An Updated Review of Toxicity Effect of the Rare Earth Elements (REEs) on Aquatic Organisms

 ,
,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Speciation of REEs in Water Bodies
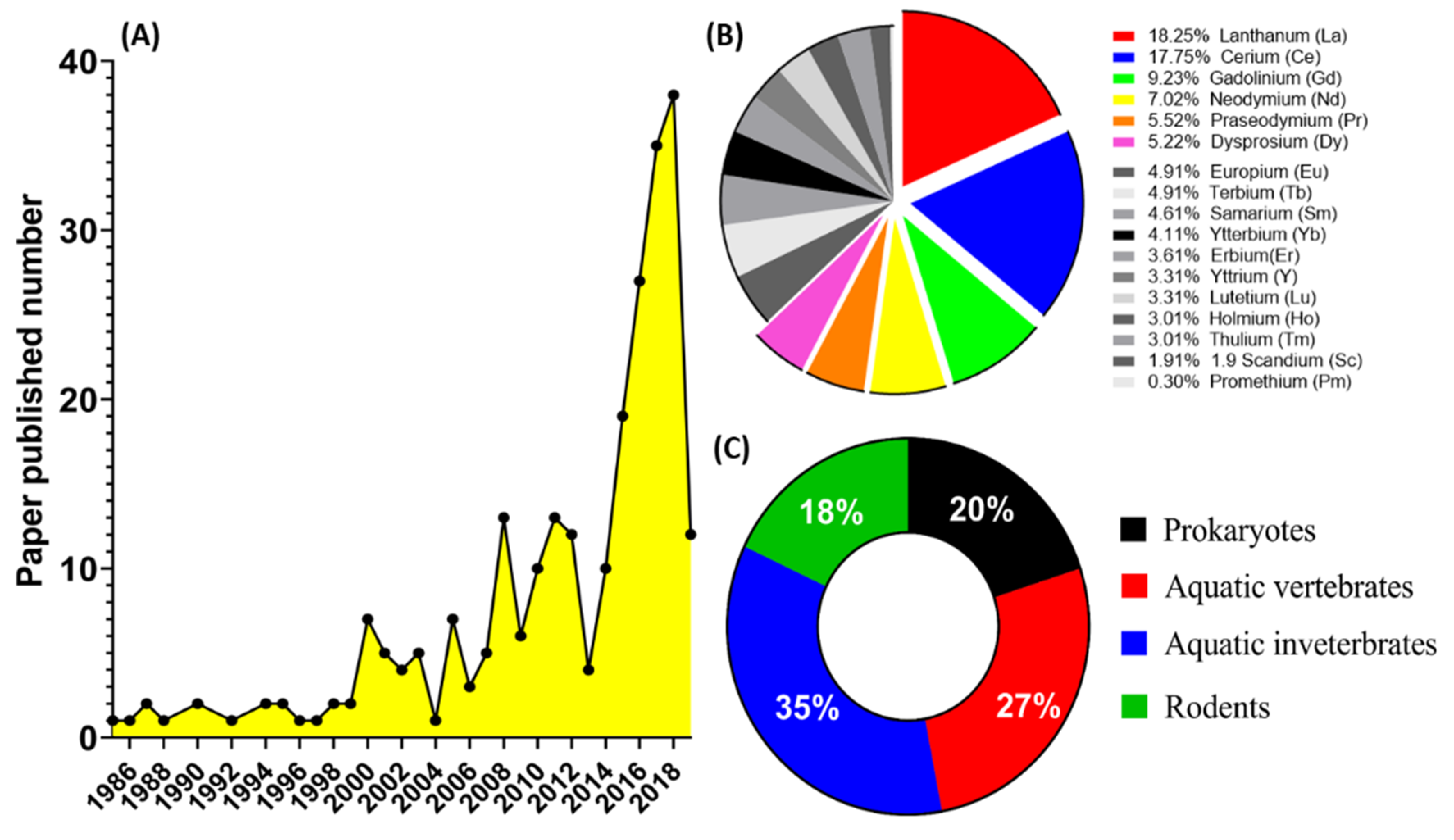
3. Data Collection for Literature Review
4. Overview of Eco-Toxicity of Lanthanides
5. Cumulative Toxicity of REEs, and Malformation in Aquatic Animals
6. Ecotoxicity of Lanthanum (La)
7. Ecotoxicity of Cerium (Ce)
8. Ecotoxicity of Gadolinium (Gd)
9. Ecotoxicity of Neodymium (Nd)
10. Ecotoxicity of Yttrium (Y)
11. Ecotoxicity of Praseodymium (Pr)
12. Ecotoxicity of Samarium (Sm)
13. Ecotoxicity of Dysprosium (Dy)
14. Ecotoxicity of Holmium (Ho)
15. Discussion and Future Research Direction
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AChE | Acetylcholine Esterase |
| ACSV | Adsorptive Cathodic Stripping Voltammetry |
| BLM | Biotic Ligand Model |
| CAT | Catalase |
| Ce | Cerium |
| Cit | Citrate |
| C route | Combustion |
| COX | Cyclooxygenase |
| CO1 | Cytochrome c-oxidase |
| DOC | Dissolved Organic Carbon |
| Dy | Dysprosium |
| EDS | Energy-Dispersive X-ray Spectroscopy |
| Er | Erbium |
| EDTA | Ethylenediaminetetraacetic |
| Eu | Europium |
| Gd | Gadolinium |
| GC-ICP-MS | Gas Chromatography-Inductively Coupled Plasma-Mass Spectrometry |
| GFR | Glomerular Filtration Rate |
| GST | Glutathione-s-transferase |
| HPLC | High- Performance Liquid Chromatography |
| HREEs | High Rare Earth Elements |
| Ho | Holmium |
| H route | Hydrothermal |
| ICP-MS | Inductively Coupled Plasma Mass Spectrometry |
| La | Lanthanum |
| LREEs | Light Rare Earth Elements |
| LPO | Lipid peroxidation |
| LOEC | Lowest Observed Effect Concentration |
| Lu | Lutetium |
| MRI | Magnetic Resonance Imaging |
| MINTEQA | Metal Speciation Equilibrium For Surface And Ground Water |
| MT | Metallothionein |
| MAA | Molecular Activation Analysis |
| NOM | Natural organic matter |
| NSF | Nephrogenic Systemic Fibrosis |
| Nd | Neodymium |
| NTA | Nitrilotriacetic acid |
| NOEC | No Observed Effect Concentration |
| NATs | Nuclear Analytical Techniques |
| OCP | O-cresolphthalexon |
| Pr | Praseodymium |
| Pm | Promethium |
| PIXE | Proton-Induced X-ray Emission |
| REEs | Rare Earth Elements |
| REOs | Rare Earth Oxides |
| Sm | Samarium |
| SEM | Scanning Electron Microscopy |
| SOD | Superoxide Dismutase |
| Tb | Terbium |
| Tm | Thulium |
| USGS | United States Geological Survey |
| XAFS | X-ray Absorption Fine Structure |
| Yb | Ytterbium |
References
- Voncken, J.H.L. The Rare Earth Elements: An Introduction; Springer: Berlin, Germany, 2016. [Google Scholar]
- Gschneidner, K.A., Jr. Rare Earths: The Fraternal Fifteen; US Atomic Energy Commission (AEC): Washington, DC, USA, 1966. [Google Scholar]
- Thyssen, P.; Binnemans, K. Accommodation of the rare earths in the periodic table: A historical analysis. In Handbook on the Physics and Chemistry of Rare Earths; Elsevier: Amsterdam, The Netherlands, 2011; Volume 41, pp. 1–93. [Google Scholar]
- Akeroyd, F.M. Prediction and the periodic table: A response to scerri and worrall. J. Gen. Philos. Sci. 2003, 34, 337. [Google Scholar] [CrossRef]
- Krishnamurthy, N.; Gupta, C.K. Extractive Metallurgy of Rare Earths; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Zhu, Y.; Zheng, Y.; Wang, A. A simple approach to fabricate granular adsorbent for adsorption of rare elements. Int. J. Biol. Macromol. 2015, 72, 410–420. [Google Scholar] [CrossRef]
- Anastopoulos, I.; Bhatnagar, A.; Lima, E.C. Adsorption of rare earth metals: A review of recent literature. J. Mol. Liq. 2016, 221, 954–962. [Google Scholar] [CrossRef]
- Wübbeke, J. Rare earth elements in china: Policies and narratives of reinventing an industry. Resour. Policy 2013, 38, 384–394. [Google Scholar] [CrossRef]
- Massari, S.; Ruberti, M. Rare earth elements as critical raw materials: Focus on international markets and future strategies. Resour. Policy 2013, 38, 36–43. [Google Scholar] [CrossRef]
- Butler, C.J. Rare earth elements: China’s monopoly and implications for us national security. Fletcher Forum World Aff. 2014, 38, 23. [Google Scholar]
- Golev, A.; Scott, M.; Erskine, P.D.; Ali, S.H.; Ballantyne, G.R. Rare earths supply chains: Current status, constraints and opportunities. Resour. Policy 2014, 41, 52–59. [Google Scholar] [CrossRef]
- Jordens, A.; Cheng, Y.P.; Waters, K.E. A review of the beneficiation of rare earth element bearing minerals. Miner. Eng. 2013, 41, 97–114. [Google Scholar] [CrossRef]
- Vereschagin, Y.A.; Kudrevatykh, N.; Malygin, M.; Emelina, T. Rare-earth magnets in russia: Raw materials, processing, properties control and output issues. J. Iron Steel Res. Int. 2006, 13, 23–32. [Google Scholar] [CrossRef]
- Chi, R.; Xu, S.; Zhu, G.; Xu, J.; Qiu, X. Beneficiation of Rare Earth Ore in China. In Light Metals 2001 as Held at the 130th TMS Annual Meeting; The Minerals, Metals & Materials Society: Warrendale, PA, USA, 2001; pp. 1159–1165. [Google Scholar]
- Charalampides, G.; Vatalis, K.I.; Apostoplos, B.; Ploutarch-Nikolas, B. Rare earth elements: Industrial applications and economic dependency of europe. Procedia Econ. Financ. 2015, 24, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Taunton, A.E.; Welch, S.A.; Banfield, J.F. Microbial controls on phosphate and lanthanide distributions during granite weathering and soil formation. Chem. Geol. 2000, 169, 371–382. [Google Scholar] [CrossRef]
- Laveuf, C.; Cornu, S. A review on the potentiality of rare earth elements to trace pedogenetic processes. Geoderma 2009, 154, 1–12. [Google Scholar] [CrossRef]
- Loell, M.; Albrecht, C.; Felix-Henningsen, P. Rare earth elements and relation between their potential bioavailability and soil properties, nidda catchment (Central Germany). Plant Soil 2011, 349, 303–317. [Google Scholar] [CrossRef]
- D’Aquino, L.; Morgana, M.; Carboni, M.A.; Staiano, M.; Antisari, M.V.; Re, M.; Lorito, M.; Vinale, F.; Abadi, K.M.; Woo, S.L. Effect of some rare earth elements on the growth and lanthanide accumulation in different trichoderma strains. Soil Biol. Biochem. 2009, 41, 2406–2413. [Google Scholar] [CrossRef]
- Yuan, Y.; Cave, M.; Zhang, C. Using local moran’s i to identify contamination hotspots of rare earth elements in urban soils of london. Appl. Geochem. 2018, 88, 167–178. [Google Scholar] [CrossRef]
- Chen, Z.; Zhu, X. Accumulation of rare earth elements in bone and its toxicity and potential hazard to health. J. Ecol. Rural Env. 2008, 24, 88–91. [Google Scholar]
- Yuan, Z.; Liu, Y.; Yu, H.; Zheng, H.; Wang, N.; Wu, M.; Liu, Y.; Yan, S. Study on relationship between rare earth level in blood and health condition of residents. Chin. Public Health 2003, 19, 133–135. [Google Scholar]
- Tang, J.; Johannesson, K.H. Controls on the geochemistry of rare earth elements along a groundwater flow path in the carrizo sand aquifer, texas, USA. Chem. Geol. 2006, 225, 156–171. [Google Scholar] [CrossRef]
- Leybourne, M.I.; Johannesson, K.H. Rare earth elements (ree) and yttrium in stream waters, stream sediments, and fe–mn oxyhydroxides: Fractionation, speciation, and controls over ree+ y patterns in the surface environment. Geochim. Et Cosmochim. Acta 2008, 72, 5962–5983. [Google Scholar] [CrossRef]
- Amyot, M.; Clayden, M.G.; MacMillan, G.A.; Perron, T.; Arscott-Gauvin, A. Fate and trophic transfer of rare earth elements in temperate lake food webs. Environ. Sci. Technol. 2017, 51, 6009–6017. [Google Scholar] [CrossRef] [Green Version]
- Migaszewski, Z.M.; Gałuszka, A. The characteristics, occurrence, and geochemical behavior of rare earth elements in the environment: A review. Crit. Rev. Environ. Sci. Technol. 2015, 45, 429–471. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis is central to toxicology, pharmacology and risk assessment. Hum. Exp. Toxicol. 2010, 29, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Pagano, G.; Guida, M.; Tommasi, F.; Oral, R. Health effects and toxicity mechanisms of rare earth elements—Knowledge gaps and research prospects. Ecotoxicol. Environ. Saf. 2015, 115, 40–48. [Google Scholar] [CrossRef]
- Oral, R.; Bustamante, P.; Warnau, M.; d’Ambra, A.; Guida, M.; Pagano, G. Cytogenetic and developmental toxicity of cerium and lanthanum to sea urchin embryos. Chemosphere 2010, 81, 194–198. [Google Scholar] [CrossRef]
- High, W.A.; Ayers, R.A.; Chandler, J.; Zito, G.; Cowper, S.E. Gadolinium is detectable within the tissue of patients with nephrogenic systemic fibrosis. J. Am. Acad. Dermatol. 2007, 56, 21–26. [Google Scholar] [CrossRef]
- Boyd, A.S.; Zic, J.A.; Abraham, J.L. Gadolinium deposition in nephrogenic fibrosing dermopathy. J. Am. Acad. Dermatol. 2007, 56, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.; Bazari, H.; Avery, L.L.; Koreishi, A.F. Case 6-2008: A 46-year-old woman with renal failure and stiffness of the joints and skin. N. Engl. J. Med. 2008, 358, 827–838. [Google Scholar] [CrossRef]
- Sanyal, S.; Marckmann, P.; Scherer, S.; Abraham, J.L. Multiorgan gadolinium (gd) deposition and fibrosis in a patient with nephrogenic systemic fibrosis—An autopsy-based review. Nephrol. Dial. Transplant. 2011, 26, 3616–3626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, E.J.; Schmidt-Lauber, C.; Kay, J. Nephrogenic systemic fibrosis: A systemic fibrosing disease resulting from gadolinium exposure. Best Pract. Res. Clin. Rheumatol. 2012, 26, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.-C.; Wang, H.-Y.; Wang, J.-J.; Kan, W.-C.; Chien, T.-W.; Lin, C.-Y.; Su, S.-B. Risk of acute kidney injury after exposure to gadolinium-based contrast in patients with renal impairment. Ren. Fail. 2011, 33, 758–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ammann, A.A. Speciation of heavy metals in environmental water by ion chromatography coupled to icp–ms. Anal. Bioanal. Chem. 2002, 372, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Gaillardet, J.; Viers, J.; Dupré, B. Trace elements in river waters. TrGeo 2003, 5, 605. [Google Scholar]
- Khan, A.M.; Bakar, N.K.A.; Bakar, A.F.A.; Ashraf, M.A. Chemical speciation and bioavailability of rare earth elements (rees) in the ecosystem: A review. Environ. Sci. Pollut. Res. 2017, 24, 22764–22789. [Google Scholar] [CrossRef] [PubMed]
- Johannesson, K.H.; Tang, J.; Daniels, J.M.; Bounds, W.J.; Burdige, D.J. Rare earth element concentrations and speciation in organic-rich blackwaters of the great dismal swamp, virginia, USA. Chem. Geol. 2004, 209, 271–294. [Google Scholar] [CrossRef]
- Janssen, R.P.; Verweij, W. Geochemistry of some rare earth elements in groundwater, vierlingsbeek, the netherlands. Water Res. 2003, 37, 1320–1350. [Google Scholar] [CrossRef] [Green Version]
- Deberdt, S.; Viers, J.; Dupré, B. New insights about the rare earth elements (ree) mobility in river waters. Bull. De La Société Géologique De Fr. 2002, 173, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Johannesson, K.H.; Stetzenbach, K.J.; Hodge, V.F.; Lyons, W.B. Rare earth element complexation behavior in circumneutral ph groundwaters: Assessing the role of carbonate and phosphate ions. Earth Planet. Sci. Lett. 1996, 139, 305–319. [Google Scholar] [CrossRef]
- Hummel, W.; Glaus, M.; Van Loon, L. Binding of Radionuclides by Humic Substances: The “Conservative Roof” Approach. In Proceedings of the NEA Workshop, Stockholm, Sweden, 25–27 January 1994; pp. 14–16. [Google Scholar]
- Cantrell, K.J.; Byrne, R.H. Rare earth element complexation by carbonate and oxalate ions. Geochim. Cosmochim. Acta 1987, 51, 597–605. [Google Scholar] [CrossRef]
- Wood, S.A. The aqueous geochemistry of the rare-earth elements and yttrium: 1. Review of available low-temperature data for inorganic complexes and the inorganic ree speciation of natural waters. Chem. Geol. 1990, 82, 159–186. [Google Scholar] [CrossRef]
- Lee, J.H.; Byrne, R.H. Complexation of trivalent rare earth elements (ce, eu, gd, tb, yb) by carbonate ions. Geochim. Cosmochim. Acta 1993, 57, 295–302. [Google Scholar]
- Millero, F.J. Stability constants for the formation of rare earth-inorganic complexes as a function of ionic strength. Geochim. Cosmochim. Acta 1992, 56, 3123–3132. [Google Scholar] [CrossRef]
- Åström, M.; Corin, N. Distribution of rare earth elements in anionic, cationic and particulate fractions in boreal humus-rich streams affected by acid sulphate soils. Water Res. 2003, 37, 273–280. [Google Scholar] [CrossRef]
- Pourret, O.; Davranche, M.; Gruau, G.; Dia, A. Rare earth elements complexation with humic acid. Chem. Geol. 2007, 243, 128–141. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M.; Xue, Y.; Kinsela, A.S.; Wilcken, K.M.; Collins, R.N. Donnan membrane speciation of al, fe, trace metals and rees in coastal lowland acid sulfate soil-impacted drainage waters. Sci. Total Environ. 2016, 547, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Usenko, C.Y.; Harper, S.L.; Tanguay, R.L. In vivo evaluation of carbon fullerene toxicity using embryonic zebrafish. Carbon 2007, 45, 1891–1898. [Google Scholar] [CrossRef] [Green Version]
- Asharani, P.; Wu, Y.L.; Gong, Z.; Valiyaveettil, S. Toxicity of silver nanoparticles in zebrafish models. Nanotechnology 2008, 19, 255102. [Google Scholar] [CrossRef]
- Cook, L.W.; Paradise, C.J.; Lom, B. The pesticide malathion reduces survival and growth in developing zebrafish. Environ. Toxicol. Chem. Int. J. 2005, 24, 1745–1750. [Google Scholar] [CrossRef]
- Glaberman, S.; Padilla, S.; Barron, M.G. Evaluating the zebrafish embryo toxicity test for pesticide hazard screening. Environ. Toxicol. Chem. 2017, 36, 1221–1226. [Google Scholar] [CrossRef]
- Jezierska, B.; Ługowska, K.; Witeska, M. The effects of heavy metals on embryonic development of fish (a review). Fish Physiol. Biochem. 2009, 35, 625–640. [Google Scholar] [CrossRef]
- Dave, G.; Xiu, R. Toxicity of mercury, copper, nickel, lead, and cobalt to embryos and larvae of zebrafish, brachydanio rerio. Arch. Environ. Contam. Toxicol. 1991, 21, 126–134. [Google Scholar] [CrossRef]
- Sfakianakis, D.; Renieri, E.; Kentouri, M.; Tsatsakis, A. Effect of heavy metals on fish larvae deformities: A review. Environ. Res. 2015, 137, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Maes, J.; Verlooy, L.; Buenafe, O.E.; De Witte, P.A.; Esguerra, C.V.; Crawford, A.D. Evaluation of 14 organic solvents and carriers for screening applications in zebrafish embryos and larvae. PLoS ONE 2012, 7, e43850. [Google Scholar] [CrossRef] [PubMed]
- Hallare, A.; Nagel, K.; Köhler, H.-R.; Triebskorn, R. Comparative embryotoxicity and proteotoxicity of three carrier solvents to zebrafish (Danio rerio) embryos. Ecotoxicol. Environ. Saf. 2006, 63, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Siregar, P.; Chen, J.-R.; Lai, Y.-H.; Huang, J.-C.; Hsiao, C.-D. Systematical exploration of the common solvent toxicity at whole organism level by behavioral phenomics in adult zebrafish. Environ. Pollut. 2020, 266, 115239. [Google Scholar] [CrossRef]
- Zhang, Z.; Bai, W.; Zhang, L.; He, X.; Ma, Y.; Liu, Y.; Chai, Z. Effects of rare earth elements la and yb on the morphological and functional development of zebrafish embryos. J. Environ. Sci. 2012, 24, 209–213. [Google Scholar]
- Kulaksız, S.; Bau, M. Rare earth elements in the rhine river, germany: First case of anthropogenic lanthanum as a dissolved microcontaminant in the hydrosphere. Environ. Int. 2011, 37, 973–979. [Google Scholar] [CrossRef]
- Kulaksız, S.; Bau, M. Anthropogenic gadolinium as a microcontaminant in tap water used as drinking water in urban areas and megacities. Appl. Geochem. 2011, 26, 1877–1885. [Google Scholar] [CrossRef]
- Anawar, H.; do Freitas, M.; Canha, N.; Dionísio, I.; Dung, H.; Galinha, C.; Pacheco, A. Assessment of bioaccumulation of rees by plant species in a mining area by inaa. J. Radioanal. Nucl. Chem. 2012, 294, 377–381. [Google Scholar] [CrossRef]
- Gonzalez, V.; Vignati, D.A.; Leyval, C.; Giamberini, L. Environmental fate and ecotoxicity of lanthanides: Are they a uniform group beyond chemistry? Environ. Int. 2014, 71, 148–157. [Google Scholar] [CrossRef]
- González, V.; Vignati, D.A.; Pons, M.-N.; Montarges-Pelletier, E.; Bojic, C.; Giamberini, L. Lanthanide ecotoxicity: First attempt to measure environmental risk for aquatic organisms. Environ. Pollut. 2015, 199, 139–147. [Google Scholar] [CrossRef]
- Herrmann, H.; Nolde, J.; Berger, S.; Heise, S. Aquatic ecotoxicity of lanthanum–a review and an attempt to derive water and sediment quality criteria. Ecotoxicol. Environ. Saf. 2016, 124, 213–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moermond, C.T.; Tijink, J.; van Wezel, A.P.; Koelmans, A.A. Distribution, speciation, and bioavailability of lanthanides in the rhine-meuse estuary, the netherlands. Environ. Toxicol. Chem. Int. J. 2001, 20, 1916–1926. [Google Scholar] [CrossRef]
- Sholkovitz, E.R. Chemical evolution of rare earth elements: Fractionation between colloidal and solution phases of filtered river water. Earth Planet. Sci. Lett. 1992, 114, 77–84. [Google Scholar] [CrossRef]
- Das, T.; Sharma, A.; Talukder, G. Effects of lanthanum in cellular systems. Biol. Trace Elem. Res. 1988, 18, 201–228. [Google Scholar] [CrossRef]
- Hanana, H.; Turcotte, P.; Pilote, M.; Auclair, J.; Gagnon, C.; Gagné, F. Biomarker assessment of lanthanum on a freshwater invertebrate, dreissena polymorpha. SOJ Biochem. 2017, 3, 1–9. [Google Scholar] [CrossRef]
- MacMillan, G.A.; Chételat, J.; Heath, J.P.; Mickpegak, R.; Amyot, M. Rare earth elements in freshwater, marine, and terrestrial ecosystems in the eastern canadian arctic. Environ. Sci. Process. Impacts 2017, 19, 1336–1345. [Google Scholar] [CrossRef]
- Drexler, J.; Fisher, N.; Henningsen, G.; Lanno, R.; McGeer, J.; Sappington, K.; Beringer, M. Issue Paper on the Bioavailability and Bioaccumulation of Metals; US Environmental Protection Agency Risk Assessment Forum: Washington, DC, USA, 2003. [Google Scholar]
- Romero-Freire, A.; Joonas, E.; Muna, M.; Cossu-Leguille, C.; Vignati, D.; Giamberini, L. Assessment of the toxic effects of mixtures of three lanthanides (ce, gd, lu) to aquatic biota. Sci. Total Environ. 2019, 661, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Trifuoggi, M.; Pagano, G.; Guida, M.; Palumbo, A.; Siciliano, A.; Gravina, M.; Lyons, D.M.; Burić, P.; Levak, M.; Thomas, P.J. Comparative toxicity of seven rare earth elements in sea urchin early life stages. Environ. Sci. Pollut. Res. 2017, 24, 20803–20810. [Google Scholar] [CrossRef] [Green Version]
- Blaise, C.; Gagné, F.; Harwood, M.; Quinn, B.; Hanana, H. Ecotoxicity responses of the freshwater cnidarian hydra attenuata to 11 rare earth elements. Ecotoxicol. Environ. Saf. 2018, 163, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Blaise, C.; Gagné, F.; Ferard, J.; Eullaffroy, P. Ecotoxicity of selected nano-materials to aquatic organisms. Environ. Toxicol. Int. J. 2008, 23, 591–598. [Google Scholar] [CrossRef]
- Karntanut, W.; Pascoe, D. The toxicity of copper, cadmium and zinc to four different hydra (cnidaria: Hydrozoa). Chemosphere 2002, 47, 1059–1064. [Google Scholar] [CrossRef]
- Holdway, D.A.; Lok, K.; Semaan, M. The acute and chronic toxicity of cadmium and zinc to two hydra species. Environ. Toxicol. 2001, 16, 557–565. [Google Scholar] [CrossRef]
- Quinn, B.; Gagné, F.; Blaise, C. An investigation into the acute and chronic toxicity of eleven pharmaceuticals (and their solvents) found in wastewater effluent on the cnidarian, hydra attenuata. Sci. Total Environ. 2008, 389, 306–314. [Google Scholar] [CrossRef]
- Blinova, I.; Lukjanova, A.; Muna, M.; Vija, H.; Kahru, A. Evaluation of the potential hazard of lanthanides to freshwater microcrustaceans. Sci. Total Environ. 2018, 642, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Tai, P.; Zhao, Q.; Su, D.; Li, P.; Stagnitti, F. Biological toxicity of lanthanide elements on algae. Chemosphere 2010, 80, 1031–1035. [Google Scholar] [CrossRef]
- Joonas, E.; Aruoja, V.; Olli, K.; Syvertsen-Wiig, G.; Vija, H.; Kahru, A. Potency of (doped) rare earth oxide particles and their constituent metals to inhibit algal growth and induce direct toxic effects. Sci. Total Environ. 2017, 593, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Kurvet, I.; Juganson, K.; Vija, H.; Sihtmäe, M.; Blinova, I.; Syvertsen-Wiig, G.; Kahru, A. Toxicity of nine (doped) rare earth metal oxides and respective individual metals to aquatic microorganisms vibrio fischeri and tetrahymena thermophila. Materials 2017, 10, 754. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Xiaorong, W.; Liansheng, W.; Lemei, D.; Zhong, L.; Yijun, C. Bioconcentration of rare earth elements lanthanum, gadolinium and yttrium in algae (chlorella vulgarize beijerinck): Influence of chemical species. Chemosphere 1997, 34, 1753–1760. [Google Scholar] [CrossRef]
- Su, D.; Li, P.; Tai, P. Toxic effects of lanthanides on chlorella autotrophica. Chin. J. Ecol. 2005, 24, 382–384. [Google Scholar]
- Wang, Y.; Zhang, M.; Wang, X. Population growth responses of tetrahymena shanghaiensis in exposure to rare earth elements. Biol. Trace Elem. Res. 2000, 75, 265–275. [Google Scholar] [CrossRef]
- Yang, G.; Tan, Q.G.; Zhu, L.; Wilkinson, K.J. The role of complexation and competition in the biouptake of europium by a unicellular alga. Environ. Toxicol. Chem. 2014, 33, 2609–2615. [Google Scholar] [CrossRef] [PubMed]
- Ishii, N.; Tagami, K.; Uchida, S. Removal of rare earth elements by algal flagellate euglena gracilis. J. Alloys Compd. 2006, 408, 417–420. [Google Scholar] [CrossRef]
- Brown, P.H.; Rathjen, A.H.; Graham, R.D.; Tribe, D.E. Rare earth elements in biological systems. Handb. Phys. Chem. Rare Earths 1990, 13, 423–452. [Google Scholar]
- Wang, K.; Cheng, Y.; Yang, X.; Li, R. Cell responses to lanthanides and potential pharmacological actions of lanthanides. Met. Ions Biol. Syst. 2003, 40, 707. [Google Scholar] [PubMed]
- Hanana, H.; Turcotte, P.; Dubé, M.; Gagnon, C.; Gagné, F. Response of the freshwater mussel, dreissena polymorpha to sub-lethal concentrations of samarium and yttrium after chronic exposure. Ecotoxicol. Environ. Saf. 2018, 165, 662–670. [Google Scholar] [CrossRef]
- Oral, R.; Pagano, G.; Siciliano, A.; Gravina, M.; Palumbo, A.; Castellano, I.; Migliaccio, O.; Thomas, P.J.; Guida, M.; Tommasi, F. Heavy rare earth elements affect early life stages in paracentrotus lividus and arbacia lixula sea urchins. Environ. Res. 2017, 154, 240–246. [Google Scholar] [CrossRef]
- Pagano, G.; Guida, M.; Siciliano, A.; Oral, R.; Koçbaş, F.; Palumbo, A.; Castellano, I.; Migliaccio, O.; Thomas, P.J.; Trifuoggi, M. Comparative toxicities of selected rare earth elements: Sea urchin embryogenesis and fertilization damage with redox and cytogenetic effects. Environ. Res. 2016, 147, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Hua, D.; Wang, J.; Yu, D.; Liu, J. Lanthanum exerts acute toxicity and histopathological changes in gill and liver tissue of rare minnow (Gobiocypris rarus). Ecotoxicology 2017, 26, 1207–1215. [Google Scholar] [CrossRef]
- Figueiredo, C.; Grilo, T.F.; Lopes, C.; Brito, P.; Diniz, M.; Caetano, M.; Rosa, R.; Raimundo, J. Accumulation, elimination and neuro-oxidative damage under lanthanum exposure in glass eels (anguilla anguilla). Chemosphere 2018, 206, 414–423. [Google Scholar] [CrossRef]
- Balusamy, B.; Taştan, B.E.; Ergen, S.F.; Uyar, T.; Tekinay, T. Toxicity of lanthanum oxide (la2o3) nanoparticles in aquatic environments. Environ. Sci. Process. Impacts 2015, 17, 1265–1270. [Google Scholar] [CrossRef]
- Balusamy, B.; Kandhasamy, Y.G.; Senthamizhan, A.; Chandrasekaran, G.; Subramanian, M.S.; Kumaravel, T.S. Characterization and bacterial toxicity of lanthanum oxide bulk and nanoparticles. J. Rare Earths 2012, 30, 1298–1302. [Google Scholar] [CrossRef]
- Peng, L.; Hongyu, X.; Xi, L.; Chaocan, Z.; Yi, L. Study on the toxic mechanism of la 3+ to escherichia coli. Biol. Trace Elem. Res. 2006, 114, 293–299. [Google Scholar] [CrossRef]
- Evseeva, T.; Geras’Kin, S.; Majstrenko, T.; Brown, J.; Belykh, E. Comparative estimation of 232th and stable ce (iii) toxicity and detoxification pathways in freshwater alga chlorella vulgaris. Chemosphere 2010, 81, 1320–1327. [Google Scholar] [CrossRef]
- Paoli, L.; Fiorini, E.; Munzi, S.; Sorbo, S.; Basile, A.; Loppi, S. Uptake and acute toxicity of cerium in the lichen xanthoria parietina. Ecotoxicol. Environ. Saf. 2014, 104, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Hua, Y.; Shen, H.; Zhong, L.; Jin, C.; Mi, Y.; Yao, H.; Xie, Y.; Wei, S.; Zhou, L. Cytochemical behavior of rare earth ions in euglena gracilis studied by xafs. J. Radioanal. Nucl. Chem. 2007, 272, 359–362. [Google Scholar] [CrossRef]
- Rogosnitzky, M.; Branch, S. Gadolinium-based contrast agent toxicity: A review of known and proposed mechanisms. Biometals 2016, 29, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Hanana, H.; Turcotte, P.; André, C.; Gagnon, C.; Gagné, F. Comparative study of the effects of gadolinium chloride and gadolinium—Based magnetic resonance imaging contrast agent on freshwater mussel, dreissena polymorpha. Chemosphere 2017, 181, 197–207. [Google Scholar] [CrossRef]
- Martino, C.; Bonaventura, R.; Byrne, M.; Roccheri, M.; Matranga, V. Effects of exposure to gadolinium on the development of geographically and phylogenetically distant sea urchins species. Mar. Environ. Res. 2017, 128, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Wilkinson, K.J. Biouptake of a rare earth metal (nd) by chlamydomonas reinhardtii–bioavailability of small organic complexes and role of hardness ions. Environ. Pollut. 2018, 243, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Wang, J.; Li, X.; Zhu, J.; Reinert, T.; Heitmann, J.; Spemann, D.; Vogt, J.; Flagmeyer, R.-H.; Butz, T.J.N.I. Study of metal bioaccumulation by nuclear microprobe analysis of algae fossils and living algae cells. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 2000, 161, 801–807. [Google Scholar] [CrossRef]
- Aiube, C.M.; Lobo, T.M.; Sousa-Moura, D.; Ferraz, I.B.M.; Osugi, M.E.; Grisolia, C.K.; Oliveira, R.; Weber, I.T. Study of yvo4 as a photocatalyst: Correlation between synthetic route and ecotoxicity. J. Environ. Chem. Eng. 2018, 6, 2846–2854. [Google Scholar] [CrossRef]
- Yang, W.; Yingjun, W.; Jinge, D.; Zhanghong, W.; Qinglian, W. Effects of yttrium under lead stress on growth and physiological characteristics of microcystis aeruginosa. J. Rare Earths 2016, 34, 747–756. [Google Scholar]
- Wang, Y.; Li, Y.; Luo, X.; Gao, H. Effects of yttrium and phosphorus on growth and physiological characteristics of microcystis aeruginosa. J. Rare Earths 2018, 36, 781–788. [Google Scholar] [CrossRef]
- Xu, T.; Su, C.; Hu, D.; Li, F.; Lu, Q.; Zhang, T.; Xu, Q. Molecular distribution and toxicity assessment of praseodymium by spirodela polyrrhiza. J. Hazard. Mater. 2016, 312, 132–140. [Google Scholar] [CrossRef]
- Rowell, J.A.; Fillion, M.A.; Smith, S.; Wilkinson, K.J. Determination of the speciation and bioavailability of samarium to chlamydomonas reinhardtii in the presence of natural organic matter. Environ. Toxicol. Chem. 2018, 37, 1623–1631. [Google Scholar] [CrossRef]
- Vukov, O.; Smith, D.S.; McGeer, J.C. Acute dysprosium toxicity to daphnia pulex and hyalella azteca and development of the biotic ligand approach. Aquat. Toxicol. 2016, 170, 142–151. [Google Scholar] [CrossRef]
- Fuma, S.; Takeda, H.; Takaku, Y.; Hisamatsu, S.; Kawabata, Z. Effects of dysprosium on the species-defined microbial microcosm. Bull. Environ. Contam. Toxicol. 2005, 74, 263–272. [Google Scholar] [CrossRef]
- Peng, L.; Yi, L.; Xi, L.; Ping, S. Microcalorimetric studies of the biological effect of holmium (iii) on halobacterium halobium r1 growth. Biol. Trace Elem. Res. 2008, 121, 80–86. [Google Scholar] [CrossRef]
- Scholz, S.; Fischer, S.; Gündel, U.; Küster, E.; Luckenbach, T.; Voelker, D. The zebrafish embryo model in environmental risk assessment—Applications beyond acute toxicity testing. Environ. Sci. Pollut. Res. 2008, 15, 394–404. [Google Scholar] [CrossRef]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Sipes, N.S.; Padilla, S.; Knudsen, T.B. Zebrafish—As an integrative model for twenty-first century toxicity testing. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.J.; Jia, Y.F.; Chen, N.; Bian, W.P.; Li, Q.K.; Ma, Y.B.; Chen, Y.L.; Pei, D.S. Zebrafish as a model system to study toxicology. Environ. Toxicol. Chem. 2014, 33, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Nagel, R. Dart: The embryo test with the zebrafish Danio rerio—A general model in ecotoxicology and toxicology. Altex 2002, 19, 38–48. [Google Scholar] [PubMed]
- Teraoka, H.; Dong, W.; Hiraga, T. Zebrafish as a novel experimental model for developmental toxicology. Congenit. Anom. 2003, 43, 123–132. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Industry Usages | Rare Earth Elements (REEs) |
|---|---|
| Cordless power tools | Praseodymium (Pr), Neodymium (Nd), Terbium (Tb), Dysprosium (Dy) |
| Optics | Yttrium (Y), Europium (Eu), Terbium (Tb) |
| Energy-saving light bulbs | Yttrium (Y), Europium (Eu) |
| Liquid-Crystal Display (LCD)/Plasma Display Panel (PDP) screen displays | Yttrium (Y), Cerium (Ce), Europium (Eu), Terbium (Tb) |
| Wind turbines | Praseodymium (Pr), Neodymium (Nd), Dysprosium (Dy) |
| Hybrid vehicles | Praseodymium (Pr), Neodymium (Nd), Samarium (Sm), Terbium (Tb), Dysprosium (Dy) |
| Digital camera lenses, Rechargeable batteries | Lanthanum (La), Cerium (Ce) |
| Speakers, earphones | Praseodymium (Pr), Neodymium (Nd), Gadolinium (Gd) |
| Magnets | Praseodymium (Pr), Neodymium (Nd), Gadolinium (Gd), Terbium (Tb), Dysprosium (Dy) |
| Defense, guidance, and control | Praseodymium (Pr), Neodymium (Nd), Samarium (Sm), Terbium (Tb), Dysprosium (Dy) |
| IPod, CD/DVD, smartphone | Lanthanum (La), Cerium (Ce), Praseodymium (Pr), Neodymium (Nd) |
| Agriculture | Lanthanum (La), Cerium (Ce) |
| Laser, Light-Emitting Diode (LED), fluorescent lamps | Lanthanum (La), Cerium (Ce), Europium (Eu), Terbium (Tb), Yttrium (Y) |
| Health | Praseodymium (Pr), Neodymium (Nd), Gadolinium (Gd), Terbium (Tb), Dysprosium (Dy) |
| Petroleum refining, automotive catalysts, diesel addictive | Lanthanum (La), Cerium (Ce), Praseodymium (Pr), Neodymium (Nd) |
| Colorants, fuel cells, capacitors, sensors, semiconductors | Lanthanum (La), Cerium (Ce), Praseodymium (Pr), Neodymium (Nd), Gadolinium (Gd), Erbium (Er), Holmium (Ho) |
| REEs | Model Organisms | Concentration | Time Duration | Toxicity Analysis | LC50/EC50 | Reference |
|---|---|---|---|---|---|---|
| Ce, Gd, and Lu | A. fischeri, R. subcapitata, C. vulgaris, B. calyciflorus, H. incongruens, Daphnia magna, and Danio rerio | CE1, CE2, CE3, Gd1, Gd2, Gd3, Lu1, Lu2, Lu3, Dual Salt mixture (A, B, C) 12 Treatments in different Concentrations (μg/L) | MIT 0.5 h LUT 4 h ALT 72 h ROT 48 h OST 144 h DAT 48 h FET 96 h | Calculated from t = oh to t = xh different results, according to concentration and time duration | NA | [74] |
| CeCl3·7H2O, GdCl3·6H2O, LuCl3·6H2O | A. fischeri, Pseudokirchneriella subcapitata, D. magna, H. incongruens, B. calyciflorus, and Hydra attenuate | 100, 200, 400, 800, 1600, 3200, and 6400 mg/L | 48, 72, 96 h and 6 days | Considering the toxicity of the three lanthanides studied, crustaceans were the least sensitive species and rotifer, and cnidarians were the most sensitive ones | LC50 values for D. magna and H. incongruens organisms, and EC50 for growth in H. incongruens, are anticipated to be greater than 6400 mg/L | [66] |
| Yttrium (Y), La, Ce, Nd, Sm, Eu, Gd | Two Sea urchin species of Sphaerechinus granularis and Arbacia lixula | 10−7 to 10−4 M | 72 hpf, 5 hpf, | Embryotoxicity and offspring damage | NA | [75] |
| Y, La, Ce, Pr, Nd, Sm, Gd, Tb, Dy, Er, Lu | Freshwater cnidarian Hydra attenuata | 1 g/L stock solution—in Hydra medium, 10 mg/L for subsequent bioanalysis | 96 h | Hydra morphology based on the Welby scale | y-0.22, La-0.21, Ce-0.33, Pr-0.56, Nd-0.31, Sm-0.77, Gd-0.52, Tb-0.70, Dy-0.69, Er-0.40, Lu-0.29 mg/L | [76] |
| La, Ce, Pr, Nd, and Gd | Thamnocephalus platyurus, Daphnia magna, and Heterocypris incongruens | 0.01, 0.1, 0.25, 0.5 and 1.0 mg Ln/L | 21 days reproduction tests | Lanthanides were very toxic to crustaceans showing potential hazards for aquatic ecosystems. | 0.3–0.5 mg Ln/L | [81] |
| LaCl3, Ce(N)3)3, NdCl3, SmCl3, Eu(NO3)3, Gd(NO3)3, TbCl3, DyCl3, HoCl3, Er(NO3)3, TmCl3, YbCl3, LuCl3, ScCl3, Y(NO3)3 | Skeletonema costatum | 5, 10, 20, 30, and 40 lmol L−1 Yttrium were 20, 30, 40, 50, and 60 lmol L−1 | 72 h | There is no difference in the LLG and HLG on inhibition of algae cells, | 96-EC50 of La, Y and Sc on S. costatum was 29.19 lmol L−1, 43.21 lmol L−1 and 21.88 lmol L−1 | [82] |
| CGO, LFO, GCO, LCO, LSM, CEO, CPO, LSC, LNO | Raphidocelis subcapitata | 1–100 mg/L | 72 h | Two main mechanisms of inhibition by REEs: Nutrient removal from the algal medium Agglomeration of particles around algal cells | EC50 1 to 98 mg/L | [83] |
| CGO, LFO, GCO, LCO, LSM, CEO, CPO, LSC, LNO | Vibrio fischeri and Tetrahymena thermophila | 1, 10 and 100 mg/L | 24 h | Most REOs produced reactive oxygen species; all studied soluble REEs were toxic to bacteria. Dopant metals (Ni2+, Fe3+) proved toxic, no toxicity of REOs to protozoa and bacteria was observed except for La2NiO4 | Bacteria EC50 3.5–21 mg metal/L Protozoa EC50 28–42 mg/L | [84] |
| La, Gd, and Y with ligands EDTA, NTA, and Cit. | Chlorella Vulgarize Beijerinck | 1.0 mg/L | 96 h | Adding organic ligands that can form the RE-Organic species complex led to a great reduction of the REEs’ bio-concentration in algae. | NA | [85] |
| La, Sm, Y, and Gd | Tetrahymena shanghaiensis | 0.125, 0.25, 0.5, 1.0, 2.0, and 4.0 mM | 24 and 96 h | 24- and 96-h assays suggest a dual effect of REEs on T. shanghaiensis of the stimulated growth at low concentrations and the toxicity at higher concentrations | NA | [87] |
| Eu and Sm | Chlamydomonas reinhardtii | 1×10−9–1×10−5 M | 60 min | REE was likely to share a common bio uptake pathway; the bio uptake of a one REE was reduced when another was present, and REE complexes were bioavailable. | NA | [88] |
| Sm and Y | Dreissena polymorpha (freshwater Mussel) | 10, 50, 250 and 1250 µg/L | 28 days | Y more toxic than Sm, (further toxicity analysis required). | NA | [92] |
| La, Yb | Danio rerio embryos | 0, 0.01, 0.1, 0.3, 0.5 and 1.0 mM | Time points—8, 24, 32, 48–60, 72, and 96 h | Yb more toxic than La (Some other Malformations Indicated Dose dependency) | Median LC50 La3+ and Yb3+ were 0.603 and 0.268 mmol/L | [61] |
| Scandium (Sc), Y, La, Ce, Pr, Nd, Sm, Eu, Gd, Tb, Dy, Ho, Er, Tm, Yb, and Lu | Euglena gracilis | 10 g/L | 7 days | Removal of REEs by E. gracilis from acidic solution was demonstrated. This ability of E. gracilis suggests a potential use in wastewater remediation. | NA | [89] |
| Dy, Ho, Er, Yb and Lu, Ce(III) | Paracentrotus lividus and Arbacia lixula (Embryo and Sperm) | Embryos exposure (10−7 to 10−5 M), 50-µL sperm pellet10−5 to 10−4 M | Embryos—10 min—72 hpf. Sperm—1 h in 30 mL FSW Offspring, residual HREE concentrations of 5 × 10−8 and 5 × 10−7 M | Different toxicity of different HREEs in concentration 10−6 M for embryos, to 10−5–10−4 M for sperm | Comparable effects of REEs according to Different endpoints in the paper. | [93] |
| Y, La, Ce, Nd, Sm, Eu and Gd | Sea urchin (Paracentrotus lividus) (embryo and sperm) | 108 to 104 M | 10 min—72 hpf | Nd (III) and Sm (III) resulted in relatively lesser toxicity in the tested endpoints | NA | [94] |
| CeCl3–7H2O GdCl3−6H2O LuCl3-6H2O | Aliivibrio fischeri Pseudokirchneriella subcapitata Daphnia magna Heterocypris incongruens, Brachionus calyciflorus Hydra attenuata | 100, 200, 400, 800, 1600, 3200, and 6400 μg/L | Different Time periods for all different test | Ecotoxicity increased with an increase in atomic number in bacteria and algae No general pattern established | NA | [66] |
| REE | Model Organisms | Concentration | Time Duration | Toxicity Analysis | LC50/EC50 | Ref. |
|---|---|---|---|---|---|---|
| Lanthanum (La (III)) | Rare minnow (Gobiocypris rarus) | (0.0, 0.1, 0.5, 1.0, 5.0, 10.0, and 40.0 mg/L) | 96 h | Severe acute Toxic effects of La (III) on organisms after 21 d of exposure, abnormal dose-dependent behavior | LC50 of La (III)at 96 h (1.92 mg/L) | [95] |
| LaCl3 | Anguilla Anguilla | 120 ng/L | 14 days | A significant increase in AChE activity, depression in lipid peroxidation and CAT inhibition | NA | [96] |
| La2O3 NP | Chlorella sp. and Daphnia magna | 10, 50, 100, 250, 500, and 1000 mg/L | 2, 24, 72 h | Absence of toxic effects of La2O3 NP on Chlorella sp., even at higher concentrations (1000 mg/L) after 72 h exposure. No significant toxic effects were observed on D. magna at conc. of 250 mg/L or less, and considerable toxic effects were noted in higher concentrations | EC50-500 mg L−1; lethal dose LD50 1000 mg/L | [97] |
| La2O3 | S. aureus, E. coli, and P. aeruginosa | 100 µL | 24 h | Antimicrobial activity against Staphylococcus aureus, but not against Escherichia coli and Pseudomonas aeruginosa | NA | [98] |
| La | Daphnia carinata | 100, 200, 400, 600, 800, and 1000 μg La/L. In Tap Water, Daphnia Water, Hard water (ASTM) | 24–48 h | Acute and chronic toxicity | NA | [99] |
| Thorium (232Th) and stable chemical analogue Ce | Chlorella vulgaris | 0.001–28.013 µM 232Th and 0.036–71.367 µM Ce | 24 h | 232Th was more toxic to Ch. vulgaris after a 24-h exposure than its nonradioactive chemical analogue Ce. Detoxification pathways of Ce (III) in Ch. vulgaris differ from those for 232Th | NA | [100] |
| Ce | Xanthoria parietina | 0.1 mM, 1 mM, 10 mM, and 100 mM | 24 h | Ce bioaccumulation, both extracellularly and intracellularly, caused acute toxicity, evident as decreased sample viability, decrease in photosynthetic performance, and changes in the ultrastructure of the lichen X. parietina | NA | [101] |
| CeCl3, Ce(SO4)2 | Euglena gracilis | 3.3 µg/mL | 2 h | Intracellular lanthanide was able to pass through the chloroplast’s internal membrane system until the replacement of Mg in chlorophyll molecules | NA | [102] |
| GdCl3, gadolinium-based MRI contrast agent (Omniscan) | D. polymorpha | 10, 50, 250, and 1250 mg/L | 28 days | Gd accumulated in zebra tissue after GdCl3 exposure was highly correlated to SOD, CAT, GST, and CO1 gene expressions and also to COX. The major factors affected by Omniscan were GST gene expression, SOD, CO1 and LPO | NA | [71] |
| Gd | Paracentrotus lividus and Arbacia lixula Heliocidaris tuberculata and Centrostephanus rodgersii | 1 to 125 µM | 48 hpf | All four species different had different sensitivity to Gd, but the effect of this agent on larval phenotype was similar | EC50 56 nM to 132 µM | [105] |
| Nd | Chlamydomonas reinhardtii | 1 × 10−9 to 1 × 10−5 M | 1, 15, 30, 45 and 60 min | A Michaelis–Menten equation described Nd bio uptake with an affinity constant, KNd, of 106.8 M−1 and a maximum internalization flux of Jmax = 1.70 × 10−14 mol cm−2 s−1 | NA | [106] |
| Nd3+ | Euglena gracilis | 50 mg/L | 30 min | The bioaccumulation efficiency of the rare earth metal neodymium by the green algae species E. gracilis can be ten times higher if the algae activity is stopped by immobilization | NA | [107] |
| YVO4 | D. rerio embryos | 18.75 mg | 6 days | Inhibition of egg hatching and mortality of D. rerio embryos after hatching | NA | [108] |
| Y3+ | Microcystis aeruginosa | Y3+ 0.00, 0.10, 0.20, 0.50, 1.00, 2.00, 5.00, 10.00 mg/L, and (1.00 mg/L of Pb2+ constant) | 16 days | 0.10–0.50 mg/L Y3+ could stimulate the growth of M. aeruginosa under lead stress. Low concentration of Y3+ would induce less ROS generation and oxidative stress under Pb stress evident from activities of SOD, POD, CAT, and MDA | NA | [109] |
| Y | Microcystis aeruginosa | Yttrium 0.0, 0.1, 0.3, 0.5, 1.0 mg/L, and 0.02, 0.2 mg/L phosphorus | 20 days | 0.1 and 0.3 mg/L yttrium doses promote the growth of M. aeruginosa, while the growth will be inhibited by 0.5 and 1.0 mg/L yttrium. The effects, which are caused by yttrium are different when algae are exposed to different doses of phosphorus | NA | [110] |
| Pr | Spirodela polyrhiza | 0 to 60 μM | 20 days | Significant increases in malondialdehyde (MDA), and decreases in photosynthetic pigment, soluble protein, and unsaturated fatty acids showed that Pr induced oxidative stress. Inhibitory effects on photosystem II and the degradation of the reaction center proteins D1 and D2 were revealed by chlorophyll a fluorescence transients, immunoblotting, and damage to chloroplast ultrastructure | NA | [111] |
| Sm3+ | Green alga Chlamydomonas reinhardtii | 10−9 and 10−5 M | 1, 5, 10, and 15 min | Toxicity is likely to be dependent on free ion concentration | NA | [112] |
| Dy | Daphnia pulex and Hyalella azteca | 500, 500, 125, 25, and 5 M | Hyalella (96 h) Daphnia (48 h) | Hyalella to be 1.4 times more sensitive than Daphnia. The addition of Ca/Na protects Hyalella against Dy toxicity. Low pH was associated with a reduction in toxicity | Acute Dy exposure to D. pulex LC50 = 3.0 µM), H. Azteca LC50 = 2.1 µM) | [113] |
| Dy | Aquatic microbial microcosm consisting of flagellate algae—Euglena Gracilis, Ciliate protozoa- Tetrahymena thermophila, Bacteria—E.coli | 50, 100, 180, 300, 560, 1000 µM | 10 days–120 days, in accordance with the microbial microcosm | Toxicity of Dy was mitigated in microcosm compared to pure culture system for all different species of microcosm | NA | [114] |
| Ho3+ | Halobacterium halobium | 10, 20, 40, 60, 80, and 100 μg/mL | 13,000 min | Values of Pm and k are linked to the concentration of Ho3+, Ho3+ causes a decrease of the maximum heat production and growth rate constants | NA | [115] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malhotra, N.; Hsu, H.-S.; Liang, S.-T.; Roldan, M.J.M.; Lee, J.-S.; Ger, T.-R.; Hsiao, C.-D. An Updated Review of Toxicity Effect of the Rare Earth Elements (REEs) on Aquatic Organisms. Animals 2020, 10, 1663. https://doi.org/10.3390/ani10091663
Malhotra N, Hsu H-S, Liang S-T, Roldan MJM, Lee J-S, Ger T-R, Hsiao C-D. An Updated Review of Toxicity Effect of the Rare Earth Elements (REEs) on Aquatic Organisms. Animals. 2020; 10(9):1663. https://doi.org/10.3390/ani10091663
Chicago/Turabian StyleMalhotra, Nemi, Hua-Shu Hsu, Sung-Tzu Liang, Marri Jmelou M. Roldan, Jiann-Shing Lee, Tzong-Rong Ger, and Chung-Der Hsiao. 2020. "An Updated Review of Toxicity Effect of the Rare Earth Elements (REEs) on Aquatic Organisms" Animals 10, no. 9: 1663. https://doi.org/10.3390/ani10091663