Effects of Oral Glutamine Supplementation on Early Postnatal Muscle Morphology in Low and Normal Birth Weight Piglets

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sampling
2.2. Measurement of Free Amino Acid Concentrations within M. Longissimus
2.3. Histology and Histochemistry
2.4. Immunohistochemistry
2.5. Western Blotting
2.6. Statistical Analysis
3. Results
3.1. Slaughter Weights
3.2. Concentrations of Free Amino Acids within M. Longissimus
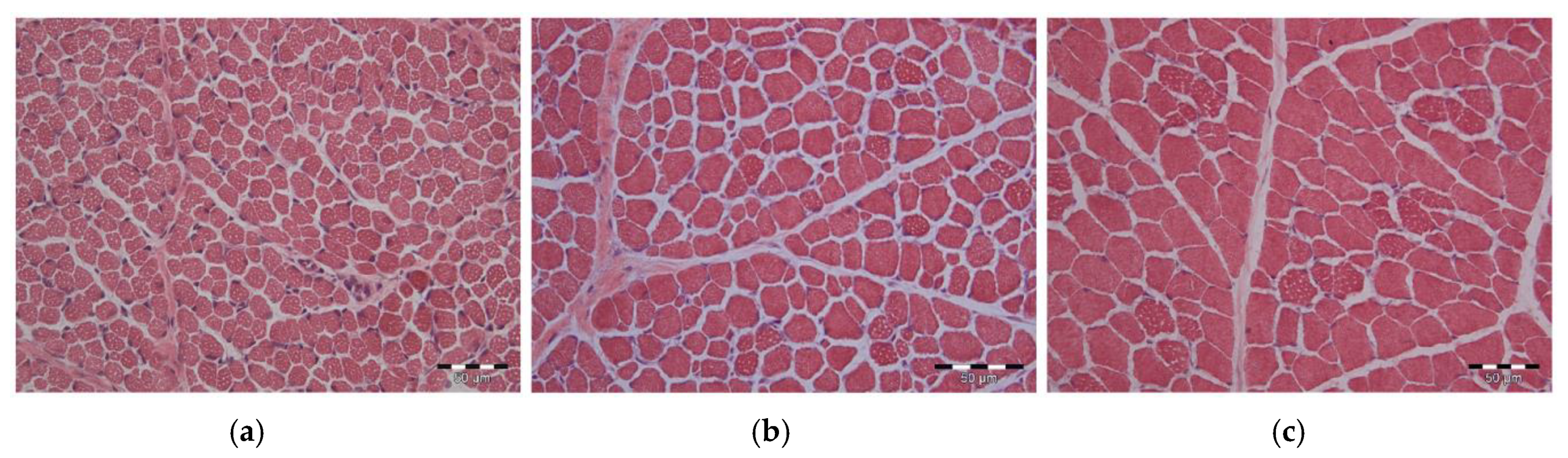
3.3. Muscle Morphology and Capillarization
3.4. Abundance of MYH Isoforms
3.5. Intramuscular Lipid Deposition
4. Discussion
4.1. Intramuscular Amino Acid Concentrations Were Altered by Oral Gln Supplementation
4.2. Muscle Fiber Size Was Influenced by Gln Supplementation
4.3. Abundance of MYH Isoforms Was Slightly Altered by Gln Supplementation
4.4. Intramyocellular Lipid Droplets Were More Abundant in LBW Piglets
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Vos, M.; Che, L.; Huygelen, V.; Willemen, S.; Michiels, J.; Van Cruchten, S.; Van Ginneken, C. Nutritional interventions to prevent and rear low-birthweight piglets. J. Anim. Physiol. Anim. Nutr. 2014, 98, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Spilsbury, M.; Ramirez-Necoechea, R.; Gonzalez-Lozano, M.; Mota-Rojas, D.; Trujillo-Ortega, M.E. Piglet survival in early lactation: A review. J. Anim. Vet. Adv. 2007, 6, 76–86. [Google Scholar]
- D’Inca, R.; Gras-Le Guen, C.; Che, L.Q.; Sangild, P.T.; Le Huerou-Luron, I. Intrauterine growth restriction delays feeding-induced gut adaptation in term newborn pigs. Neonatology 2011, 99, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Meas, T.; Deghmoun, S.; Alberti, C.; Carreira, E.; Armoogum, P.; Chevenne, D.; Levy-Marchal, C. Independent effects of weight gain and fetal programming on metabolic complications in adults born small for gestational age. Diabetologia 2010, 53, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Krueger, R.; Derno, M.; Goers, S.; Metzler-Zebeli, B.U.; Nuernberg, G.; Martens, K.; Pfuhl, R.; Nebendahl, C.; Zeyner, A.; Hammon, H.M.; et al. Higher body fatness in intrauterine growth retarded juvenile pigs is associated with lower fat and higher carbohydrate oxidation during ad libitum and restricted feeding. Eur. J. Nutr. 2014, 53, 583–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegarty, P.V.; Allen, C.E. Effect of pre-natal runting on the post-natal development of skeletal muscles in swine and rats. J. Anim. Sci. 1978, 46, 1634–1640. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Maternal nutrition and fetal development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Johnson, G.A.; Knabe, D.A.; Burghardt, R.C.; Spencer, T.E.; Li, X.L.; Wang, J.J. Triennial Growth Symposium: Important roles for L-glutamine in swine nutrition and production. J. Anim. Sci. 2011, 89, 2017–2030. [Google Scholar] [CrossRef] [Green Version]
- Curi, R.; Lagranha, C.J.; Doi, S.Q.; Sellitti, D.F.; Procopio, J.; Pithon-Curi, T.C.; Corless, M.; Newsholme, P. Molecular mechanisms of glutamine action. J. Cell Physiol. 2005, 204, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336 Pt 1, 1–17. [Google Scholar] [CrossRef]
- Marc Rhoads, J.; Wu, G. Glutamine, arginine, and leucine signaling in the intestine. Amino Acids 2009, 37, 111–122. [Google Scholar] [CrossRef]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Bazer, F.W.; Dai, Z.; Li, D.; Wang, J.; Wu, Z. Amino acid nutrition in animals: Protein synthesis and beyond. Annu. Rev. Anim. Biosci. 2014, 2, 387–417. [Google Scholar] [CrossRef]
- Berard, J.; Kreuzer, M.; Bee, G. In large litters birth weight and gender is decisive for growth performance but less for carcass and pork quality traits. Meat Sci. 2010, 86, 845–851. [Google Scholar] [CrossRef]
- Losel, D.; Kalbe, C.; Rehfeldt, C. L-Carnitine supplementation during suckling intensifies the early postnatal skeletal myofiber formation in piglets of low birth weight. J. Anim. Sci. 2009, 87, 2216–2226. [Google Scholar] [CrossRef] [PubMed]
- Spray, C.M.; Widdowson, E.M. The effect of growth and development on the composition of mammals. Br. J. Nutr. 1950, 4, 332–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morise, A.; Louveau, I.; Le Huerou-Luron, I. Growth and development of adipose tissue and gut and related endocrine status during early growth in the pig: Impact of low birth weight. Animal 2008, 2, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Farnworth, E.R.; Kramer, J.K.G. Fat-Metabolism in Growing Swine—A Review. Can. J. Anim. Sci. 1987, 67, 301–318. [Google Scholar] [CrossRef]
- Dodson, M.V.; Jiang, Z.; Du, M.; Hausman, G.J. Adipogenesis: It is not just lipid that comprises adipose tissue. J. Genom. 2013, 1, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, T.A.; Fiorotto, M.L. Regulation of muscle growth in neonates. Curr. Opin. Clin. Nutr. 2009, 12, 78–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardo, C.E.; Berard, J.; Kreuzer, M.; Bee, G. Intrauterine crowding impairs formation and growth of secondary myofibers in pigs. Animal 2013, 7, 430–438. [Google Scholar] [CrossRef] [Green Version]
- Rehfeldt, C.; Lang, I.S.; Gors, S.; Hennig, U.; Kalbe, C.; Stabenow, B.; Brussow, K.P.; Pfuhl, R.; Bellmann, O.; Nurnberg, G.; et al. Limited and excess dietary protein during gestation affects growth and compositional traits in gilts and impairs offspring fetal growth. J. Anim. Sci. 2011, 89, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Amdi, C.; Krogh, U.; Flummer, C.; Oksbjerg, N.; Hansen, C.F.; Theil, P.K. Intrauterine growth restricted piglets defined by their head shape ingest insufficient amounts of colostrum. J. Anim. Sci. 2013, 91, 5605–5613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haynes, T.E.; Li, P.; Li, X.; Shimotori, K.; Sato, H.; Flynn, N.E.; Wang, J.; Knabe, D.A.; Wu, G. L-Glutamine or L-alanyl-L-glutamine prevents oxidant- or endotoxin-induced death of neonatal enterocytes. Amino Acids 2009, 37, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Nebendahl, C.; Kruger, R.; Gors, S.; Albrecht, E.; Martens, K.; Hennig, S.; Storm, N.; Hoppner, W.; Pfuhl, R.; Metzler-Zebeli, B.U.; et al. Effects on transcriptional regulation and lipid droplet characteristics in the liver of female juvenile pigs after early postnatal feed restriction and refeeding are dependent on birth weight. PLoS ONE 2013, 8, e76705. [Google Scholar] [CrossRef] [Green Version]
- Kuhla, B.; Kucia, M.; Görs, S.; Albrecht, D.; Langhammer, M.; Kuhla, S.; Metges, C.C. Effect of a high-protein diet on food intake and liver metabolism during pregnancy, lactation and after weaning in mice. Proteomics 2010, 10, 2573–2588. [Google Scholar] [CrossRef]
- Zitnan, R.; Albrecht, E.; Kalbe, C.; Miersch, C.; Revajova, V.; Levkut, M.; Rontgen, M. Muscle characteristics in chicks challenged with Salmonella Enteritidis and the effect of preventive application of the probiotic Enterococcus faecium. Poult. Sci. 2019, 98, 2014–2025. [Google Scholar] [CrossRef]
- Dahl, N.; Albrecht, E.; Dannenberger, D.; Uken, K.L.; Hammon, H.M.; Maak, S. Consequences of maternal essential fatty acid and conjugated linoleic acid supplementation on the development of calf muscle and adipose tissue. Animals 2020, 10, 1598. [Google Scholar] [CrossRef]
- Liu, Y.; Albrecht, E.; Schering, L.; Kuehn, C.; Yang, R.; Zhao, Z.; Maak, S. Agouti Signaling Protein and Its Receptors as Potential Molecular Markers for Intramuscular and Body Fat Deposition in Cattle. Front Physiol. 2018, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Yin, Y.L.; Chu, W.; Liu, Z.; Deng, D.; Li, T.; Huang, R.; Zhang, J.; Tan, B.; Wang, W.; et al. Dietary arginine supplementation increases mTOR signaling activity in skeletal muscle of neonatal pigs. J. Nutr. 2008, 138, 867–872. [Google Scholar] [CrossRef]
- Tan, B.; Li, X.G.; Kong, X.; Huang, R.; Ruan, Z.; Yao, K.; Deng, Z.; Xie, M.; Shinzato, I.; Yin, Y.; et al. Dietary L-arginine supplementation enhances the immune status in early-weaned piglets. Amino Acids 2009, 37, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; McPherson, R.L.; Wu, G.Y. Dietary arginine supplementation enhances the growth of milk-fed young pigs. FASEB J. 2004, 18, A377–A378. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yin, Y.; Tan, B.; Kong, X.; Wu, G. Leucine nutrition in animals and humans: mTOR signaling and beyond. Amino Acids 2011, 41, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Li, W.; Huang, X.; Wang, Y.; Zhang, L.; Zhou, Y.; Hussain, A.; Wang, T. Effects of glutamine supplementation on the immune status in weaning piglets with intrauterine growth retardation. Arch. Anim. Nutr. 2012, 66, 347–356. [Google Scholar] [CrossRef]
- Chen, S.; Xia, Y.; Zhu, G.; Yan, J.; Tan, C.; Deng, B.; Deng, J.; Yin, Y.; Ren, W. Glutamine supplementation improves intestinal cell proliferation and stem cell differentiation in weanling mice. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef]
- Maclennan, P.A.; Brown, R.A.; Rennie, M.J. A positive relationship between protein synthetic rate and intracellular glutamine concentration in perfused rat skeletal-muscle. FEBS Lett. 1987, 215, 187–191. [Google Scholar] [CrossRef] [Green Version]
- Jepson, M.M.; Bates, P.C.; Broadbent, P.; Pell, J.M.; Millward, D.J. Relationship between glutamine concentration and protein-synthesis in rat skeletal-muscle. Am. J. Physiol. 1988, 255, E166–E172. [Google Scholar] [CrossRef]
- Wu, G.Y.; Thompson, J.R. The effect of glutamine on protein turnover in chick skeletal muscle in vitro. Biochem. J. 1990, 265, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, L.; Li, P.; Li, X.; Zhou, H.; Wang, F.; Li, D.; Yin, Y.; Wu, G. Gene expression is altered in piglet small intestine by weaning and dietary glutamine supplementation. J. Nutr. 2008, 138, 1025–1032. [Google Scholar] [CrossRef] [Green Version]
- Rogero, M.M.; Tirapegui, J.; Pedrosa, R.G.; Pires, I.S.D.; de Castro, I.A. Plasma and tissue glutamine response to acute and chronic supplementation with L-glutamine and L-alanyl-L-glutamine in rats. Nutr. Res. 2004, 24, 261–270. [Google Scholar] [CrossRef]
- Yoo, S.S.; Field, C.J.; McBurney, M.I. Glutamine supplementation maintains intramuscular glutamine concentrations and normalizes lymphocyte function in infected early weaned pigs. J. Nutr. 1997, 127, 2253–2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aristoy, M.C.; Toldrá, F. Concentration of free amino acids and dipeptides in porcine skeletal muscles with different oxidative patterns. Meat Sci. 1998, 50, 327–332. [Google Scholar] [CrossRef]
- Cornet, M.; Bousset, J. Free amino acids and dipeptides in porcine muscles: Differences between ‘red’ and ‘white’ muscles. Meat Sci. 1999, 51, 215–219. [Google Scholar] [CrossRef]
- Kriketos, A.D.; Pan, D.A.; Lillioja, S.; Cooney, G.J.; Baur, L.A.; Milner, M.R.; Sutton, J.R.; Jenkins, A.B.; Bogardus, C.; Storlien, L.H. Interrelationships between muscle morphology, insulin action, and adiposity. Am. J. Physiol. 1996, 270, R1332–R1339. [Google Scholar] [CrossRef] [PubMed]
- Aniansson, A.; Hedberg, M.; Henning, G.B.; Grimby, G. Muscle morphology, enzymatic activity, and muscle strength in elderly men: A follow-up study. Muscle Nerve 1986, 9, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How muscle structure and composition influence meat and flesh quality. Sci. World J. 2016, 2016, 3182746. [Google Scholar] [CrossRef] [Green Version]
- Lefaucheur, L. A second look into fibre typing—Relation to meat quality. Meat Sci. 2010, 84, 257–270. [Google Scholar] [CrossRef]
- Stange, K.; Miersch, C.; Sponder, G.; Rontgen, M. Low birth weight influences the postnatal abundance and characteristics of satellite cell subpopulations in pigs. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Gondret, F.; Lefaucheur, L.; Juin, H.; Louveau, I.; Lebret, B. Low birth weight is associated with enlarged muscle fiber area and impaired meat tenderness of the longissimus muscle in pigs. J. Anim. Sci. 2006, 84, 93–103. [Google Scholar] [CrossRef] [PubMed]
- House, J.D.; Pencharz, P.B.; Ball, R.O. Glutamine supplementation to total parenteral nutrition promotes extracellular fluid expansion in piglets. J. Nutr. 1994, 124, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.D. Histochemical analysis of capillary and fiber-type distributions in skeletal muscles of spontaneously hypertensive rats. Microvasc. Res. 1988, 36, 228–238. [Google Scholar] [CrossRef]
- Degens, H.; Turek, Z.; Hoofd, L.J.; Van’t Hof, M.A.; Binkhorst, R.A. The relationship between capillarisation and fibre types during compensatory hypertrophy of the plantaris muscle in the rat. J. Anat. 1992, 180 (Pt 3), 455–463. [Google Scholar]
- Maxwell, L.C.; White, T.P.; Faulkner, J.A. Oxidative capacity, blood-flow, and capillarity of skeletal-muscles. J. Appl. Physiol. 1980, 49, 627–633. [Google Scholar] [CrossRef]
- Bauer, R.; Gedrange, T.; Bauer, K.; Walter, B. Intrauterine growth restriction induces increased capillary density and accelerated type I fiber maturation in newborn pig skeletal muscles. J. Perinat. Med. 2006, 34, 235–242. [Google Scholar] [CrossRef]
- Bauer, R.; Wank, V.; Walter, B.; Blickhan, R.; Zwiener, U. Reduced muscle vascular resistance in intrauterine growth restricted newborn piglets. Exp. Toxicol. Pathol. 2000, 52, 271–276. [Google Scholar] [CrossRef]
- Mesires, N.T.; Doumit, M.E. Satellite cell proliferation and differentiation during postnatal growth of porcine skeletal muscle. Am. J. Physiol. Cell Physiol. 2002, 282, C899–C906. [Google Scholar] [CrossRef]
- Mulvaney, D.R.; Marple, D.N.; Merkel, R.A. Proliferation of skeletal-muscle satellite cells after castration and administration of testosterone propionate. Exp. Biol. Med. 1988, 188, 40–45. [Google Scholar] [CrossRef]
- Cheek, D.B.; Holt, A.B.; Hill, D.E.; Talbert, J.L. Skeletal muscle cell mass and growth—Concept of deoxyribonucleic acid unit. Pediatr. Res. 1971, 5, 312–328. [Google Scholar] [CrossRef] [Green Version]
- Pette, D.; Staron, R.S. Myosin isoforms, muscle fiber types, and transitions. Microsc. Res. Tech. 2000, 50, 500–509. [Google Scholar] [CrossRef]
- Chang, K.C.; da Costa, N.; Blackley, R.; Southwood, O.; Evans, G.; Plastow, G.; Wood, J.D.; Richardson, R.I. Relationships of myosin heavy chain fibre types to meat quality traits in traditional and modern pigs. Meat Sci. 2003, 64, 93–103. [Google Scholar] [CrossRef]
- Wank, V.; Bauer, R.; Walter, B.; Kluge, H.; Fischer, M.S.; Blickhan, R.; Zwiener, U. Accelerated contractile function and improved fatigue resistance of calf muscles in newborn piglets with IUGR. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R304–R310. [Google Scholar] [CrossRef] [PubMed]
- Hocquette, J.F.; Gondret, F.; Baeza, E.; Medale, F.; Jurie, C.; Pethick, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehfeldt, C.; Kuhn, G. Consequences of birth weight for postnatal growth performance and carcass quality in pigs as related to myogenesis1. J. Anim. Sci. 2006, 84, E113–E123. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Age | LBW- | LBW- | NBW- | NBW- | SE | Age | SE | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ALA | GLN | ALA | GLN | BiW | Sup | BiW × Sup | |||||
| Arg | 5 | 126 | 144 | 134 | 176 | 47 | 145 B | 23 | 0.670 | 0.520 | 0.882 |
| 12 | 534 | 468 | 414 | 526 | 47 | 495 A | 23 | 0.282 | 0.950 | 0.089 | |
| 26 | 131 | 160 | 134 | 92 | 47 | 129 B | 23 | 0.493 | 0.888 | 0.782 | |
| Leu | 5 | 153 | 146 | 133 | 129 | 12 | 140 | 8 | 0.057 | 0.634 | 0.269 |
| 12 | 154 a | 153 a | 114 b | 129 a,b | 13 | 138 | 9 | 0.001 | 0.538 | 0.007 | |
| 26 | 146 | 153 | 143 | 143 | 12 | 146 | 8 | 0.501 | 0.727 | 0.866 | |
| Ile | 5 | 138 | 137 | 124 | 108 | 10 | 127 A | 6 | 0.028 | 0.401 | 0.107 |
| 12 | 82 | 99 | 63 | 71 | 11 | 79 B | 6 | 0.017 | 0.228 | 0.061 | |
| 26 | 113 | 99 | 96 | 97 | 10 | 101 A | 6 | 0.298 | 0.505 | 0.548 | |
| Lys | 5 | 241 | 291 | 180 | 210 | 51 | 231 B | 39 | 0.043 | 0.343 | 0.163 |
| 12 | 299 | 264 | 254 | 251 | 53 | 267 B | 41 | 0.409 | 0.664 | 0.780 | |
| 26 | 444 a,b | 530 a | 396 a,b | 373 b | 49 | 436 A | 36 | 0.004 | 0.490 | 0.010 | |
| Val | 5 | 221 a | 207 a,b | 182 b | 182 a,b | 14 | 198 | 10 | 0.003 | 0.574 | 0.020 |
| 12 | 205 a | 226 a | 159 b | 192 a,b | 13 | 195 | 10 | <.001 | 0.033 | <.001 | |
| 26 | 181 | 16 7 | 175 | 171 | 14 | 173 | 9 | 0.904 | 0.486 | 0.862 | |
| Gln | 5 | 5198 a,b | 6177 a | 4665 b | 5945 a | 357 | 5496 A | 222 | 0.211 | 0.001 | 0.007 |
| 12 | 3682 | 4321 | 4006 | 4336 | 365 | 4086 B | 229 | 0.578 | 0.166 | 0.477 | |
| 26 | 3393 | 2727 | 3047 | 3085 | 348 | 3063 C | 210 | 0.985 | 0.373 | 0.545 | |
| Ala | 5 | 9453 a | 7360 b | 8738 a | 6935 b | 357 | 8122 A | 209 | 0.078 | <.001 | <.001 |
| 12 | 6436 a | 5075 b | 6402 a | 4885 b | 362 | 5700 B | 213 | 0.728 | <.001 | 0.001 | |
| 26 | 3427 | 3533 | 3360 | 3499 | 350 | 3455 C | 199 | 0.876 | 0.730 | 0.986 | |
| Glu | 5 | 3934 | 4175 | 3908 | 3500 | 225 | 3879 A | 140 | 0.070 | 0.700 | 0.101 |
| 12 | 3678 | 3291 | 3523 | 3055 | 230 | 3387 B | 145 | 0.310 | 0.053 | 0.187 | |
| 26 | 2204 | 2227 | 2222 | 2047 | 219 | 2175 C | 133 | 0.675 | 0.732 | 0.904 | |
| Asp | 5 | 702 | 663 | 619 | 588 | 384 | 643 A | 23 | 0.022 | 0.347 | 0.101 |
| 12 | 511 | 518 | 497 | 417 | 391 | 486 B | 24 | 0.094 | 0.335 | 0.149 | |
| 26 | 316 | 365 | 327 | 349 | 75 | 339 C | 22 | 0.938 | 0.349 | 0.789 | |
| Cit | 5 | 252 | 288 | 239 | 267 | 20 | 261 A | 14 | 0.246 | 0.069 | 0.194 |
| 12 | 293 | 323 | 29 6 | 292 | 21 | 301 A | 15 | 0.343 | 0.456 | 0.424 | |
| 26 | 153 | 146 | 138 | 120 | 19 | 139 B | 13 | 0.173 | 0.506 | 0.486 | |
| Orn | 5 | 142 | 175 | 173 | 150 | 38 | 160 B | 24 | 0.931 | 0.891 | 0.855 |
| 12 | 322 a,b | 364 a | 316 a,b | 233 b | 39 | 309 A | 25 | 0.034 | 0.574 | 0.036 | |
| 26 | 154 | 190 | 134 | 99 | 37 | 144 B | 23 | 0.085 | 0.987 | 0.244 | |
| Pro | 5 | 4766 a,b | 5164 a | 4132 a,b | 4099 b | 395 | 4541 A | 293 | 0.003 | 0.588 | 0.019 |
| 12 | 1840 | 2373 | 2165 | 1822 | 410 | 2162 B | 309 | 0.686 | 0.704 | 0.113 | |
| 26 | 1815 | 1804 | 1567 | 1798 | 380 | 1746 B | 273 | 0.645 | 0.761 | 0.918 | |
| Gly | 5 | 7347 | 7002 | 8123 | 7486 | 365 | 7490 B | 244 | 0.032 | 0.146 | 0.074 |
| 12 | 9537 | 9979 | 10284 | 10399 | 376 | 10080 A | 254 | 0.047 | 0.419 | 0.176 | |
| 26 | 7713 | 8161 | 7722 | 7458 | 354 | 7764 B | 229 | 0.234 | 0.795 | 0.394 | |
| β-Ala | 5 | 3051 | 3232 | 2585 | 2601 | 258 | 2867 A | 182 | 0.005 | 0.667 | 0.041 |
| 12 | 1730 | 1685 | 1480 | 1181 | 267 | 1519 B | 191 | 0.053 | 0.462 | 0.191 | |
| 26 | 863 | 531 | 762 | 552 | 249 | 677 C | 170 | 0.835 | 0.267 | 0.709 | |
| Car | 5 | 2774 | 3348 | 3093 | 3551 | 455 | 3192 C | 268 | 0.519 | 0.245 | 0.615 |
| 12 | 8633 | 9615 | 8159 | 9637 | 462 | 9011 B | 275 | 0.577 | 0.007 | 0.050 | |
| 26 | 13738 | 14178 | 13818 | 13613 | 445 | 13837 A | 256 | 0.549 | 0.795 | 0.785 | |
| Anser | 5 | 48 | 55 | 53 | 62 | 10 | 54 C | 8 | 0.362 | 0.313 | 0.592 |
| 12 | 129 b | 141 a,b | 141 a,b | 159 a | 10 | 142 B | 8 | 0.019 | 0.060 | 0.028 | |
| 26 | 268 b | 282 a,b | 296 a | 297 a | 9 | 286 A | 7 | 0.001 | 0.348 | 0.005 | |
| Trait | Age | LBW- | LBW- | NBW- | NBW- | SE | Age | SE | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ALA | GLN | ALA | GLN | BiW | Sup | BiW × Sup | |||||
| Muscle cross sectional area (mm2) | 5 | 238.5 | 250.7 | 300.3 | 327.1 | 33.1 | 279.1 C | 24.4 | 0.004 | 0.492 | 0.030 |
| 12 | 404.6 b | 455.9 a,b | 497.2 a | 510.1 a | 34.4 | 467.0 B | 25.7 | 0.002 | 0.268 | 0.011 | |
| 26 | 635.4 b | 668.2 b | 791.8 a | 784.6 a | 32.0 | 720.0 A | 22.7 | <0.001 | 0.675 | <0.001 | |
| Overall | 426.1 b | 458.3 b | 529.8 a | 540.6 a | 19.2 | ||||||
| Muscle fiber size (µm2) | 5 | 257.1 | 280.0 | 271.0 | 312.4 | 32.5 | 280.1 C | 20.8 | 0.393 | 0.297 | 0.584 |
| 12 | 393.8 | 391.1 | 405.2 | 401.9 | 33.4 | 398.0 B | 21.5 | 0.682 | 0.924 | 0.981 | |
| 26 | 538.5 b | 619.6 a,b | 594.8 a,b | 671.8 a | 31.7 | 606.2 A | 19.6 | 0.048 | 0.014 | 0.021 | |
| Overall | 396.5 b | 430.3 a,b | 423.7 a,b | 462.0 a | 18.8 | ||||||
| Apparent total muscle fiber number (×105) | 5 | 8.9 | 9.6 | 10.9 | 11.4 | 0.9 | 10.2 B | 0.6 | 0.022 | 0.562 | 0.129 |
| 12 | 10.5 | 12.2 | 12.3 | 12.8 | 0.9 | 11.9 A,B | 0.6 | 0.129 | 0.231 | 0.230 | |
| 26 | 11.7 | 10.8 | 14.0 | 11.9 | 0.9 | 12.1 A | 0.5 | 0.042 | 0.100 | 0.063 | |
| Overall | 10.4 | 10.8 | 12.4 | 12.0 | 0.5 | ||||||
| Number of nuclei per muscle fiber | 5 | 0.61 | 0.63 | 0.69 | 0.70 | 0.05 | 0.66 | 0.03 | 0.087 | 0.814 | 0.384 |
| 12 | 0.61 | 0.64 | 0.62 | 0.58 | 0.05 | 0.61 | 0.03 | 0.620 | 0.864 | 0.865 | |
| 26 | 0.62 | 0.69 | 0.73 | 0.74 | 0.05 | 0.70 | 0.03 | 0.075 | 0.422 | 0.239 | |
| Overall | 0.62 | 0.65 | 0.68 | 0.67 | 0.03 | ||||||
| Capillary density (number per mm2) | 5 | 84.1 | 88.8 | 98.9 | 100.3 | 11.3 | 93.0 B | 7.5 | 0.149 | 0.772 | 0.527 |
| 12 | 113.7 | 137.5 | 122.6 | 147.4 | 11.7 | 130.3 A | 7.9 | 0.300 | 0.025 | 0.108 | |
| 26 | 133.7 | 121.5 | 140.8 | 123.8 | 11.0 | 130.0 A | 7.1 | 0.602 | 0.187 | 0.149 | |
| Overall | 110.5 | 115.9 | 120.7 | 123.8 | 6.5 | ||||||
| Capillary size (µm2) | 5 | 81.2 | 87.6 | 86.5 | 99.3 | 9.5 | 88.7 B | 5.9 | 0.298 | 0.290 | 0.500 |
| 12 | 143.7 | 121.9 | 140.2 | 123.5 | 9.8 | 132.3 A | 6.1 | 0.904 | 0.040 | 0.228 | |
| 26 | 100.6 | 113.6 | 93.3 | 96.0 | 9.3 | 100.9 B | 5.6 | 0.129 | 0.404 | 0.334 | |
| Overall | 108.5 | 107.7 | 106.7 | 106.3 | 5.5 | ||||||
| Trait | Age | LBW- | LBW- | NBW- | NBW- | SE | Age | SE | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ALA | GLN | ALA | GLN | BiW | Sup | BiW × Sup | |||||
| Muscle cross sectional area (mm2) | 5 | 302.7 | 275.5 | 307.1 | 279.3 | 54.7 | 291.1 B | 44.3 | 0.900 | 0.509 | 0.928 |
| 12 | 395.0 | 400.0 | 389.4 | 460.3 | 57.3 | 411.1 B | 46.9 | 0.407 | 0.369 | 0.476 | |
| 26 | 605.3 | 543.8 | 644.1 | 659.3 | 52.3 | 613.1 A | 40.9 | 0.022 | 0.613 | 0.076 | |
| Overall | 434.3 | 406.3 | 446.9 | 466.3 | 31.7 | ||||||
| Muscle fiber size (µm2) | 5 | 375.6 | 407.1 | 434.2 | 460.5 | 42.6 | 419.4 C | 25.2 | 0.142 | 0.486 | 0.447 |
| 12 | 541.7 | 576.0 | 563.3 | 594.5 | 43.4 | 568.9 B | 25.8 | 0.598 | 0.439 | 0.830 | |
| 26 | 828.6 | 865.7 | 900.3 | 867.8 | 41.7 | 865.6 A | 24.1 | 0.332 | 0.957 | 0.617 | |
| Overall | 582.0 | 616.3 | 632.6 | 640.9 | 24.6 | ||||||
| Apparent total muscle fiber number (×105) | 5 | 0.39 | 0.41 | 0.53 | 0.57 | 0.09 | 0.47 B | 0.07 | 0.012 | 0.661 | 0.084 |
| 12 | 0.61 | 0.61 | 0.74 | 0.70 | 0.09 | 0.66 B | 0.07 | 0.061 | 0.794 | 0.290 | |
| 26 | 1.20 | 0.98 | 1.17 | 1.13 | 0.08 | 1.12 A | 0.06 | 0.338 | 0.096 | 0.115 | |
| Overall | 0.73 a,b | 0.67 b | 0.81 a | 0.80 a | 0.05 | ||||||
| Number of nuclei per muscle fiber | 5 | 0.55 | 0.54 | 0.56 | 0.71 | 0.05 | 0.59 B | 0.03 | 0.080 | 0.205 | 0.080 |
| 12 | 0.60 | 0.63 | 0.62 | 0.79 | 0.05 | 0.66 B | 0.03 | 0.068 | 0.068 | 0.043 | |
| 26 | 0.80 | 0.79 | 0.85 | 0.78 | 0.05 | 0.81 A | 0.03 | 0.719 | 0.474 | 0.823 | |
| Overall | 0.65 | 0.65 | 0.68 | 0.76 | 0.03 | ||||||
| Capillary density (number per mm2) | 5 | 82.2 | 75.5 | 82.9 | 77.6 | 8.3 | 79.6 | 4.1 | 0.869 | 0.468 | 0.905 |
| 12 | 80.7 | 77.3 | 76.7 | 76.2 | 8.3 | 77.7 | 4.1 | 0.762 | 0.814 | 0.981 | |
| 26 | 73.2 a,b | 84.7 a,b | 98.9 a | 65.8 b | 8.3 | 80.6 | 4.1 | 0.684 | 0.197 | 0.031 | |
| Overall | 78.7 | 79.2 | 86.2 | 73.2 | 4.8 | ||||||
| Capillary size (µm2) | 5 | 200.3 | 211.8 | 230.9 | 225.1 | 17.8 | 217.0 A | 10.3 | 0.177 | 0.869 | 0.543 |
| 12 | 199.1 | 188.6 | 172.8 | 205.5 | 18.1 | 191.5 A | 10.5 | 0.774 | 0.534 | 0.523 | |
| 26 | 135.8 | 109.2 | 114.8 | 128.2 | 17.5 | 122.0 B | 9.8 | 0.948 | 0.711 | 0.646 | |
| Overall | 178.4 | 169.9 | 172.8 | 186.3 | 10.3 | ||||||
| Item | Age | LBW- | LBW- | NBW- | NBW- | SE | Age | SE | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ALA | GLN | ALA | GLN | BiW | Sup | BiW × Sup | |||||
| MYH1 | 5 | 0.49 | 0.48 | 0.52 | 0.51 | 0.07 | 0.50 | 0.05 | 0.643 | 0.855 | 0.968 |
| 12 | 0.64 | 0.59 | 0.46 | 0.57 | 0.07 | 0.57 | 0.05 | 0.074 | 0.599 | 0.140 | |
| 26 | 0.42 | 0.58 | 0.57 | 0.44 | 0.07 | 0.50 | 0.04 | 0.935 | 0.862 | 0.115 | |
| Overall | 0.52 | 0.55 | 0.52 | 0.51 | 0.04 | ||||||
| MYH2 | 5 | 0.51 | 0.51 | 0.53 | 0.54 | 0.06 | 0.52 A | 0.04 | 0.619 | 0.984 | 0.969 |
| 12 | 0.49 | 0.52 | 0.47 | 0.64 | 0.06 | 0.53 A | 0.04 | 0.210 | 0.055 | 0.051 | |
| 26 | 0.35 | 0.36 | 0.42 | 0.39 | 0.06 | 0.38 B | 0.04 | 0.240 | 0.882 | 0.662 | |
| Overall | 0.45 | 0.46 | 0.48 | 0.52 | 0.03 | ||||||
| MYH4 | 5 | 0.31 | 0.35 | 0.23 | 0.30 | 0.05 | 0.30 | 0.03 | 0.144 | 0.305 | 0.360 |
| 12 | 0.29 | 0.23 | 0.23 | 0.22 | 0.05 | 0.24 | 0.03 | 0.411 | 0.487 | 0.698 | |
| 26 | 0.24 | 0.27 | 0.32 | 0.24 | 0.05 | 0.27 | 0.03 | 0.531 | 0.555 | 0.581 | |
| Overall | 0.28 | 0.28 | 0.26 | 0.25 | 0.03 | ||||||
| MYH7 | 5 | 20.31 | 12.73 | 13.95 | 13.02 | 2.58 | 15.00 | 1.83 | 0.117 | 0.065 | 0.035 |
| 12 | 9.44 | 14.26 | 13.96 | 11.60 | 2.62 | 12.32 | 1.92 | 0.628 | 0.579 | 0.262 | |
| 26 | 8.57 | 10.18 | 10.27 | 13.53 | 2.51 | 10.64 | 1.71 | 0.211 | 0.310 | 0.421 | |
| Overall | 12.77 | 12.39 | 12.73 | 12.72 | 1.49 | ||||||
| Item | Age | LBW- | LBW- | NBW- | NBW- | SE | Age | SE | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ALA | GLN | ALA | GLN | BiW | Sup | BiW × Sup | |||||
| MYH1 | 5 | 0.39 | 0.38 | 0.33 | 0.40 | 0.05 | 0.38 | 0.04 | 0.529 | 0.511 | 0.533 |
| 12 | 0.34 | 0.33 | 0.32 | 0.33 | 0.05 | 0.33 | 0.04 | 0.800 | 0.996 | 0.953 | |
| 26 | 0.42 | 0.42 | 0.51 | 0.44 | 0.05 | 0.45 | 0.03 | 0.128 | 0.517 | 0.250 | |
| Overall | 0.38 | 0.38 | 0.39 | 0.39 | 0.03 | ||||||
| MYH2 | 5 | 0.040 | 0.038 | 0.040 | 0.043 | 0.003 | 0.040 A,B | 0.002 | 0.177 | 0.782 | 0.319 |
| 12 | 0.042 | 0.038 | 0.038 | 0.041 | 0.002 | 0.040 B | 0.002 | 0.898 | 0.770 | 0.193 | |
| 26 | 0.046 | 0.045 | 0.045 | 0.049 | 0.002 | 0.046 A | 0.002 | 0.340 | 0.367 | 0.289 | |
| Overall | 0.042 | 0.040 | 0.041 | 0.044 | 0.001 | ||||||
| MYH7 | 5 | 1.29 | 2.52 | 0.83 | 3.30 | 1.22 | 1.98 | 0.95 | 0.842 | 0.062 | 0.248 |
| 12 | 1.11 | 1.27 | 0.92 | 1.54 | 1.27 | 1.21 | 1.00 | 0.962 | 0.696 | 0.971 | |
| 26 | 2.40 b | 3.69 a,b | 7.07 a | 3.52 b | 1.17 | 4.17 | 0.88 | 0.006 | 0.294 | <0.001 | |
| Overall | 1.60 | 2.50 | 2.94 | 2.79 | 0.70 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Albrecht, E.; Sciascia, Q.L.; Li, Z.; Görs, S.; Schregel, J.; Metges, C.C.; Maak, S. Effects of Oral Glutamine Supplementation on Early Postnatal Muscle Morphology in Low and Normal Birth Weight Piglets. Animals 2020, 10, 1976. https://doi.org/10.3390/ani10111976
Zhao Y, Albrecht E, Sciascia QL, Li Z, Görs S, Schregel J, Metges CC, Maak S. Effects of Oral Glutamine Supplementation on Early Postnatal Muscle Morphology in Low and Normal Birth Weight Piglets. Animals. 2020; 10(11):1976. https://doi.org/10.3390/ani10111976
Chicago/Turabian StyleZhao, Yaolu, Elke Albrecht, Quentin L. Sciascia, Zeyang Li, Solvig Görs, Johannes Schregel, Cornelia C. Metges, and Steffen Maak. 2020. "Effects of Oral Glutamine Supplementation on Early Postnatal Muscle Morphology in Low and Normal Birth Weight Piglets" Animals 10, no. 11: 1976. https://doi.org/10.3390/ani10111976