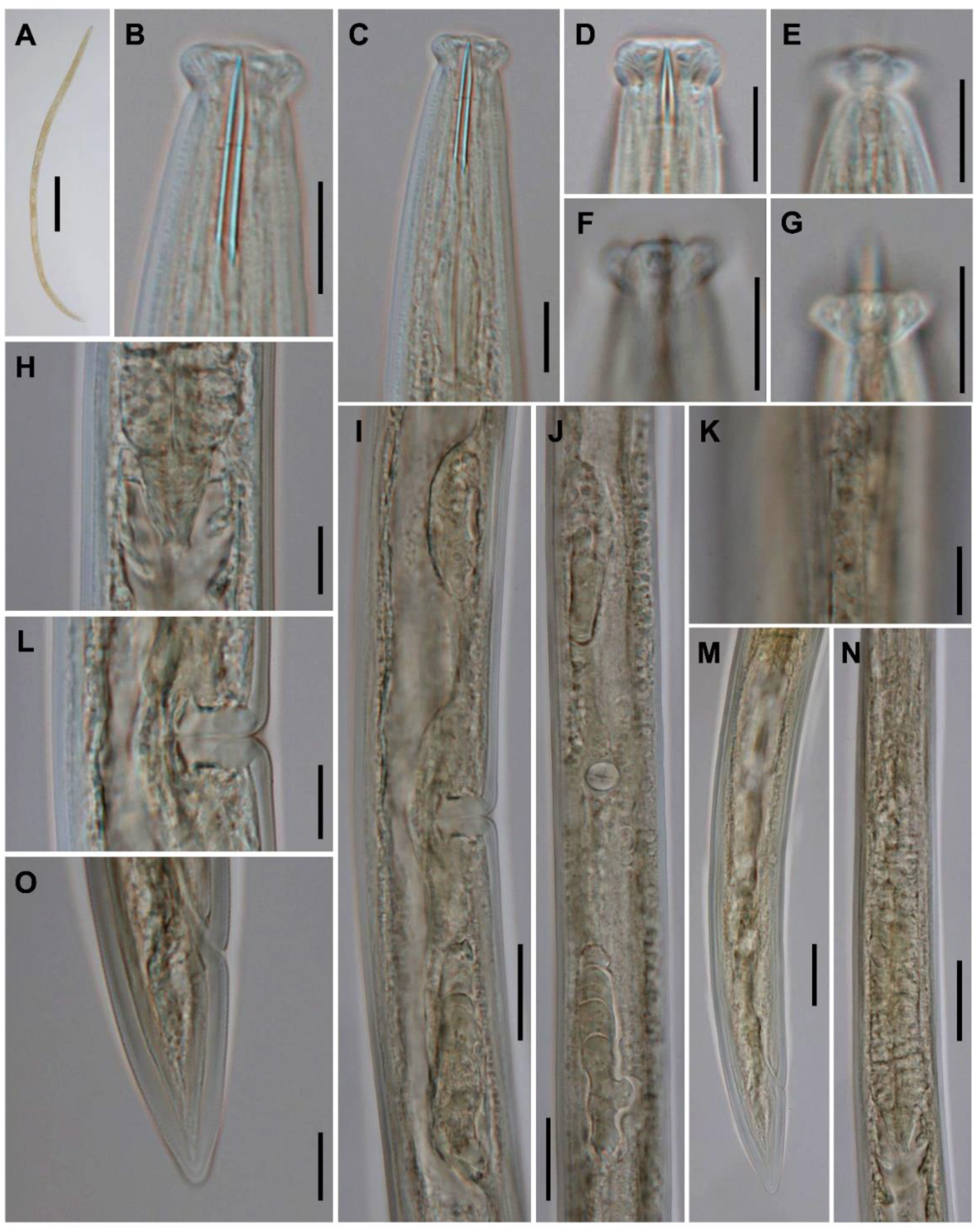
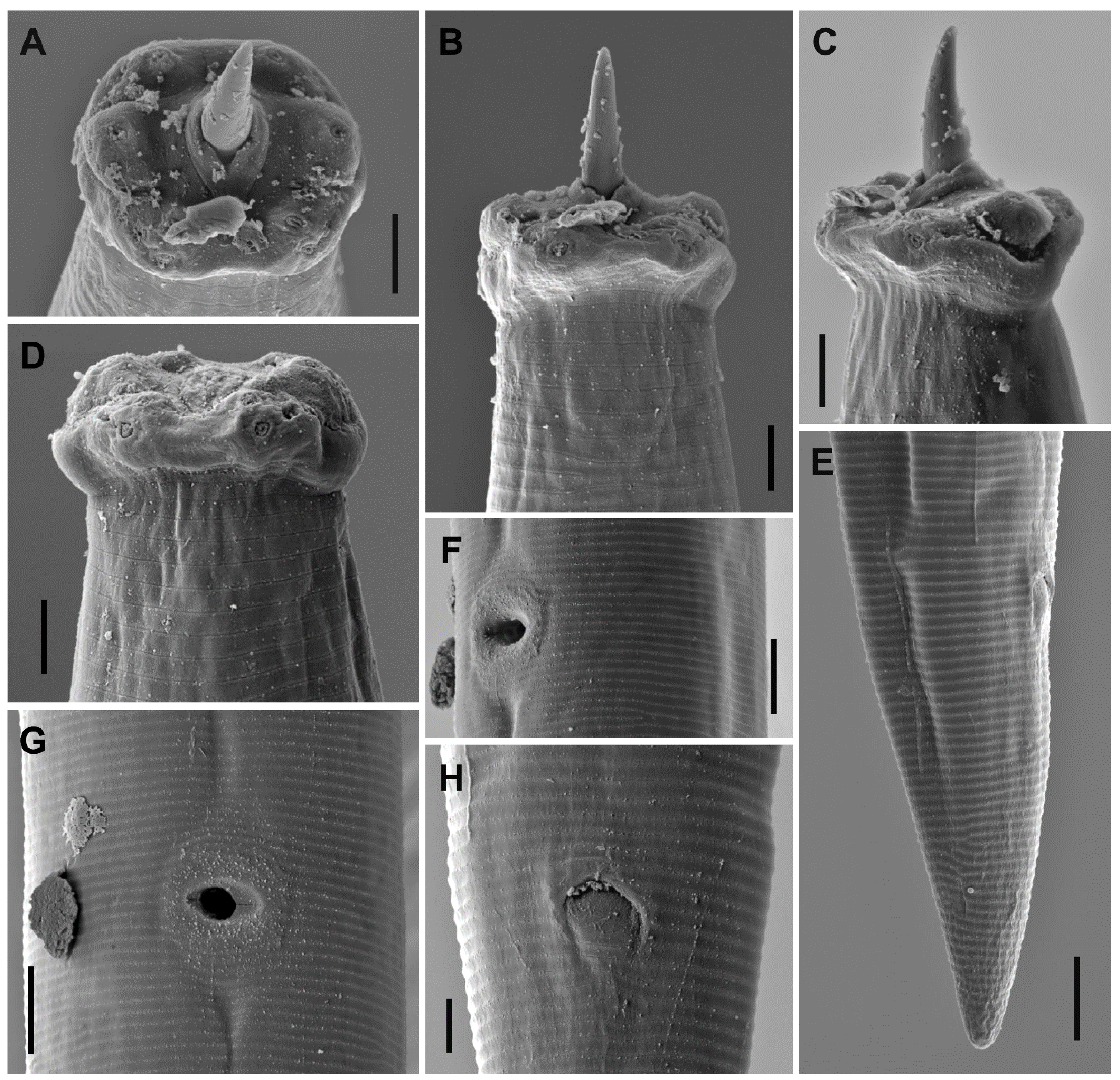
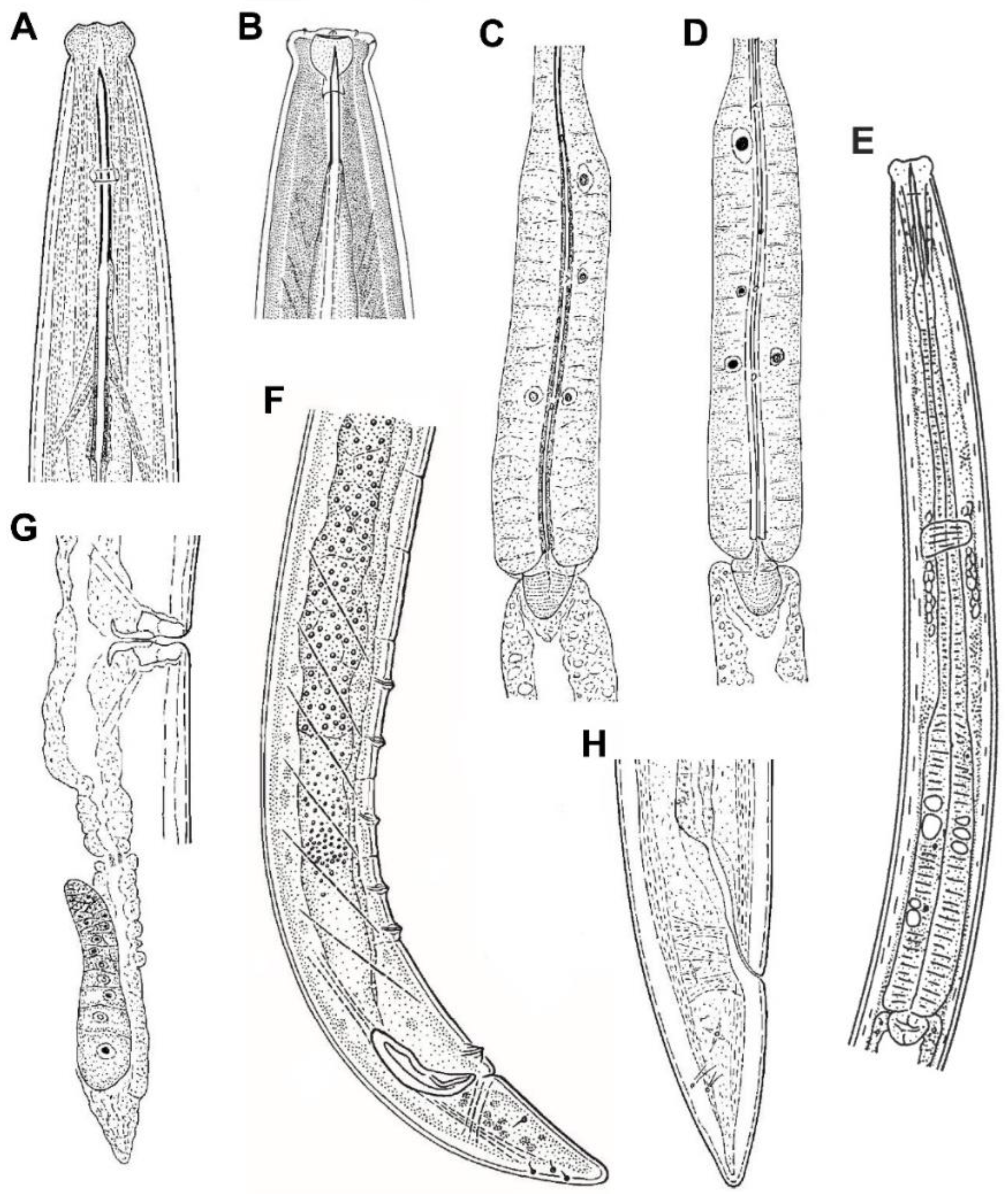
3.1.3. Description
3.1.4. Molecular Characterization
3.1.5. Diagnosis (Based on Specimens Examined)
3.1.6. Remarks
3.1.7. Distribution
3.2. On the Identity and the Taxonomy of Kochinema
3.2.1. Historical Outline
3.2.2. General Morphology
3.3. On the Phylogeny of Kochinema
3.4. Updated Taxonomy of Kochinema
3.4.1. Diagnosis
3.4.2. List of Species
3.4.3. Key to Species Identification
| 1 | - Longer odontostyle, more than 30 μm | 2 |
| - Shorter odontostyle, equal or less than 22 μm | 4 | |
| 2 | - Longer odontostyle, 41 μm long; and comparatively shorter female tail, c′ = 1.0 | crassatum |
| - Shorter odontostyle, up to 38 μm; and comparatively longer female tail, c′ ≥ 1.6 | 3 | |
| 3 | - Odontostyle 35–38 μm long or 2.6–2.7 times the lip region diameter; comparatively longer neck in females, b = 3.1–4.0; comparatively shorter female tail, c′ = 1.6–1.7; and male absent | caudatum |
| - Odontostyle 31–35 μm long or 2.0 times the lip region diameter; comparatively shorter neck in females, b = 4.0–4.9; comparatively longer female tail, c′ = 2.0; and male present | proamphidum | |
| 4 | - Body 1.36–1.60 mm long and very slender, a = 48–59; vulva more posterior, V = 59 | longum |
| - Body up to 1.30 mm long and less slender, a up to 45; vulva less posterior, V = 51–56 | 5 | |
| 5 | - Shorter odontostyle, 13–15 μm long; and male present | secutum |
| - Longer odontostyle, equal or more than 15 μm long; and male absent | 6 | |
| 6 | - Lip region 9–10 μm broad; odontostyle aperture occupying 20% of total odontostyle length; and double guiding ring | tenue |
| - Lip region 10–11 μm broad; and odontostyle aperture occupying 25–35% of total odontostyle length; and simple guiding ring | farodai |
3.4.4. Notes about Some Species
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Andrássy, I. Free-Living Nematodes of HUNGARY, III (Nematoda errantia). Pedozoologica Hungarica No. 5; Hungarian Natural History Museum: Budapest, Hungary, 2009; 608p. [Google Scholar]
- Barker, K.R. Nematode extraction and bioassays. In An Advanced Treatise on Meloidogyne, Volume II. Methodology; Barker, K.R., Carter, C.C., Sasser, J.N., Eds.; North Carolina State University: Raleigh, NC, USA, 1985; pp. 19–35. [Google Scholar]
- Siddiqi, M.R. Studies on Discolaimus spp. (Nematoda: Dorylaimidae) from India. Z. Zool. Syst. Evol. 1964, 2, 174–184. [Google Scholar] [CrossRef]
- Álvarez-Ortega, S.; Peña-Santiago, R. Aporcella charidemiensis sp. n. (Dorylaimida: Aporcelaimidae) from the southern Iberian Peninsula, with comments on the phylogeny of the genus. Nematology 2016, 18, 811–821. [Google Scholar] [CrossRef]
- Álvarez-Ortega, S.; Subbotin, S.A.; Peña-Santiago, R. Morphological and molecular characterisation of Aporcelaimellus simplex (Thorne & Swanger, 1936) Loof & Coomans, 1970 and a new concept for Aporcella Andrássy, 2002 (Dorylaimida: Aporcelaimidae). Nematology 2013, 15, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Subbotin, S.A.; Sturhan, D.; Chizhov, V.N.; Vovlas, N.; Baldwin, J.G. Phylogenetic analysis of Tylenchida Thorne, 1949 as inferred from D2 and D3 expansion fragments of the 28S rRNA gene sequences. Nematology 2006, 8, 455–474. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Holterman, M.; Rybarczyk, K.; van den Elsen, S.; van Megen, H.; Mooyman, P.; Peña-Santiago, R.; Bongers, T.; Bakker, J.; Helder, J. A ribosomal DNA-based framework for the detection and quantification of stress-sensitive nematode families in terrestrial habitats. Mol. Ecol. Resour. 2008, 8, 23–34. [Google Scholar] [CrossRef]
- Álvarez-Ortega, S.; Peña-Santiago, R. Morphology, phylogeny and taxonomy of the genus Sectonema (Nematoda, Aporcelaimidae). Zool. Scr. 2019, 48, 535–544. [Google Scholar] [CrossRef]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Heyns, J. New species of the superfamily Dorylaimoidea (Nemata) from South Africa soil with a description of the genus Kochinema. S. Afr. J. Agric. Sci. 1963, 6, 289–302. [Google Scholar]
- Siddiqi, M.R. Seven new species of Dorylaimoidea (Nematoda) from India, with description of Lenonchium n. gen. and Galophinema n. gen. Proc. Helm. Soc. Wash. 1965, 32, 81–90. [Google Scholar]
- Argo, A.O.; Van den Berg, E. Three new species of the genus Kochinema (Nematoda: Dorylaimoidea) from South Africa. Phytophylactica 1971, 3, 45–50. [Google Scholar]
- Siddiqi, M.R. Crateronema n. gen. (Crateronematidae n. fam.), Poronemella n. gen. (Lordellonematinae n. sub. fam) and Chrysonemoides n. gen. (Chrysonematidae n. fam.) with a revised classification of Dorylaimoidea (Nematoda). Nematologica 1969, 15, 18–100. [Google Scholar] [CrossRef] [Green Version]
- Darekar, K.S.; Khan, E. Soil and plant-parasitic nematodes from Maharashtra, India. VII. Indokochinema conicauda n. gen., n. sp. and Kochinematidae n. fam. (Dorylaimida: Nematoda). Indian J. Nematol. 1979, 8, 140–143. [Google Scholar]
- Jairajpuri, M.S.; Ahmad, W. Dorylaimida—Free-Living, Predaceous and Plant-Parasitic Nematodes; Brill: Leiden, The Netherlands, 1992; 458p. [Google Scholar]
- Jana, A.; Baqri, Q.H. Nematodes from West Bengal (India) XV. On the species of some rare genera having narrow odontostyle of the super family Dorylaimoidea (Dorylaimida). Bull. Zool. Surv. India 1985, 7, 297–304. [Google Scholar]
- Baqri, Q.H.; Bohra, P. Nematodes from Rajasthan, India. 1. Six new species of Dorylaimida. Nematology 2001, 3, 113–127. [Google Scholar] [CrossRef]
- De Ley, P.; Coomans, A. Terrestrial nematodes from the Galápagos Archipelago. 11. The morphology of three rare Dorylaimoidea (Dorylaimida). J. Nematode Morphol. Syst. 2013, 16, 101–109. [Google Scholar]
- Vazifeh, N.; Niknam, G.; Jabbari, H. Report of five dorylaimid nematodes from East Azerbaijan province, Iran. Iran. J. Plant Pathol. 2018, 49, 203–215. [Google Scholar] [CrossRef]
- Pedram, M.; Pourjam, E.; Robbins, R.T.; Ye, W.; Peña-Santiago, R. Description of Rhyssocolpus vinciguerrae sp. n. (Dorylaimida: Nordiidae) from Iran and the first molecular study of this genus. Nematology 2011, 13, 927–937. [Google Scholar] [CrossRef]
- Peña-Santiago, R.; Guerrero, P.; Liébanas, G.; García, M.C.; Palomeque, T.; Lorite, P. Characterisation of an Iberian population of Rhyssocolpus iuventutis Andrássy, 1971 (Dorylaimida: Nordiidae), with a revised taxonomy of the genus. Nematology 2015, 17, 139–153. [Google Scholar] [CrossRef]
- Elshishka, M.; Lazarova, S.; Radoslavov, G.; Hristov, P.; Peneva, V.K. New data on two remarkable Antarctic species Amblydorylaimus isokaryon (Loof, 1975) Andrássy, 1998 and Pararhyssocolpus paradoxus (Loof, 1975), gen. n., comb. n. (Nematoda: Dorylaimida). ZooKeys 2015, 511, 25–68. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | El Dorado County, California, USA | |
|---|---|---|
| Character * | n | 7 Females |
| L | 1.23 ± 0.07 (1.13–1.30) | |
| a | 42.7 ± 1.8 (41–45) | |
| b | 4.7 ± 0.4 (4.3–5.1) | |
| c | 38.6 ± 4.1 (35–44) | |
| c′ | 1.8 ± 0.2 (1.5–1.9) | |
| V | 52.9 ± 1.5 (51–55) | |
| Lip region diameter | 10.9 ± 0.2 (10.5–11.0) | |
| Odontostyle length | 17.9 ± 0.2 (17.5–18.0) | |
| Odontophore length | 19.8 ± 0.8 (19–21) | |
| Guiding ring from anterior end | 9.5 ± 0.4 (9–10) | |
| Neck length | 266 ± 21.3 (252–298) | |
| Pharyngeal expansion length | 105 ± 9.6 (98–122) | |
| Diameter at neck base | 27.6 ± 1.2 (26.0–29.5) | |
| at midbody | 28.9 ± 1.1 (27.5–30.0) | |
| at anus | 18.1 ± 0.9 (17–19) | |
| Prerectum length | 102 ± 21.5 (80–123) | |
| Rectum length | 24.5 ± 1.5 (22–26) | |
| Tail length | 32.4 ± 3.1 (29–37) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez-Ortega, S. Morphological and Molecular Characterization of Kochinema farodai Baqri and Bohra, 2001 (Dorylaimida: Nordiidae) from California, with the First Molecular Study and an Updated Taxonomy of the Genus. Animals 2020, 10, 2300. https://doi.org/10.3390/ani10122300
Álvarez-Ortega S. Morphological and Molecular Characterization of Kochinema farodai Baqri and Bohra, 2001 (Dorylaimida: Nordiidae) from California, with the First Molecular Study and an Updated Taxonomy of the Genus. Animals. 2020; 10(12):2300. https://doi.org/10.3390/ani10122300
Chicago/Turabian StyleÁlvarez-Ortega, Sergio. 2020. "Morphological and Molecular Characterization of Kochinema farodai Baqri and Bohra, 2001 (Dorylaimida: Nordiidae) from California, with the First Molecular Study and an Updated Taxonomy of the Genus" Animals 10, no. 12: 2300. https://doi.org/10.3390/ani10122300