Multifactorial Causes of Chronic Mortality in Juvenile Sturgeon (Huso huso)

 , , , , ,
, , , , ,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Pathological Sampling
2.3. Virological Investigation
2.4. Bacteriological Examination
2.5. Chemical Investigations
2.6. Next-Generation Sequencing (NGS): DNA Extraction, Library Preparation, and Sequencing
2.7. NGS: Bioinformatics Analysis
2.8. PCR Confirmation of NGS Data
3. Results
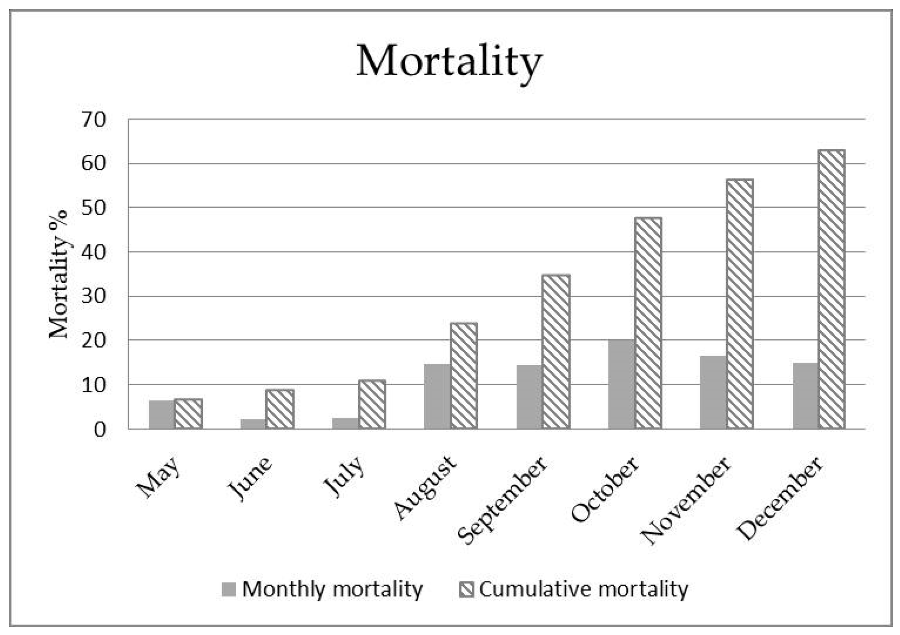
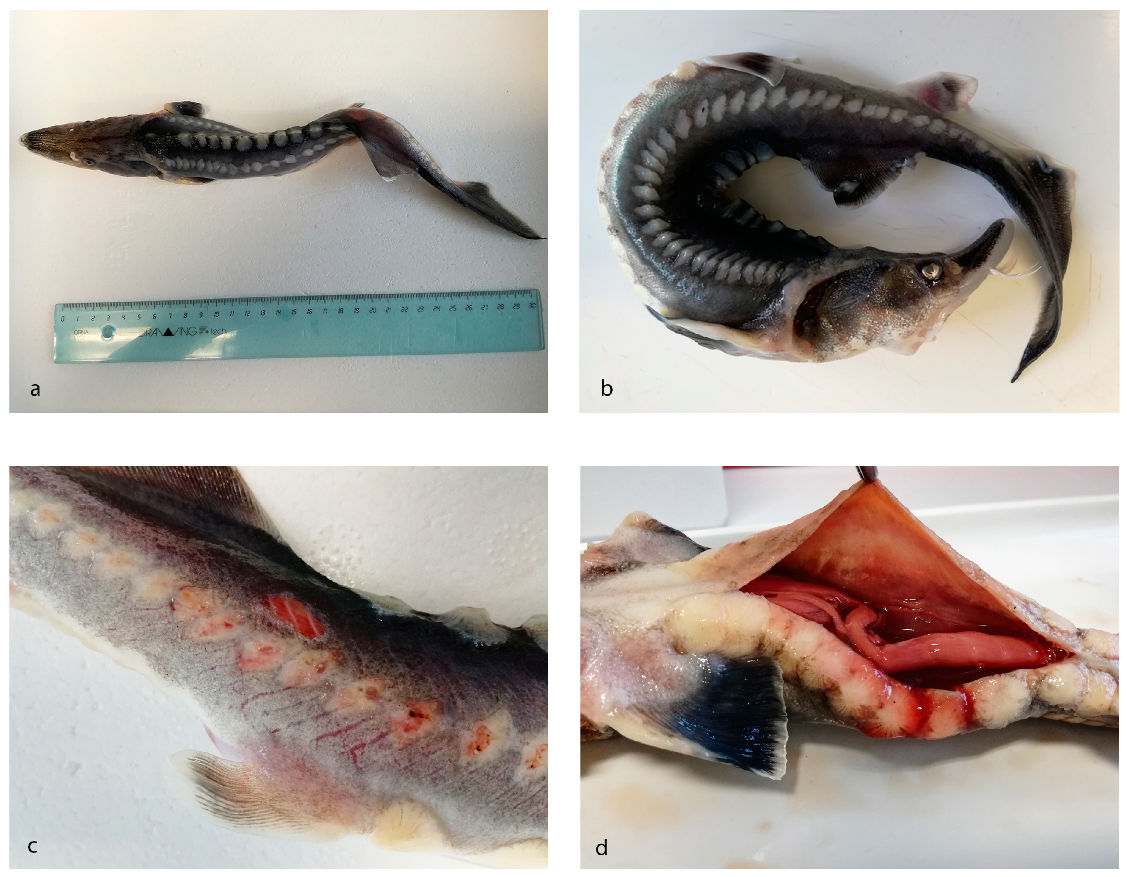
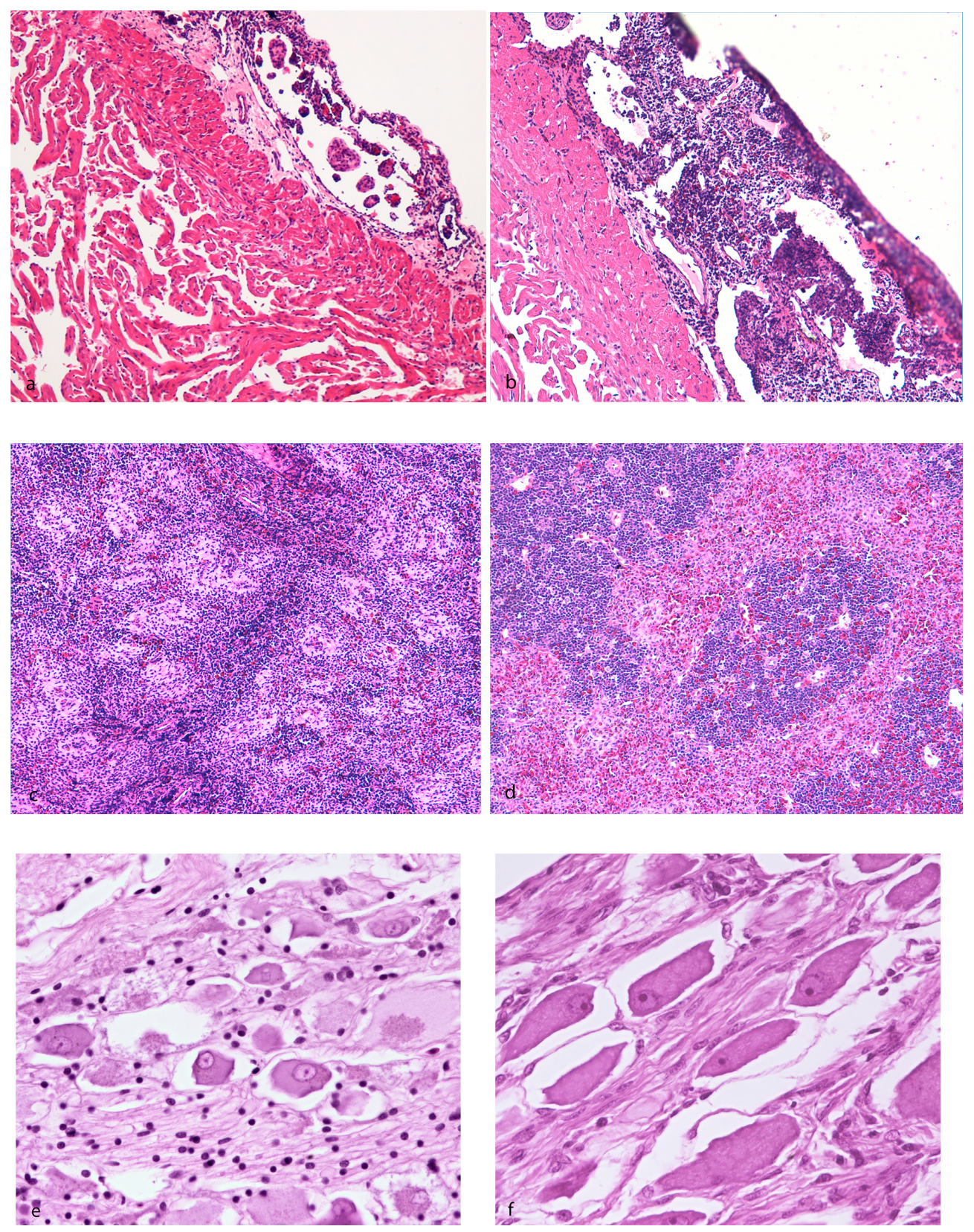
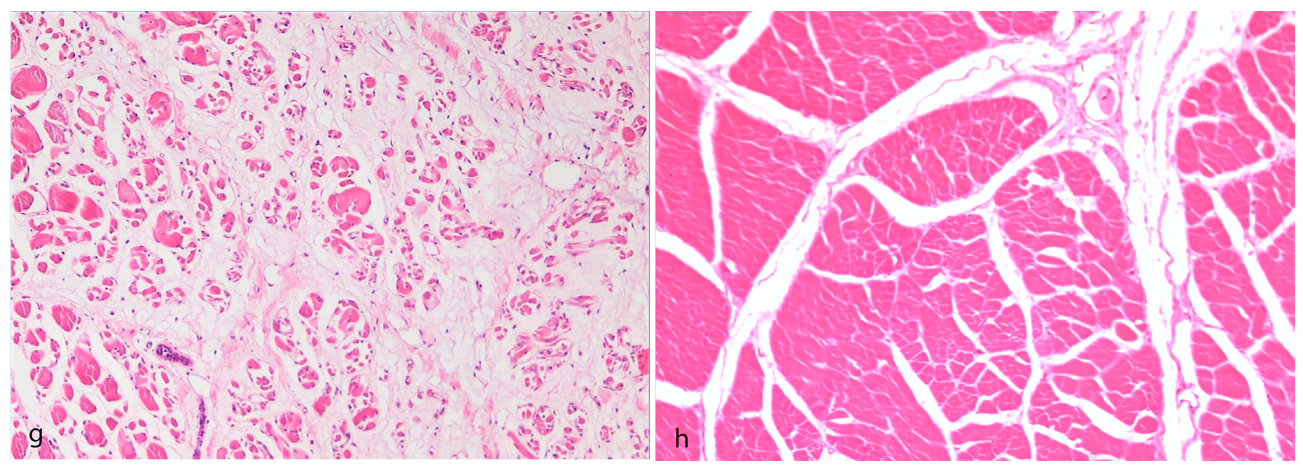
3.1. Pathological Findings
3.2. Virus Investigation
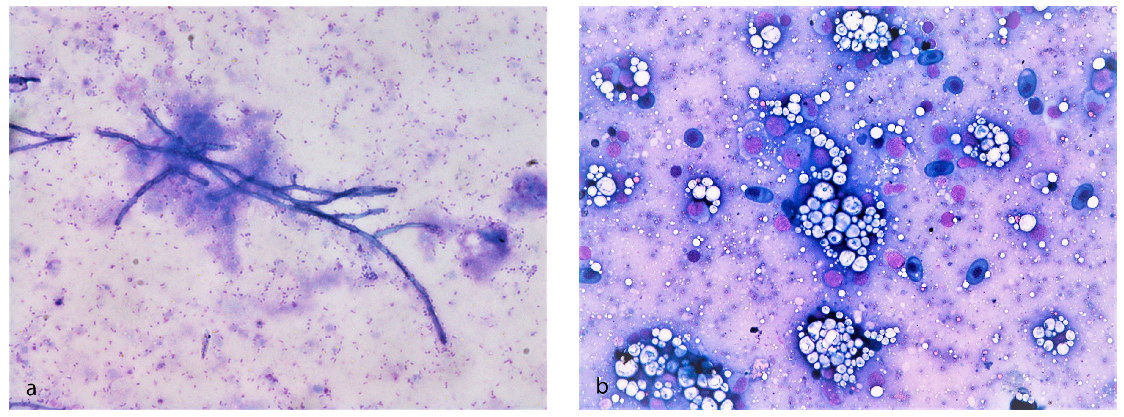
3.3. Bacteriological Examination
3.4. Chemical Investigations
3.5. NGS and Scaffold Analysis
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bronzi, P.; Chebanov, M.; Michaels, J.T.; Wei, Q.; Rosenthal, H.; Gessner, J. Sturgeon meat and caviar production: Global update 2017. J. Appl. Ichthyol. 2019, 35, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Radosavljevic, V.; Milicevic, V.; Maksimović-Zorić, J.; Veljović, L.; Nesic, K.; Pavlović, M.; Ljubojević-Pelić, D.; Marković, Z. Sturgeon diseases in aquaculture. Arch. Vet. Med. 2019, 12. [Google Scholar] [CrossRef]
- Georgiadis, M.P.; Hedrick, R.P.; Johnson, W.O.; Yun, S.; Gardner, I.A. Risk factors for outbreaks of disease attributable to white sturgeon iridovirus and white sturgeon herpesvirus-2 at a commercial sturgeon farm. Am. J. Vet. Res. 2000, 61, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Ciulli, S.; Volpe, E.; Sirri, R.; Passalacqua, P.L.; Cesa Bianchi, F.; Serratore, P.; Mandrioli, L. Outbreak of mortality in Russian (Acipenser gueldenstaedtii) and Siberian (Acipenser baerii) sturgeons associated with sturgeon nucleo-cytoplasmatic large DNA virus. Vet. Microbiol. 2016, 191, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Bigarré, L.; Lesne, M.; Lautraite, A.; Chesneau, V.; Leroux, A.; Jamin, M.; Boitard, P.M.; Toffan, A.; Prearo, M.; Labrut, S.; et al. Molecular identification of iridoviruses infecting various sturgeon species in Europe. J. Fish Dis. 2017, 40, 105–118. [Google Scholar] [CrossRef]
- Axén, C.; Vendramin, N.; Toffan, A. Outbreak of mortality associated with Acipenser Iridovirus European (AcIV-E) detection in siberian sturgeon (Acipenser baerii) farmed in Sweden. Fishes 2018, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Soto, E.; Richey, C.; Stevens, B.; Yun, S.; Kenelty, K.; Reichley, S.; Griffin, M.; Kurobe, T.; Camus, A. Co-infection of Acipenserid herpesvirus 2 (AciHV-2) and Streptococcus iniae in cultured white sturgeon Acipenser transmontanus. Dis. Aquat. Organ. 2017, 124, 11–20. [Google Scholar] [CrossRef]
- Doszpoly, A.; Kalabekov, I.M.; Breyta, R.; Shchelkunov, I.S. Isolation and characterization of an atypical Siberian sturgeon herpesvirus strain in Russia: Novel North American Acipenserid herpesvirus 2 strain in Europe? J. Fish Dis. 2017, 40, 1363–1372. [Google Scholar] [CrossRef]
- Coleman, D.J.; Camus, A.C.; Martínez-López, B.; Yun, S.; Stevens, B.; Soto, E. Effects of temperature on Veronaea botryosa infections in white sturgeon Acipenser transmontanus and fungal induced cytotoxicity of fish cell lines. Vet. Res. 2018, 49, 11. [Google Scholar] [CrossRef] [Green Version]
- Gholamhosseini, A.; Taghadosi, V.; Shiry, N.; Akhlaghi, M.; Sharifiyazdi, H.; Soltanian, S.; Ahmadi, N. First isolation and identification of Aeromonas veronii and Chryseobacterium joostei from reared sturgeons in Fars province, Iran. Vet. Res. Forum Int. Q. J. 2018, 9, 113–119. [Google Scholar] [CrossRef]
- Santi, M.; Pastorino, P.; Foglini, C.; Righetti, M.; Pedron, C.; Prearo, M. A survey of bacterial infections in sturgeon farming in Italy. J. Appl. Ichthyol. 2019, 35, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Ciulli, S.; Volpe, E.; Tura, G.; Zavatta, E.; Renzi, A.; Toffan, A.; Pretto, T.; Mordenti, O.; Errani, F.; Sirri, R.; et al. Multiple co-infections and environmental stressors as causes of chronic mortalities in juvenile sturgeons (Huso huso). In Co-Infections and Multiple Stressors in Fish. Bull. Eur. Assoc. Fish Pathol. 2020, 40, 13–15. [Google Scholar]
- Volpe, E.; Grodzki, M.; Panzarin, V.; Guercio, A.; Purpari, G.; Serratore, P.; Ciulli, S. Detection and molecular characterization of betanodaviruses retrieved from bivalve molluscs. J. Fish Dis. 2018, 41, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Baud, M.; Cabon, J.; Salomoni, A.; Toffan, A.; Panzarin, V.; Bigarré, L. First generic one step real-time Taqman RT-PCR targeting the RNA1 of betanodaviruses. J. Virol. Methods 2015, 211, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Benkó, M.; Elo, P.; Ursu, K.; Ahne, W.; LaPatra, S.E.; Thomson, D.; Harrach, B. First molecular evidence for the existence of distinct fish and snake adenoviruses. J. Virol. 2002, 76, 10056–10059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellehan, J.F.X.; Johnson, A.J.; Harrach, B.; Benkö, M.; Pessier, A.P.; Johnson, C.M.; Garner, M.M.; Childress, A.; Jacobson, E.R. Detection and analysis of six lizard adenoviruses by consensus primer PCR provides further evidence of a reptilian origin for the atadenoviruses. J. Virol. 2004, 78, 13366–13369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, L.A.; Rudis, M.R.; Vasquez-Lee, M.; Montgomery, R.D. A broadly applicable method to characterize large DNA viruses and adenoviruses based on the DNA polymerase gene. Virol. J. 2006, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Caccamo, D.; Di Cello, F.; Fani, R.; Gugliandolo, C.; Maugeri, T.L. Polyphasic approach to the characterisation of marine luminous bacteria. Res. Microbiol. 1999, 150, 221–230. [Google Scholar] [CrossRef]
- Yáñez, M.A.; Catalán, V.; Apráiz, D.; Figueras, M.J.; Martínez-Murcia, A.J. Phylogenetic analysis of members of the genus Aeromonas based on gyrB gene sequences. Int. J. Syst. Evol. Microbiol. 2003, 53, 875–883. [Google Scholar] [CrossRef] [Green Version]
- Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 10 June 2020).
- EPA, U.S. Environmental Protection Agency. Method 3052. Microwave Assisted Acid Digestion of Siliceous and Organically Based MATRICES; U.S. EPA, Office of Solid Waste: Washington, DC, USA, 1996. Available online: http://www.epa.gov/osw/hazard/testmethods/sw8467pdfs/3052.pdf (accessed on 10 December 2019).
- EPA, U.S. Environmental Protection Agency. Method 6010C. Inductively Coupled PlasmaAtomic Emission Spectrometry; U.S. EPA, Office of Solid Waste: Washington, DC, USA, 2007. Available online: http://www.epa.gov/osw/hazard/testmethods/sw846/pdfs/6010c.pdf (accessed on 10 December 2019).
- Simon, A. FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 February 2019).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Sickle v1.33. Available online: https://github.com/najoshi/sickle (accessed on 30 April 2020).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.-J.; Tappu, R. MEGAN Community Edition—Interactive Exploration and Analysis of Large-Scale Microbiome Sequencing Data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Fänge, R. Lymphoid organs in sturgeons (Acipenseridae). Vet. Immunol. Immunopathol. 1986, 12, 153–161. [Google Scholar] [CrossRef]
- Gradil, A.M.; Wright, G.M.; Speare, D.J.; Wadowska, D.W.; Purcell, S.; Fast, M.D. The effects of temperature and body size on immunological development and responsiveness in juvenile shortnose sturgeon (Acipenser brevirostrum). Fish Shellfish Immunol. 2014, 40, 545–555. [Google Scholar] [CrossRef]
- Parsley, M.J.; van der Leeuw, B.K.; Elliott, D.G. Characterization of the Contents and Histology of the Gastrointestinal Tracts of White Sturgeon (Acipenser transmontanus) Captured from Upper Lake Roosevelt, Washington, October 2008; U.S. Geological Survey: Reston, VA, USA, 2010.
- Leprévost, A.; Sire, J.-Y. Architecture, mineralization and development of the axial skeleton in Acipenseriformes, and occurrences of axial anomalies in rearing conditions; can current knowledge in teleost fish help? J. Appl. Ichthyol. 2014, 30, 767–776. [Google Scholar] [CrossRef] [Green Version]
- Athanassopoulou, F.; Billinis, C.; Prapas, T. Important disease conditions of newly cultured species in intensive freshwater farms in Greece: First incidence of nodavirus infection in Acipenser sp. Dis. Aquat. Organ. 2004, 60, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Toffan, A.; Pascoli, F.; Pretto, T.; Panzarin, V.; Abbadi, M.; Buratin, A.; Quartesan, R.; Gijón, D.; Padrós, F. Viral nervous necrosis in gilthead sea bream (Sparus aurata) caused by reassortant betanodavirus RGNNV/SJNNV: An emerging threat for Mediterranean aquaculture. Sci. Rep. 2017, 7, 46755. [Google Scholar] [CrossRef]
- Volpe, E.; Gustinelli, A.; Caffara, M.; Errani, F.; Quaglio, F.; Fioravanti, M.L.; Ciulli, S. Viral nervous necrosis outbreaks caused by the RGNNV/SJNNV reassortant betanodavirus in gilthead sea bream (Sparus aurata) and European sea bass (Dicentrarchus labrax). Aquaculture 2020, 523, 735155. [Google Scholar] [CrossRef]
- Johansen, R.; Sommerset, I.; Tørud, B.; Korsnes, K.; Hjortaas, M.J.; Nilsen, F.; Nerland, A.H.; Dannevig, B.H. Characterization of nodavirus and viral encephalopathy and retinopathy in farmed turbot, Scophthalmus maximus (L.). J. Fish Dis. 2004, 27, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Lama, R.; Pereiro, P.; Valenzuela-Muñoz, V.; Gallardo-Escárate, C.; Tort, L.; Figueras, A.; Novoa, B. RNA-Seq analysis of European sea bass (Dicentrarchus labrax L.) infected with nodavirus reveals powerful modulation of the stress response. Vet. Res. 2020, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Adamek, D.; Rzepkowska, M.; Panagiotopoulou, H.; Fajkowska, M.; Kamaszewski, M.; Kolman, R. Morphological Differences of White Muscle Fibers and Genetic Diversity of Fast and Slow Growing Atlantic Sturgeons (Acipenser oxyrinchus). Turk. J. Fish. Aquat. Sci. 2017, 966, 959–966. [Google Scholar] [CrossRef]
- Cocchi, M.; De Zan, G.; Di Giusto, T.; Deotto, S.; Pretto, T.; Manfrin, A.; Brunetta, R.; Toffan, A. Systemic polymicrobic infection involving Shewanella putrefaciens group in koi. J. Fish Dis. 2018, 41, 1929–1931. [Google Scholar] [CrossRef]
- Mugetti, D.; Pastorino, P.; Menconi, V.; Pedron, C.; Prearo, M. The Old and the New on Viral Diseases in Sturgeon. Pathogens 2020, 9, 146. [Google Scholar] [CrossRef] [Green Version]
- Kayiş, Ş.; Er, A.; Kangel, P.; Kurtoğlu, I.Z. Bacterial pathogens and health problems of Acipenser gueldenstaedtii and Acipenser baerii sturgeons reared in the eastern Black Sea region of Turkey. Iran. J. Vet. Res. 2017, 18, 18–24. [Google Scholar]
- Chen, M.-H.; Hung, S.-W.; Shyu, C.-L.; Lin, C.-C.; Liu, P.-C.; Chang, C.-H.; Shia, W.-Y.; Cheng, C.-F.; Lin, S.-L.; Tu, C.-Y.; et al. Lactococcus lactis subsp. lactis infection in Bester sturgeon, a cultured hybrid of Huso huso × Acipenser ruthenus, in Taiwan. Res. Vet. Sci. 2012, 93, 581–588. [Google Scholar] [CrossRef]
- Safari, R.; Adel, M.; Ghiasi, M.; Asl, M.R.S.; Khalili, E. First isolation and identification of Vibrio vulnificus (biotype 2) from cultured beluga, Huso huso in Iran. Casp. J. Environ. Sci. 2015, 13, 275–281. [Google Scholar]
- Pierezan, F.; Shahin, K.; Heckman, T.I.; Ang, J.; Byrne, B.A.; Soto, E. Outbreaks of severe myositis in cultured white sturgeon (Acipenser transmontanus L.) associated with Streptococcus iniae. J. Fish Dis. 2020, 43, 485–490. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, H.; Li, T.; Luo, L.; Gao, J. Isolation and identification of pathogenic Aeromonas veronii isolated from infected Siberian sturgeon (Acipenser baerii). Wei Sheng Wu Xue Bao 2009, 49, 1289–1294. [Google Scholar]
- Sebastião, F.; Loch, T.P.; Marancik, D.P.; Griffin, M.J.; Maret, J.; Richey, C.; Soto, E. Identification of Chryseobacterium spp. isolated from clinically affected fish in California, USA. Dis. Aquat. Organ. 2019, 136, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Loch, T.P.; Faisal, M. Polyphasic characterization reveals the presence of novel fish-associated Chryseobacterium spp. in the Great Lakes of North America. Dis. Aquat. Organ. 2015, 113, 113–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pękala, A.; Kozińska, A.; Paździor, E.; Głowacka, H. Phenotypical and genotypical characterization of Shewanella putrefaciens strains isolated from diseased freshwater fish. J. Fish Dis. 2015, 38, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Volpe, E.; Mandrioli, L.; Errani, F.; Serratore, P.; Zavatta, E.; Rigillo, A.; Ciulli, S. Evidence of fish and human pathogens associated with doctor fish (Garra rufa, Heckel, 1843) used for cosmetic treatment. J. Fish Dis. 2019, 42, 1637–1644. [Google Scholar] [CrossRef]
- Mehdi, Y.; Hornick, J.-L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, S.S.O. Recent advances in sturgeon nutrition. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2017, 3, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.A.; Govyadinova, A.A.; Khrushchev, N.G. Study on Localization of Hemopoietic Tissue in Sturgeon. Russ. J. Dev. Biol. 2000, 31, 372–376. [Google Scholar] [CrossRef]
- Charmi, A.; Parto, P.; Bahmani, M.; Kazemi, R.; Abbas, B. Morphological and Histological Study of Kidney in Juvenile Great Sturgeon (Huso huso) and Persian Sturgeon (Acipenser persicus). Am. J. Agric. Environ. Sci. 2010, 7, 505–511. [Google Scholar]
- Nagy, É. Chapter 10—Adenoviruses of Fish. In Aquaculture Virology; Kibenge, F.S.B., Godoy, M.G., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 173–176. ISBN 978-0-12-801573-5. [Google Scholar]
- Hedrick, R.P.; Speas, J.; Kent, M.L.; McDowell, T. Adenovirus-Like Particles Associated with a Disease of Cultured White Sturgeon, Acipenser transmontanus. Can. J. Fish. Aquat. Sci. 1985, 42, 1321–1325. [Google Scholar] [CrossRef]
- Noga, E.J. Fish Disease: Diagnosis and Treatment, 2nd ed.; Blackwell Publishing: Ames, IA, USA, 2000; ISBN 9780813806976. [Google Scholar]
- Romanov, A.A.; Lepilina, I.N.; Romanov, A.A. Morphofunctional disturbances in kidneys of sturgeons and bony fishes of the Volga-Caspian reservoir under natural conditions. Tsitologiia 2006, 48, 5–8. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence | Scaffold Orientation | Amplicon (bp) |
|---|---|---|---|
| P1_for | agcctggaacggtatctg | 3′→ 5′ | 273 |
| P1_rev | tacggtgctgtcggt | ||
| p1tq_for | ctttccgcaagcgcagatac | 5′→ 3′ | 331 |
| p1tq_rev | ttcgtcggcggatacgttag | ||
| p1cr_for | ttcgtcggcggatacgttag | 3′→ 5′ | 331 |
| p1cr_rev | cgctgccatcgatttgtgag | ||
| P3tq_for | taacgtatccgccgacgaag | 5′→ 3′ | 325 |
| P3tq_rev | acctccctgtaccgctactt | ||
| P3cr_for | acctccctgtaccgctactt | 3′→ 5′ | 325 |
| P3cr_rev | taacgtatccgccgacgaag |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciulli, S.; Volpe, E.; Sirri, R.; Tura, G.; Errani, F.; Zamperin, G.; Toffan, A.; Silvi, M.; Renzi, A.; Abbadi, M.; et al. Multifactorial Causes of Chronic Mortality in Juvenile Sturgeon (Huso huso). Animals 2020, 10, 1866. https://doi.org/10.3390/ani10101866
Ciulli S, Volpe E, Sirri R, Tura G, Errani F, Zamperin G, Toffan A, Silvi M, Renzi A, Abbadi M, et al. Multifactorial Causes of Chronic Mortality in Juvenile Sturgeon (Huso huso). Animals. 2020; 10(10):1866. https://doi.org/10.3390/ani10101866
Chicago/Turabian StyleCiulli, Sara, Enrico Volpe, Rubina Sirri, Giorgia Tura, Francesca Errani, Gianpiero Zamperin, Anna Toffan, Marina Silvi, Andrea Renzi, Miriam Abbadi, and et al. 2020. "Multifactorial Causes of Chronic Mortality in Juvenile Sturgeon (Huso huso)" Animals 10, no. 10: 1866. https://doi.org/10.3390/ani10101866