Seasonal Expression of NGF and Its Cognate Receptors in the Ovaries of Grey Squirrels (Sciurus carolinensis)

 , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Capture and Sample Collection
2.2. Immunohistochemistry
2.3. RNA Extraction and Real-Time PCR
2.4. NGF Plasma Levels
2.5. Statistical Analysis
3. Results
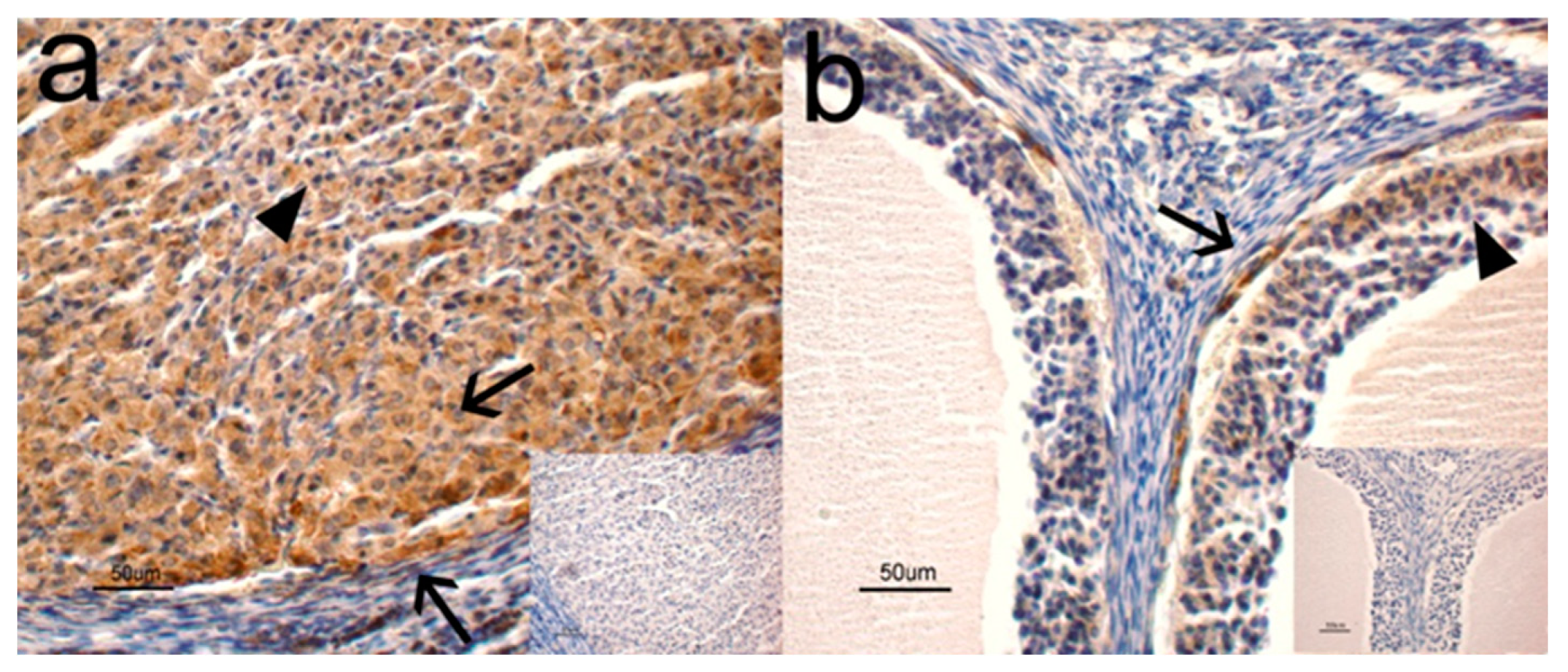
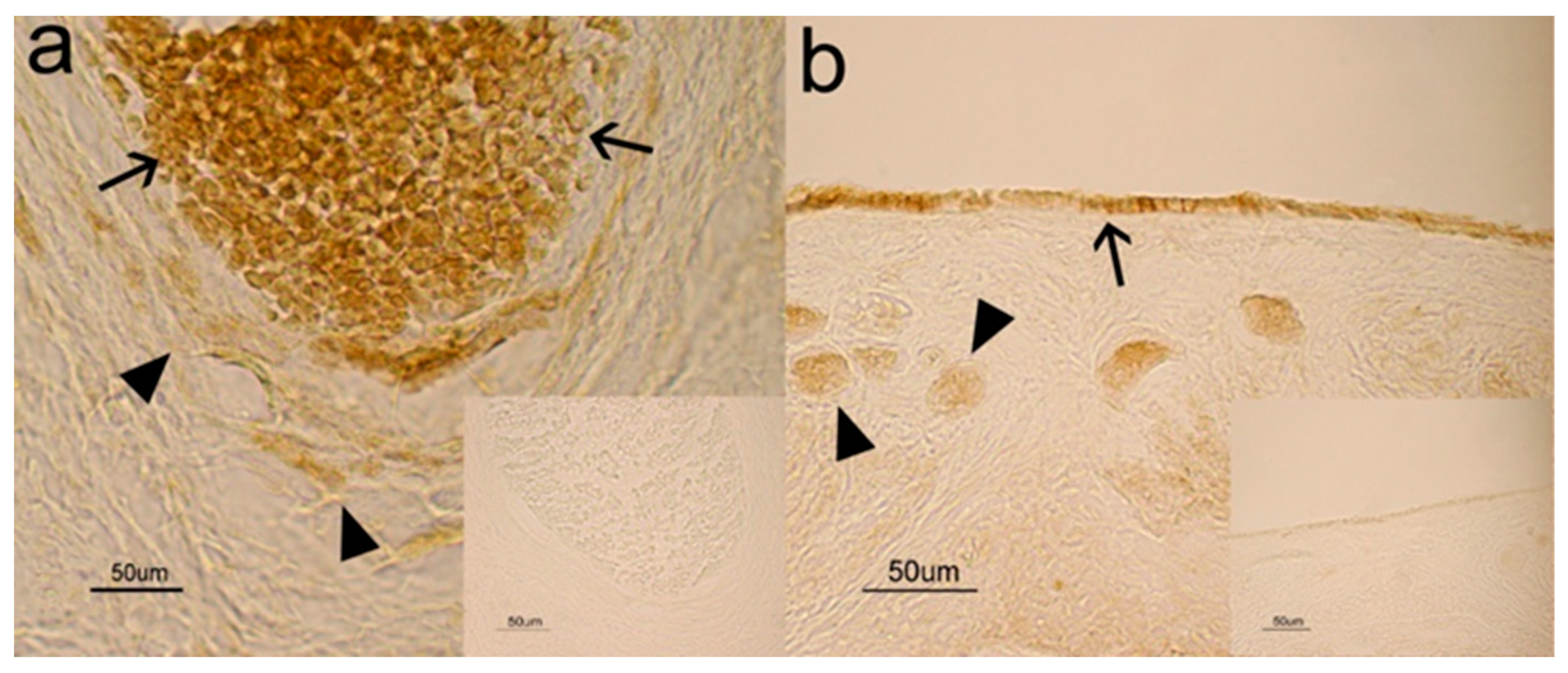
3.1. Immunohistochemistry
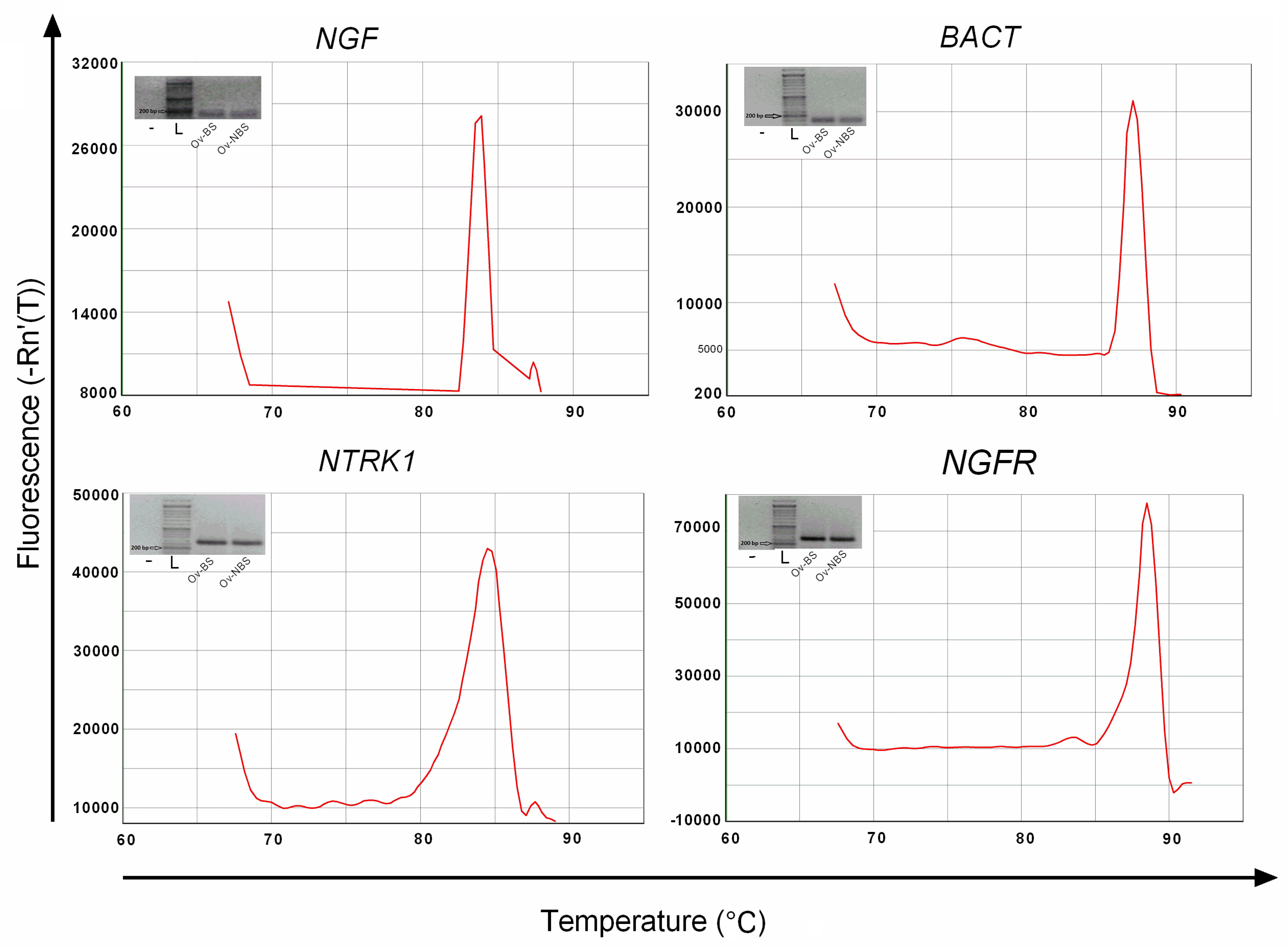
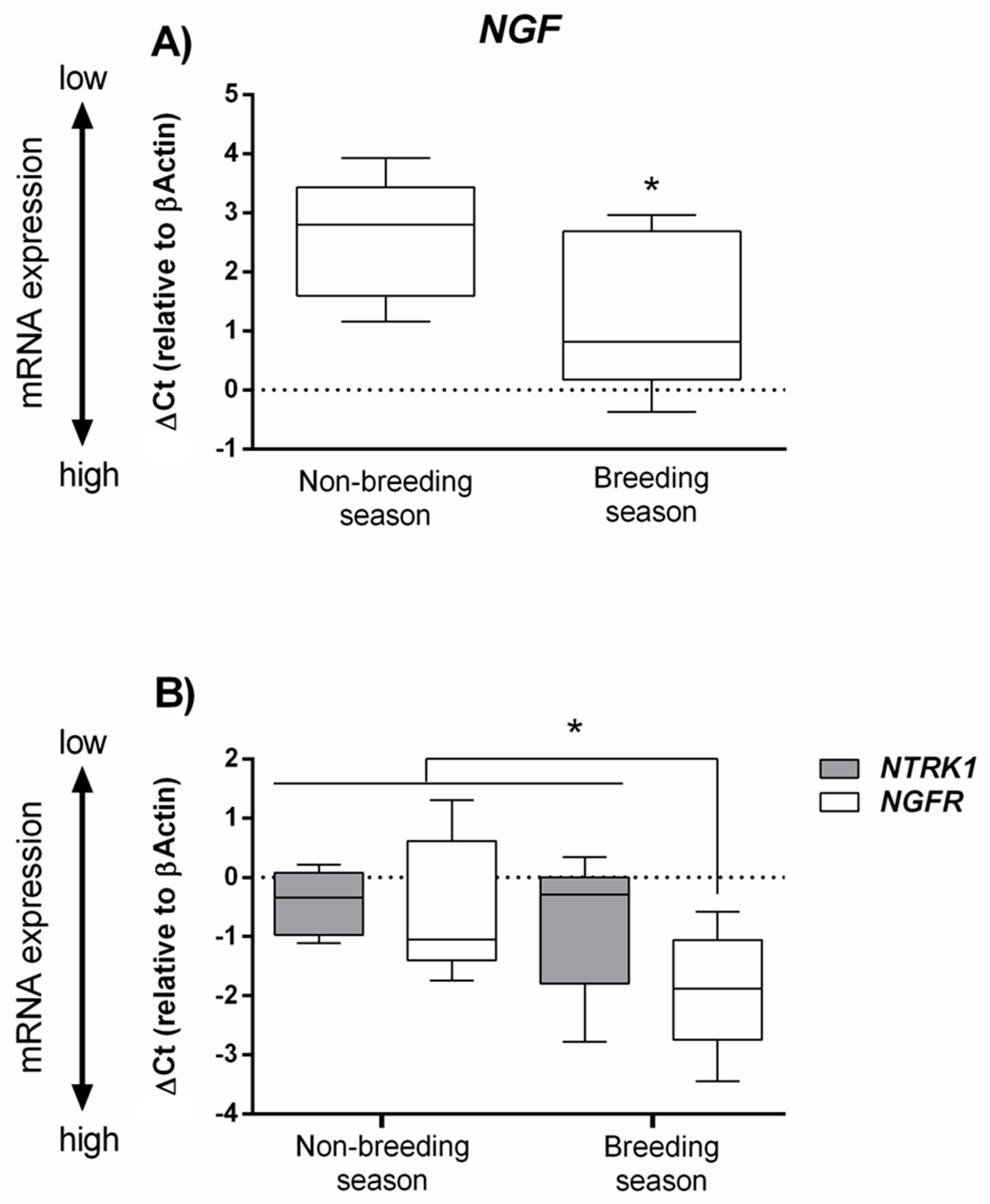
3.2. Real-Time PCR
3.3. NGF Plasma Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Levi-Montalcini, R. The nerve growth factor 35 years later. Science 1987, 237, 1154–1162. [Google Scholar] [CrossRef]
- Schecterson, L.C.; Bothwell, M. Neurotrophin receptors: Old friends with new partners. Dev. Neurobiol. 2010, 70, 332–338. [Google Scholar] [CrossRef]
- Thoenen, H.; Barde, Y.A. Physiology of nerve growth factor. Physiol. Rev. 1980, 60, 1284–1335. [Google Scholar] [CrossRef]
- Snider, W.D. Functions of the neurotrophins during nervous system development: What the knockouts are teaching us. Cell 1994, 77, 627–638. [Google Scholar] [CrossRef]
- Bogle, O.A.; Carrasco, R.A.; Ratto, M.H.; Singh, J.; Adams, G.P. Source and localization of ovulation-inducing factor/nerve growth factor in male reproductive tissues among mammalian species. Biol. Reprod. 2018, 99, 1194–1204. [Google Scholar] [CrossRef]
- Ratto, M.H.; Berland, M.; Silva, M.E.; Adams, G.P. New insights of the role of β-NGF in the ovulation mechanism of induced ovulating species. Reproduction 2019, 157, R199–R207. [Google Scholar] [CrossRef] [Green Version]
- Bjorling, D.E.; Beckman, M.; Clayton, M.K.; Wang, Z.Y. Modulation of nerve growth factor in peripheral organs by estrogen and progesterone. Neuroscience 2002, 110, 155–167. [Google Scholar] [CrossRef]
- Brauer, M.M.; Shockley, K.P.; Chávez, R.; Richeri, A.; Cowen, T.; Crutcher, K.A. The role of NGF in pregnancy-induced degeneration and regeneration of sympathetic nerves in the guinea pig uterus. J. Auton. Nerv. Syst. 2000, 79, 19–27. [Google Scholar] [CrossRef]
- Wessels, J.M.; Wu, L.; Leyland, N.A.; Wang, H.; Foster, W.G. The brain uterus connection: Brain derived neurotrophic factor (BDNF) and its receptor (Ntrk2) are conserved in the mammalian uterus. PLoS ONE 2014, 9, e94036. [Google Scholar] [CrossRef] [Green Version]
- Mirshokraei, P.; Hassanpour, H.; Rahnama, A.; Foster, W.G. Gene expression of BDNF and its receptors, TrkB and p75 in the uterus and oviduct of pregnant and non-pregnant ewes. Res. Vet. Sci. 2013, 95, 164–168. [Google Scholar] [CrossRef]
- Lobos, E.; Gebhardt, C.; Kluge, A.; Spanel-Borowski, K. Expression of nerve growth factor (NGF) isoforms in the rat uterus during pregnancy: Accumulation of precursor proNGF. Endocrinology 2005, 146, 1922–1929. [Google Scholar] [CrossRef] [Green Version]
- Maranesi, M.; Parillo, F.; Leonardi, L.; Rebollar, P.G.; Alonso, B.; Petrucci, L.; Gobbetti, A.; Boiti, C.; Arruda-Alencar, J.; Moura, A.; et al. Expression of nerve growth factor and its receptors in the uterus of rabbits: Functional involvement in prostaglandin synthesis. Domest. Anim. Endocrinol. 2016, 56, 20–28. [Google Scholar] [CrossRef]
- Dissen, G.A.; Romero, C.; Hirshfield, A.N.; Ojeda, S.R. Nerve growth factor is required for early follicular development in the mammalian ovary. Endocrinology 2001, 142, 2078–2086. [Google Scholar] [CrossRef]
- Dissen, G.A.; Hirshfield, A.N.; Malamed, S.; Ojeda, S.R. Expression of neurotrophins and their receptors in the mammalian ovary is developmentally regulated: Changes at the time of folliculogensis. Endocrinology 1995, 136, 4681–4692. [Google Scholar] [CrossRef]
- Weng, Q.; Shi, Z.; Tukada, J.; Wantanabe, G.; Taya, K. Immunodetection of NGF, trkA, p75 and inhibin alpha-subunit in interstitial cells of golden hamsters treated with hCG. J. Reprod. Dev. 2009, 55, 622–628. [Google Scholar] [CrossRef] [Green Version]
- Mattioli, M.; Barboni, B.; Gioia, L.; Lucidi, P. Nerve growth factor production in sheep antral follicles. Domest. Anim. Endocrinol. 1999, 17, 361–371. [Google Scholar] [CrossRef]
- Ren, L.; Medan, M.S.; Weng, Q.; Jin, W.; Li, C.; Watanabe, G.; Taya, K. Immunolocalization of nerve growth factor (NGF) and its receptors (TrkA and p75LNGFR) in the reproductive organs of shiba goats. J. Reprod. Dev. 2005, 51, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Tanaka, A.; Watanabe, G.; Matsuda, H.; Taya, K. Effect of NGF on the motility and acrosome reaction of golden hamster spermatozoa in vitro. J. Reprod. Dev. 2010, 56, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Dissen, G.A.; Parrott, J.A.; Skinner, M.K.; Hill, D.F.; Costa, M.E.; Ojeda, S.R. Direct effects of nerve growth factor on thecal cells from antral ovarian follicles. Endocrinology 2000, 141, 4736–4750. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, R.; Singh, J.; Adams, G.P. The dynamics of trkA expression in the bovine ovary are associated with a luteotrophic effect of ovulation-inducingfactor/nerve growth factor (OIF/NGF). Reprod. Biol. Endocrinol. 2016, 14, 47. [Google Scholar] [CrossRef] [Green Version]
- Maranesi, M.; Petrucci, L.; Leonardi, L.; Piro, F.; Rebollar, P.G.; Millán, P.; Cocci, P.; Vullo, C.; Parillo, F.; Moura, A.; et al. New insights on a NGF-mediated pathway to induce ovulation in rabbits (Oryctolaguscuniculus). Biol. Reprod. 2018, 98, 634–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Sheng, X.; Song, M.; Zhang, H.; Weng, J.; Zhang, M.; Hu, X.; Zhou, J.; Xu, M.; Weng, Q.; et al. Expression of nerve growth factor and its receptors TrkA and p75 in the uterus of wild female ground squirrel,(Citellusdauricus Brandt). Gen. Comp. Endocrinol. 2012, 176, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Li, Q.; Liu, Y.; Li, B.; Sheng, X.; Han, Y.; Weng, Q. Immunolocalization of NGF and its receptors in ovarian surface epithelium of the wild ground squirrel during the breeding and nonbreeding seasons. Eur. J. Histochem. 2014, 58, 2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koprowski, J.L. Sciurus carolinensis. Mamm. Species 1994, 480, 1–9. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species a Selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG) a Specialist Group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN): Auckland, New Zealand, 2000; pp. 1–12. [Google Scholar]
- Gurnell, J.; Pepper, H. A critical look at conserving the British red squirrel Sciurus vulgaris. Mammal Rev. 1993, 23, 125–136. [Google Scholar] [CrossRef]
- Lawton, C.; Cowan, P.; Bertolino, S.; Lurz, P.W.W.; Peters, A.R. Consequences of introduction of non-indigenous species: Two case studies, the grey squirrel in Europe and the brushtail possum in New Zealand. Rev. Sci. Tech. 2010, 29, 113–122. [Google Scholar]
- Martinoli, A.; Bertolino, S.; Preatoni, D.G.; Balduzzi, A.; Marsan, A.; Genovesi, P.; Tosi, G.; Wauters, L.A. Headcount 2010: The multiplication of the grey squirrel introduced in Italy. Hystrix 2010, 21, 127–136. [Google Scholar]
- Maranesi, M.; Bufalari, A.; Dall’Aglio, C.; Paoloni, D.; Moretti, G.; Crotti, S.; Manuali, E.; Stazi, M.; Bergamasco, F.; Cruciani, D.; et al. Reproductive Traits of an Invasive Alien Population of Grey Squirrel (Sciurus carolinensis) in Central Italy. Animals 2020, 10, 738. [Google Scholar] [CrossRef]
- Cowles, C.J.; Kirkpatrick, R.L.; Newell, J.O. Ovarian follicular changes in gray squirrels as affected by season, age, and reproductive state. J. Mammal. 1977, 58, 67–73. [Google Scholar] [CrossRef]
- Signorile, A.L.; Paoloni, D.; Reuman, D.C. Grey squirrels in central Italy: A new threat for endemic red squirrel subspecies. Biol. Invasions 2014, 16, 2339–2350. [Google Scholar] [CrossRef] [Green Version]
- Santicchia, F.; Romeo, C.; Grilli, G.; Vezzoso, S.; Wauters, L.A.; Mazzamuto, M.V.; Martinoli, A.; Ferrari, N. The use of uterine scars to explore fecundity levels in invasive alien tree squirrels. Hystrix 2015, 26, 95–101. [Google Scholar]
- La Morgia, V.; Paoloni, D.; Genovesi, P. Eradicating the grey squirrel Sciurus carolinensis from urban areas: An innovative decision-making approach based on lessons learnt in Italy. Pest Manag. Sci. 2017, 73, 354–363. [Google Scholar] [CrossRef]
- Bufalari, A.; Maranesi, M.; Marenzoni, M.L.; Mercati, F.; Nannarone, S.; Dall’Aglio, C.; Bellocchi, F.; Zerani, M.; Petrucci, L.; Paoloni, D. Traditional gonadectomy vs gonadectomy by Ligasure® in grey squirrels (Sciurus carolinensis): Comparing surgical techniques. Reprod. Domest. Anim. 2019, 54 (Suppl. S3), 137. [Google Scholar]
- Parillo, F.; Dall’Aglio, C.; Brecchia, G.; Maranesi, M.; Polisca, A.; Boiti, C.; Zerani, M. Aglepristone (RU534) effects on luteal function of pseudopregnant rabbits: Steroid receptors, enzymatic activities, and hormone productions in corpus luteum and uterus. Anim. Reprod. Sci. 2013, 138, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Troisi, A.; Dall’Aglio, C.; Maranesi, M.; Orlandi, R.; Speranza, R.; Zerani, M.; Polisca, A. Presence and expression of apelin and apelin receptor in bitch placenta. Theriogenology 2020, 147, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Dall’Aglio, C.; Mercati, F.; Maranesi, M.; Boiti, C. Identification of orexins and cognate receptors in the lacrimal gland of sheep. Peptides 2012, 35, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Parillo, F.; Catone, G.; Maranesi, M.; Gobbetti, A.; Gasparrini, B.; Russo, M.; Boiti, C.; Zerani, M. Immunolocalization, gene expression, and enzymatic activity of cyclooxygenases, prostaglandin e2-9-ketoreductase, and nitric oxide synthases in Mediterranean buffalo (Bubalusbubalis) corpora lutea during diestrus. Microsc. Res. Tech. 2012, 75, 1682–1690. [Google Scholar] [CrossRef] [PubMed]
- Livak, M.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mercati, F.; Maranesi, M.; Dall’Aglio, C.; Petrucci, L.; Pasquariello, R.; Tardella, F.M.; De Felice, E.; Scocco, P. Apelin System in Mammary Gland of Sheep Reared in Semi-Natural Pastures of the Central Apennines. Animals 2018, 8, 223. [Google Scholar] [CrossRef] [Green Version]
- Maranesi, M.; Zerani, M.; Leonardi, L.; Pistilli, A.; Arruda-Alencar, J.; Stabile, A.M.; Rende, M.; Castellini, C.; Petrucci, L.; Parillo, F.; et al. Gene Expression and Localization of NGF and Its Cognate Receptors NTRK1 and NGFR in the Sex Organs of Male Rabbits. Reprod. Domest. Anim. 2015, 50, 918–925. [Google Scholar] [CrossRef]
- Chaves, R.N.; Alves, A.M.; Lima, L.F.; Matos, H.M.; Rodrigues, A.P.; Figueiredo, J.R. Role of nerve growth factor (NGF) and its receptors in folliculogenesis. Zygote 2013, 21, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Streiter, S.; Fisch, B.; Sabbah, B.; Ao, A.; Abir, R. The importance of neuronalgrowth factors in the ovary. Mol. Hum. Reprod. 2016, 22, 3–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.M.; Wu, H.C.; Sun, Z.G.; Lian, F.; Leung, P.C.K. Neurotrophins and glial cell line-derived neurotrophic factor in the ovary: Physiological andpathophysiological implications. Hum. Reprod. Updat. 2019, 25, 224–242. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Güell, M.; Serrano, L. Correlation of mRNA and protein in complex biological samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohn, J.; Aloyz, R.S.; Toma, J.G.; Haak-Frendscho, M.; Miller, F.D. Functionally antagonistic interactions between the TrkA and p75 neurotrophin receptors regulate sympathetic neuron growth and target innervation. J. Neurosci. 1999, 19, 5393–5408. [Google Scholar] [CrossRef] [Green Version]
- Valderrama, X.P.; Goicochea, J.F.; Silva, M.E.; Ratto, M.H. The effect of seminal plasma β-NGF on follicular fluid hormone concentration and gene expression ofsteroidogenic enzymes in llama granulosa cells. Reprod. Biol. Endocrinol. 2019, 17, 60. [Google Scholar] [CrossRef]
- Li, B.; Sheng, X.; Bao, L.; Huang, S.; Li, Q.; Liu, Y.; Han, Y.; Watanabe, G.; Taya, K.; Weng, Q. Seasonal changes in expression of nerve growth factor and its receptors TrkA and p75 in the ovary of wild ground squirrel (Citellusdauricus Brandt). J. Ovarian Res. 2014, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.P.; Ratto, M.H.; Huanca, W.; Singh, J. Ovulation-inducing factor in the seminal plasma of alpacas and llamas. Biol. Reprod. 2005, 73, 452–457. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Ulloa-Leal, C.; Valderrama, X.P.; Bogle, O.A.; Adams, G.P.; Ratto, M.H. Nerve growth factor from seminal plasma origin (spβ-NGF) increases CL vascularization and level of mRNA expression of steroidogenic enzymes during the early stage of Corpus Luteum development in llamas. Theriogenology 2017, 103, 69–75. [Google Scholar] [CrossRef]
- Tribulo, P.; Bogle, O.; Mapletoft, R.J.; Adams, G.P. Bioactivity of ovulation inducing factor (or nerve growth factor) in bovine seminal plasma and its effects onovarian function in cattle. Theriogenology 2015, 83, 1394–1401. [Google Scholar] [CrossRef]
- Stewart, J.L.; Mercadante, V.R.G.; Dias, N.W.; Canisso, I.F.; Yau, P.; Imai, B.; Lima, F.S. Nerve Growth Factor-Beta, purified from bull seminal plasma, enhances corpus luteum formation and conceptus development in Bostaurus cows. Theriogenology 2018, 106, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Ulloa-Leal, C.; Norambuena, C.; Fernández, A.; Adams, G.P.; Ratto, M.H. Ovulation-inducing factor (OIF/NGF) from seminal plasma origin enhances Corpus Luteum function in llamas regardless the preovulatory follicle diameter. Anim. Reprod. Sci. 2014, 148, 221–227. [Google Scholar] [CrossRef]
- Ulloa-Leal, C.E.; Bogle, O.A.; Adams, G.P.; Ratto, M.H. Luteotrophic effect of ovulation-inducing factor/nerve growth factor (OIF/NGF) present in the seminal plasma of llamas. Theriogenology 2014, 81, 1101–1107. [Google Scholar] [CrossRef]
- Kershaw-Young, C.M.; Druart, X.; Vaughan, J.; Maxwell, W.M. β-Nerve growth factor is a major component of alpaca seminal plasma and induces ovulation in female alpacas. Reprod. Fertil. Dev. 2012, 24, 1093–1097. [Google Scholar] [CrossRef] [Green Version]
- Alleva, E.; Aloe, L.; Cirulli, F.; Della Seta, D.; Tirassa, P. Serum NGF levels increase during lactation and following maternal aggression in mice. Physiol. Behav. 1996, 59, 461–466. [Google Scholar] [CrossRef]
- Suda, K.; Barde, Y.A.; Thoenen, H. Nerve growth factor in mouse and rat serum: Correlation between bioassay and radioimmunoassay determinations. Proc. Natl. Acad. Sci. USA 1978, 75, 4042–4046. [Google Scholar] [CrossRef] [Green Version]
- Ando, I.; Karasawa, K.; Matsuda, H.; Tanaka, A. Changes in serum NGF levels after the exercise load in dogs: A pilot study. J. Vet. Med. Sci. 2016, 78, 1709–1712. [Google Scholar] [CrossRef]
- Lise, M.C.; Sparsa, A.; Marie, I.; Lalloué, F.; Ly, K.; Martel, C.; Bezanahary, H.; Gondran, G.; Loustaud-Ratti, V.; Bonnetblanc, J.M.; et al. Serum neurotrophin profile in systemic sclerosis. PLoS ONE 2010, 5, e13918. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | bp | |
|---|---|---|---|
| NGF | F | TCCACCCACCCAGTCTTC | 178 |
| R | GCTCGGCACTTGGTCTCA | ||
| NTRK1 | F | TCGGACCATGCTGCCCATCC | 261 |
| R | AGGCGTTGCTGCGGTTCTCG | ||
| NGFR | F | GGAGGACACGAGTCCTGAGC | 295 |
| R | CAGTGGAGAGTGCTGCAAAG | ||
| BACT | F | TTGTGATGGACTCCGGAGAC | 186 |
| R | TGATGTCACGCACGATTTCC |
| NGF | NTRK1 | NGFR | |
|---|---|---|---|
| Breeding | ++ | + | ++ |
| Nonbreeding | ++ | + | ++ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maranesi, M.; Palermo, F.A.; Bufalari, A.; Mercati, F.; Paoloni, D.; Cocci, P.; Moretti, G.; Crotti, S.; Zerani, M.; Dall’Aglio, C. Seasonal Expression of NGF and Its Cognate Receptors in the Ovaries of Grey Squirrels (Sciurus carolinensis). Animals 2020, 10, 1558. https://doi.org/10.3390/ani10091558
Maranesi M, Palermo FA, Bufalari A, Mercati F, Paoloni D, Cocci P, Moretti G, Crotti S, Zerani M, Dall’Aglio C. Seasonal Expression of NGF and Its Cognate Receptors in the Ovaries of Grey Squirrels (Sciurus carolinensis). Animals. 2020; 10(9):1558. https://doi.org/10.3390/ani10091558
Chicago/Turabian StyleMaranesi, Margherita, Francesco Alessandro Palermo, Antonello Bufalari, Francesca Mercati, Daniele Paoloni, Paolo Cocci, Giulia Moretti, Silvia Crotti, Massimo Zerani, and Cecilia Dall’Aglio. 2020. "Seasonal Expression of NGF and Its Cognate Receptors in the Ovaries of Grey Squirrels (Sciurus carolinensis)" Animals 10, no. 9: 1558. https://doi.org/10.3390/ani10091558