Comparison of MicroRNA Transcriptomes Reveals the Association between MiR-148a-3p Expression and Rumen Development in Goats

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Rumen Tissue Collection
2.2. RNA Isolation, Library Construction, and Sequencing
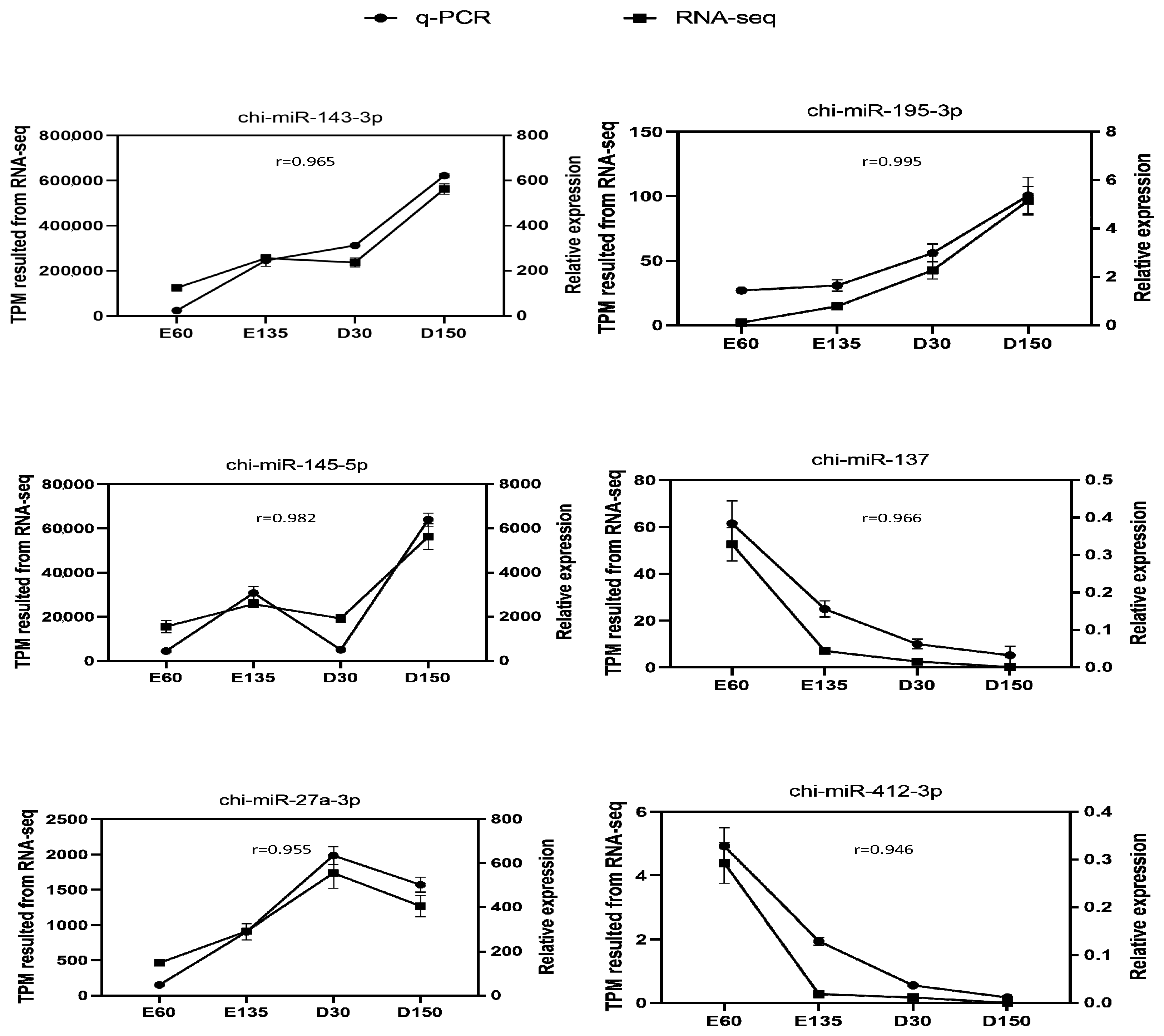
2.3. miRNAs Data Analyses and qPCR Validation
2.4. Expression Vector Construction
2.5. Cell Culture, Transfection, and Expression Detection
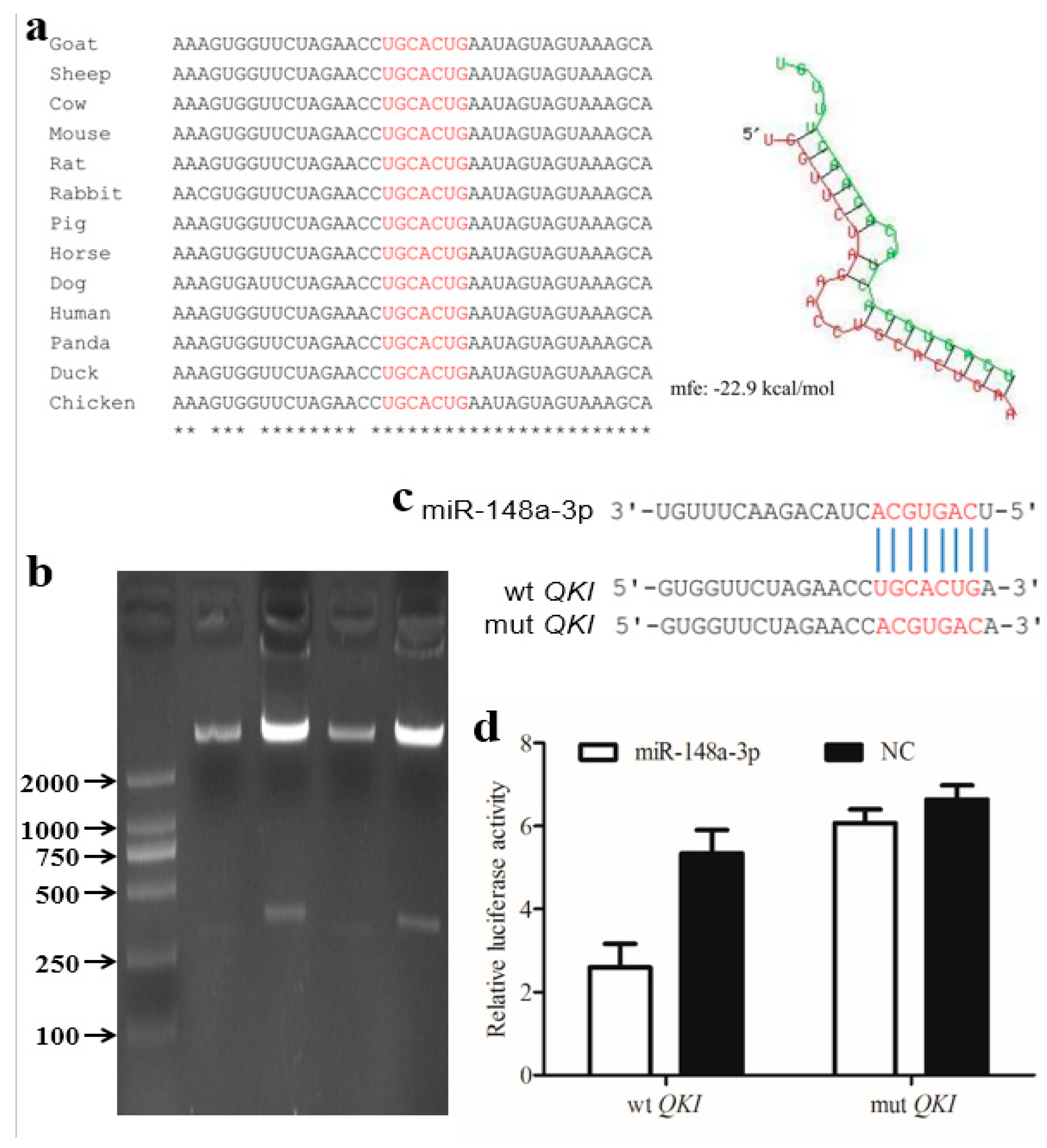
2.6. Luciferase Reporter Assay
2.7. Cell Proliferation Assays
2.8. Dual-Color Fluorescence In Situ Hybridization
2.9. Statistical Analyses
3. Results
3.1. miRNAs Expression Profile and DEMs in Goat Rumens
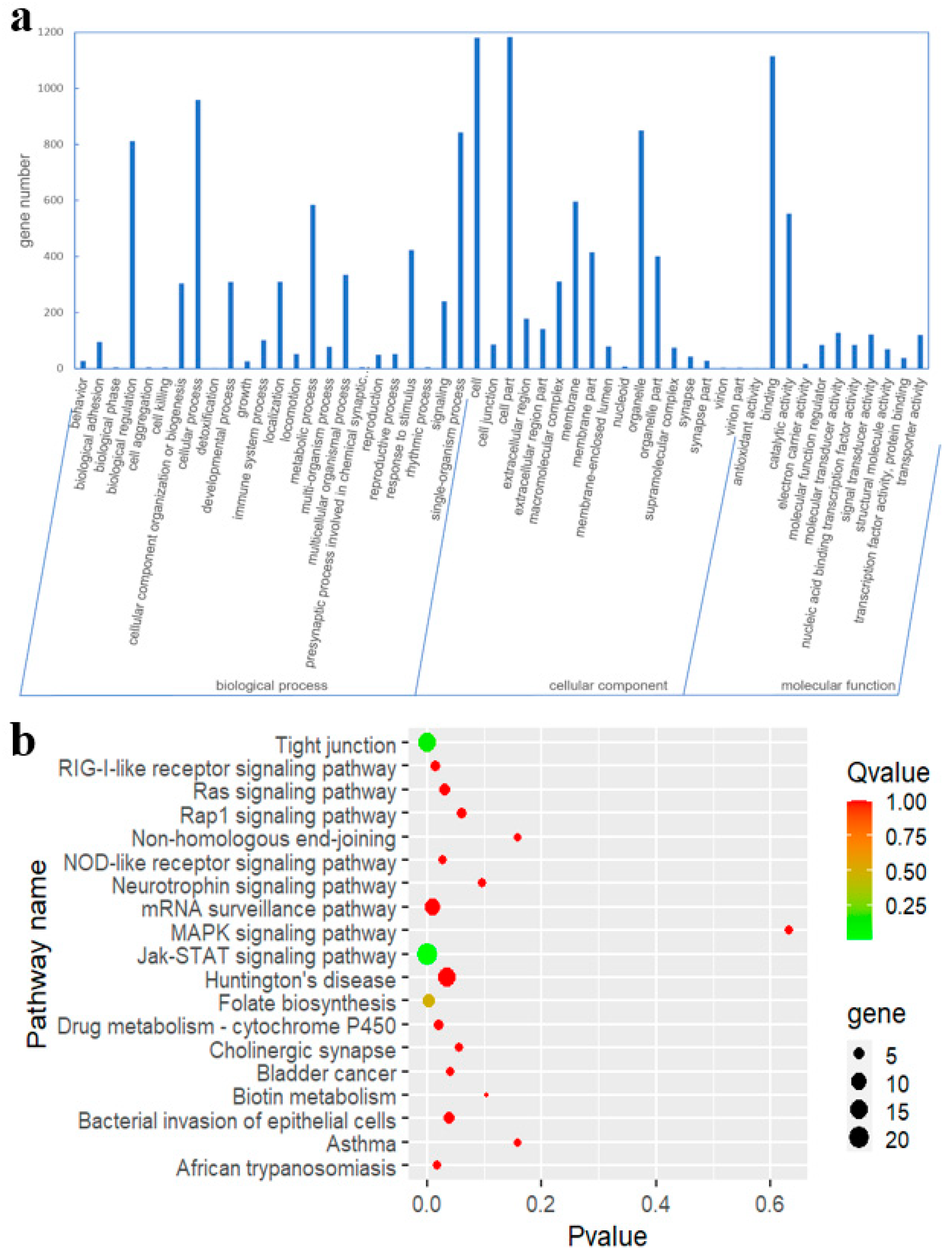
3.2. GO and KEGG Pathway Analyses of Putative Target Genes
3.3. Candidate miR-148a-3p Expression and Target Gene Analysis
3.4. Co-Localization of miR-148a-3p and QKI in Goat Digestive Tissues
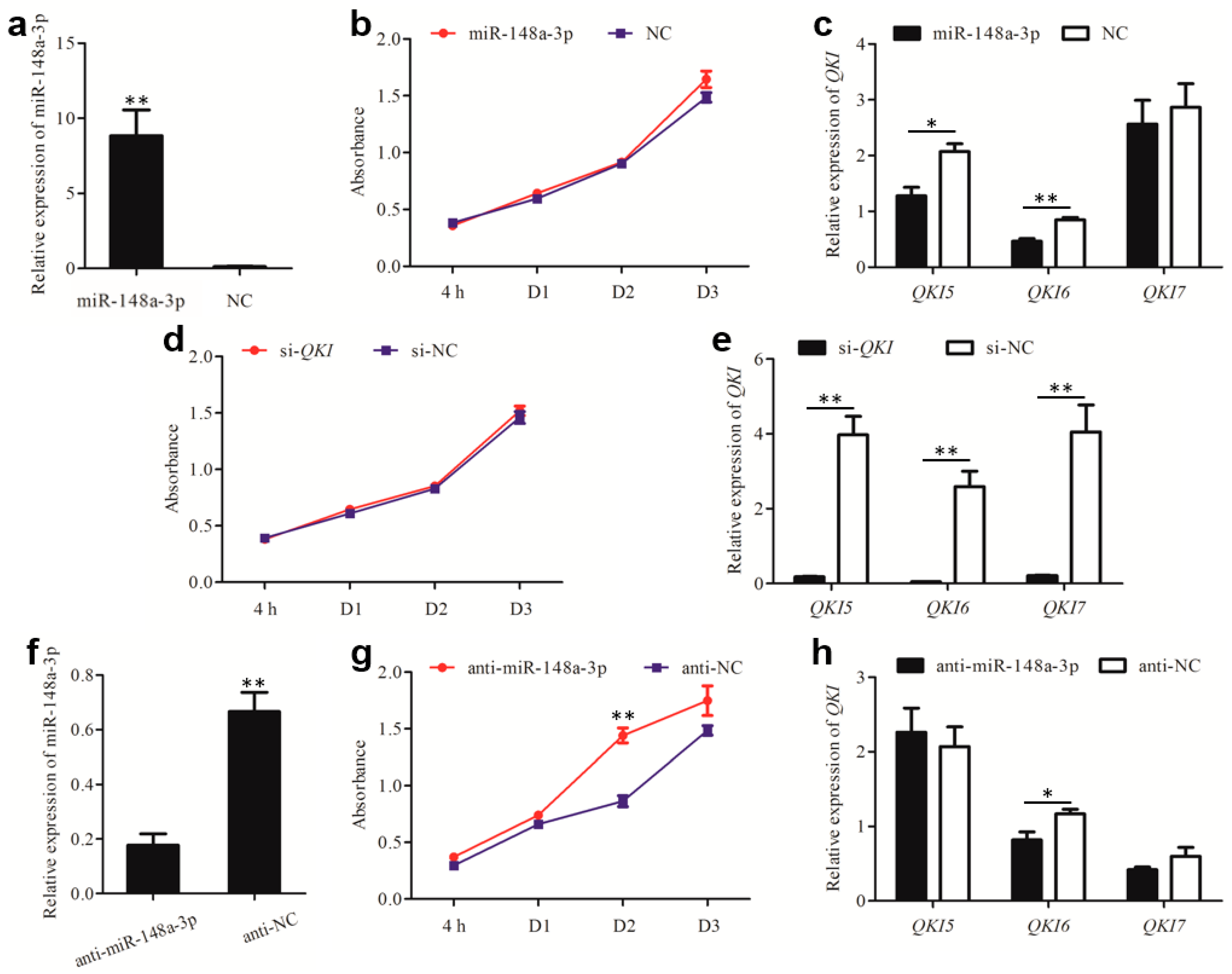
3.5. miR-148a-3p Affects Cell Proliferation by Targeting QKI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Garcia, A.; Masot, J.; Franco, A.; Gazquez, A.; Redondo, E. Histomorphometric and immunohistochemical study of the goat rumen during prenatal development. Anat. Rec. 2012, 295, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Connor, E.E.; Baldwin, R.L.T.; Li, C.J.; Li, R.W.; Chung, H. Gene expression in bovine rumen epithelium during weaning identifies molecular regulators of rumen development and growth. Funct. Integr. Genom. 2013, 13, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.Z.; Xu, Q.; Wang, L.Z.; Wang, J.W.; Guo, W.; Zhou, M.L. The impact of diet on the composition and relative abundance of rumen microbes in goat. Asian-Australas. J. Anim. Sci. 2017, 30, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.P.; Peng, Q.H.; Hu, R.; Zou, H.W.; Wang, H.Z.; Yu, X.Q.; Zhou, J.W.; Degen, A.; Wang, Z.S. Dietary supplements during the cold season increase rumen microbial abundance and improve rumen epithelium development in Tibetan sheep. J. Anim. Sci. 2018, 96, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Carballo, O.C.; Khan, M.A.; Knol, F.W.; Lewis, S.J.; Stevens, D.R.; Laven, R.A.; McCoard, S.A. Impact of weaning age on rumen development in artificially reared lambs. J. Anim. Sci. 2019, 97, 3498–3510. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Shukla, G.C.; Singh, J.; Barik, S. MicroRNAs: Processing, Maturation, Target Recognition and Regulatory Functions. Mol. Cell. Pharmacol. 2011, 3, 83–92. [Google Scholar]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Yuan, C.; Zhang, K.; Yue, Y.J.; Guo, T.T.; Liu, J.B.; Niu, C.N.; Sun, X.P.; Feng, R.L.; Wang, X.L.; Yang, B.H. Analysis of dynamic and widespread lncRNA and miRNA expression in fetal sheep skeletal muscle. PeerJ 2020, 8. [Google Scholar] [CrossRef]
- Sun, L.M.; Lu, S.Y.; Bai, M.; Xiang, L.J.; Li, J.R.; Jia, C.; Jiang, H.Z. Integrative microRNA-mRNA Analysis of Muscle Tissues in Qianhua Mutton Merino and Small Tail Han Sheep Reveals Key Roles for oar-miR-655-3p and oar-miR-381-5p. DNA Cell Biol. 2019, 38, 423–435. [Google Scholar] [CrossRef]
- Ling, Y.H.; Zheng, Q.; Jing, J.; Sui, M.H.; Zhu, L.; Li, Y.S.; Zhang, Y.H.; Liu, Y.; Fang, F.G.; Zhang, X.R. RNA-Seq Reveals miRNA Role Shifts in Seven Stages of Skeletal Muscles in Goat Fetuses and Kids. Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Zhou, G.X.; Wang, X.L.; Yuan, C.; Kang, D.J.; Xu, X.C.; Zhou, J.P.; Geng, R.Q.; Yang, Y.X.; Yang, Z.X.; Chen, Y.L. Integrating miRNA and mRNA Expression Profiling Uncovers miRNAs Underlying Fat Deposition in Sheep. Biomed. Res. Int. 2017, 2017. [Google Scholar] [CrossRef]
- Pan, Y.; Jing, J.; Qiao, L.; Liu, J.; An, L.; Li, B.; Ren, D.; Liu, W. MiRNA-seq reveals that miR-124-3p inhibits adipogenic differentiation of the stromal vascular fraction in sheep via targeting C/EBP alpha. Domest. Anim. Endocrinol. 2018, 65, 17–23. [Google Scholar] [CrossRef]
- Ma, L.A.; Qiu, H.L.; Chen, Z.; Li, L.; Zeng, Y.; Luo, J.; Gou, D.M. miR-25 modulates triacylglycerol and lipid accumulation in goat mammary epithelial cells by repressing PGC-1beta. J. Anim. Sci. Biotechnol. 2018, 9. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, J.J.; He, Q.Y.; Loor, J.J.; Luo, J. Association between the expression of miR-26 and goat milk fatty acids. Reprod. Domest. Anim. 2018, 53, 1478–1482. [Google Scholar] [CrossRef]
- Bai, M.; Sun, L.M.; Jia, C.; Li, J.R.; Han, Y.; Liu, H.; Chen, Y.; Jiang, H.Z. Integrated Analysis of miRNA and mRNA Expression Profiles Reveals Functional miRNA-Targets in Development Testes of Small Tail Han Sheep. G3-Genes Genomes Genet. 2019, 9, 523–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, T.; Hu, J.T.; Xiao, P.; Zhan, S.Y.; Wang, L.J.; Guo, J.Z.; Li, L.; Zhang, H.P.; Niu, L.L. Identification and Characterization of MicroRNAs in the Goat (Capra hircus) Rumen during Embryonic Development. Front. Genet. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Song, C.C.; Yang, J.M.; Jiang, R.; Yang, Z.X.; Li, H.; Huang, Y.Z.; Lan, X.Y.; Lei, C.Z.; Ma, Y.; Qi, X.L.; et al. miR-148a-3p regulates proliferation and apoptosis of bovine muscle cells by targeting KLF6. J. Cell. Physiol. 2019, 234, 15742–15750. [Google Scholar] [CrossRef]
- Yin, H.D.; He, H.R.; Cao, X.A.; Shen, X.X.; Han, S.S.; Cui, C.; Zhao, J.; Wei, Y.H.; Chen, Y.Q.; Xia, L.; et al. MiR-148a-3p Regulates Skeletal Muscle Satellite Cell Differentiation and Apoptosis via the PI3K/AKT Signaling Pathway by Targeting Meox2. Front. Genet. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Vogel, G.; Yu, Z.; Richard, S. The QKI-5 and QKI-6 RNA binding proteins regulate the expression of microRNA 7 in glial cells. Mol. Cell. Biol. 2013, 33, 1233–1243. [Google Scholar] [CrossRef] [Green Version]
- Van Mil, A.; Grundmann, S.; Goumans, M.J.; Lei, Z.; Oerlemans, M.I.; Jaksani, S.; Doevendans, P.A.; Sluijter, J.P. MicroRNA-214 inhibits angiogenesis by targeting Quaking and reducing angiogenic growth factor release. Cardiovasc. Res. 2012, 93, 655–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerez, S.; Araya, H.; Hevia, D.; Irarrazaval, C.E.; Thaler, R.; van Wijnen, A.J.; Galindo, M. Extracellular vesicles from osteosarcoma cell lines contain miRNAs associated with cell adhesion and apoptosis. Gene 2019, 710, 246–257. [Google Scholar] [CrossRef]
- Guo, J.Z.; Zhao, W.; Zhan, S.Y.; Li, L.; Zhong, T.; Wang, L.J.; Dong, Y.; Zhang, H.P. Identification and Expression Profiling of miRNAome in Goat longissimus dorsi Muscle from Prenatal Stages to a Neonatal Stage. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehmsmeier, M.; Steffen, P.; Hochsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Lee, E.J.; Jiang, J.M.; Sarkar, A.; Yang, L.Q.; Elton, T.S.; Chen, C.F. Real-time PCR quantification of precursor and mature microRNA. Methods 2008, 44, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.L.; Yu, H.B.; Mu, S.M.; Dong, Y.D.; Li, D.Y. MiR-26a-5p Inhibits Cell Proliferation and Enhances Doxorubicin Sensitivity in HCC Cells via Targeting AURKA. Technol. Cancer Res. Treat. 2019, 18. [Google Scholar] [CrossRef]
- Yang, Z.H.; Liao, B.; Xiang, X.Y.; Ke, S. miR-21-5p promotes cell proliferation and G1/S transition in melanoma by targeting CDKN2C. FEBS Open Bio 2020, 10, 752–760. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Al-Quraishy, S.; Abdel-Baki, A.A.S.; Delic, D.; Wunderlich, F. Differential miRNA Expression in the Liver of Balb/c Mice Protected by Vaccination during Crisis of Plasmodium chabaudi Blood-Stage Malaria. Front. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- He, H.B.; Cai, M.C.; Zhu, J.Y.; Xiao, W.D.; Liu, B.W.; Shi, Y.; Yang, X.; Liang, X.H.; Zheng, T.H.; Hu, S.Q.; et al. miR-148a-3p promotes rabbit preadipocyte differentiation by targeting PTEN. Cell. Dev. Biol. Anim. 2018, 54, 241–249. [Google Scholar] [CrossRef]
- Mai, M.M.; Jin, L.; Tian, S.L.; Liu, R.; Huang, W.Y.; Tang, Q.Z.; Ma, J.D.; Jiang, A.A.; Wang, X.; Hu, Y.D.; et al. Deciphering the microRNA transcriptome of skeletal muscle during porcine development. PeerJ 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibeagha-Awemu, E.M.; Do, D.N.; Dudemaine, P.L.; Fomenky, B.E.; Bissonnette, N. Integration of lncRNA and mRNA Transcriptome Analyses Reveals Genes and Pathways Potentially Involved in Calf Intestinal Growth and Development during the Early Weeks of Life. Genes 2018, 9, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, D.N.; Dudernaine, P.L.; Fomenky, B.E.; Ibeagha-Awemu, E.M. Integration of miRNA weighted gene co-expression network and miRNA-mRNA co-expression analyses reveals potential regulatory functions of miRNAs in calf rumen development. Genomics 2019, 111, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Htoo, N.N.; Zeshan, B.; Khaing, A.T.; Kyaw, T.; Woldegiorgis, E.A.; Khan, M.A. Creep Feeding Supplemented with Roughages Improve Rumen Morphology in Pre-Weaning Goat Kids. Pak. J. Zool. 2018, 50, 703–709. [Google Scholar] [CrossRef]
- Greco, G.; Hagen, F.; Meissner, S.; Shen, Z.; Lu, Z.; Amasheh, S.; Aschenbach, J.R. Effect of individual SCFA on the epithelial barrier of sheep rumen under physiological and acidotic luminal pH conditions. J. Anim. Sci. 2018, 96, 126–142. [Google Scholar] [CrossRef]
- Jing, X.; Wang, W.; Degen, A.; Guo, Y.; Kang, J.; Liu, P.; Ding, L.; Shang, Z.; Fievez, V.; Zhou, J.; et al. Tibetan sheep have a high capacity to absorb and to regulate metabolism of SCFA in the rumen epithelium to adapt to low energy intake. Br. J. Nutr. 2020, 123, 721–736. [Google Scholar] [CrossRef]
- Xiao, Z.Q.; Majumdar, A.P. Increased in vitro activation of EGFR by membrane-bound TGF-alpha from gastric and colonic mucosa of aged rats. Am. J. Physiol. Gastrointest. Liver. Physiol. 2001, 281, G111–G116. [Google Scholar] [CrossRef] [Green Version]
- Osaki, L.H.; Figueiredo, P.M.; Alvares, E.P.; Gama, P. EGFR is involved in control of gastric cell proliferation through activation of MAPK and Src signalling pathways in early-weaned rats. Cell Prolif. 2011, 44, 174–182. [Google Scholar] [CrossRef]
- Xiao, C.H.; Hong, H.; Yu, H.Z.; Yuan, J.F.; Guo, C.Y.; Cao, H.Y.; Li, W.B. MiR-340 affects gastric cancer cell proliferation, cycle, and apoptosis through regulating SOCS3/JAK-STAT signaling pathway. Immunopharmacol. Immunotoxicol. 2018, 40, 278–283. [Google Scholar] [CrossRef]
- Depeille, P.; Henricks, L.M.; van de Ven, R.A.; Lemmens, E.; Wang, C.Y.; Matli, M.; Werb, Z.; Haigis, K.M.; Donner, D.; Warren, R.; et al. RasGRP1 opposes proliferative EGFR-SOS1-Ras signals and restricts intestinal epithelial cell growth. Nat. Cell Biol. 2015, 17, 804–815. [Google Scholar] [CrossRef] [Green Version]
- Waseem, T.; Duxbury, M.; Ashley, S.W.; Robinson, M.K. Ghrelin promotes intestinal epithelial cell proliferation through PI3K/Akt pathway and EGFR trans-activation both converging to ERK 1/2 phosphorylation. Peptides 2014, 52, 113–121. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNAs | Sequences (5′–3′) | E60 (TPM) | E135 (TPM) | D30 (TPM) | D150 (TPM) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S01 | S02 | S03 | S04 | S05 | S06 | S07 | S08 | S09 | S10 | S11 | S12 | ||
| chi-miR-143-3p | UGAGAUGAAGCACUGUAGCUCG | 129,899 | 108,660 | 133,032 | 282,443 | 236,491 | 248,819 | 169,226 | 283,572 | 256,290 | 587,757 | 615,886 | 483,343 |
| chi-miR-148a-3p | UCAGUGCACUACAGAACUUUGU | 49,694.4 | 63,517.7 | 61,760 | 53,322.6 | 57,020.7 | 73,458.6 | 162,467 | 123,649 | 134,682 | 20,601.2 | 16,858.1 | 41,078.9 |
| chi-miR-21-5p | UAGCUUAUCAGACUGAUGUUGAC | 34,325.5 | 42,267.3 | 40,249.6 | 36,373.6 | 38,252.6 | 35,896.3 | 52,697.9 | 67,017.6 | 55,942.2 | 55,781.8 | 50,200.3 | 56,630.2 |
| chi-miR-26a-5p | UUCAAGUAAUCCAGGAUAGGCU | 42,298.4 | 29,747.9 | 38,170.5 | 37,739 | 42,638.6 | 38,951.3 | 58,679.8 | 50,392.9 | 45,516.7 | 43,799.4 | 40,410.2 | 53,231.2 |
| chi-let-7f-5p | UGAGGUAGUAGAUUGUAUAGUU | 48,221.2 | 37,486.7 | 48,836.2 | 39,702 | 48,107.2 | 43,888 | 42,274.6 | 27,161.3 | 34,006.6 | 18,615.6 | 18,521.2 | 29,367 |
| chi-let-7i-5p | UGAGGUAGUAGUUUGUGCUGUU | 89,488.7 | 59,637.3 | 80,713.4 | 48,114 | 55,336 | 54,656.8 | 103,139 | 72,450 | 83,735.8 | 24,565 | 18,454.3 | 40,592.7 |
| chi-let-7g-5p | UGAGGUAGUAGUUUGUACAGUU | 39,837.1 | 27,269.6 | 33,591.3 | 25,213.5 | 27,818.7 | 25,772 | 39,747.4 | 26,331.8 | 30,419.7 | 18,546.2 | 16,769 | 25,938 |
| chi-miR-10a-5p | UACCCUGUAGAUCCGAAUUUGU | 39,578.2 | 56,845 | 25,375.6 | 46,804.6 | 49,822.8 | 49,492.8 | 16,458.1 | 18,478 | 18,093.6 | 14,346.4 | 8928.24 | 12,395.3 |
| novel_miR_816 | GUGAAAUGUUUAGGACCACUAG | 7716.68 | 10,842.2 | 5830.39 | 43,031.7 | 51,744.6 | 38,203.8 | 25,770.8 | 31,988.9 | 23,516 | 7712.28 | 6032.93 | 10,368.6 |
| chi-miR-27b-3p | UUCACAGUGGCUAAGUUCUGC | 22,255.2 | 22,535.4 | 26,817.6 | 34,768.4 | 35,165 | 32,998.9 | 22,256.4 | 24,188 | 22,918.6 | 18,671 | 19812.2 | 17,478.8 |
| chi-miR-7-5p | UGGAAGACUAGUGAUUUUGUUGUU | 40,591.3 | 12,547.5 | 37,260.2 | 2969.46 | 3014.75 | 3654.06 | 5162.28 | 3850.75 | 2571.84 | 1782.93 | 1498.76 | 2180.65 |
| chi-miR-127-3p | UCGGAUCCGUCUGAGCUUGG | 22,241.9 | 33,430.2 | 22,459.3 | 10,290.6 | 11,178.6 | 11,424.1 | 6808.42 | 9133.77 | 10,262.4 | 266.207 | 233.613 | 1682.39 |
| chi-miR-99a-5p | AACCCGUAGAUCCGAUCUUGU | 21,043.9 | 23,685.3 | 22,250.8 | 21,408.8 | 21,530.7 | 23,416.2 | 29,877.8 | 23,867 | 25,090.2 | 7432.25 | 6398.27 | 13,073.1 |
| chi-miR-1 | UGGAAUGUAAAGAAGUAUGUAU | 6661.52 | 24,793.4 | 3678.91 | 12,652.4 | 12,841.1 | 10,218.5 | 5476.6 | 6946.71 | 8299.96 | 13,206.9 | 12,632.8 | 11,555.9 |
| chi-miR-145-5p | GUCCAGUUUUCCCAGGAAUCCCU | 23,695.1 | 6810.14 | 16,205.4 | 27,621.6 | 26,220.7 | 23,251.9 | 14,500.2 | 18,636.5 | 24,478.5 | 58,611.7 | 72,595 | 37,617.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, T.; Wang, C.; Hu, J.; Chen, X.; Niu, L.; Zhan, S.; Wang, L.; Guo, J.; Cao, J.; Li, L.; et al. Comparison of MicroRNA Transcriptomes Reveals the Association between MiR-148a-3p Expression and Rumen Development in Goats. Animals 2020, 10, 1951. https://doi.org/10.3390/ani10111951
Zhong T, Wang C, Hu J, Chen X, Niu L, Zhan S, Wang L, Guo J, Cao J, Li L, et al. Comparison of MicroRNA Transcriptomes Reveals the Association between MiR-148a-3p Expression and Rumen Development in Goats. Animals. 2020; 10(11):1951. https://doi.org/10.3390/ani10111951
Chicago/Turabian StyleZhong, Tao, Cheng Wang, Jiangtao Hu, Xiaoyong Chen, Lili Niu, Siyuan Zhan, Linjie Wang, Jiazhong Guo, Jiaxue Cao, Li Li, and et al. 2020. "Comparison of MicroRNA Transcriptomes Reveals the Association between MiR-148a-3p Expression and Rumen Development in Goats" Animals 10, no. 11: 1951. https://doi.org/10.3390/ani10111951