TGF-β1-Mediated FDNCR1 Regulates Porcine Preadipocyte Differentiation via the TGF-β Signaling Pathway

 , and
, and 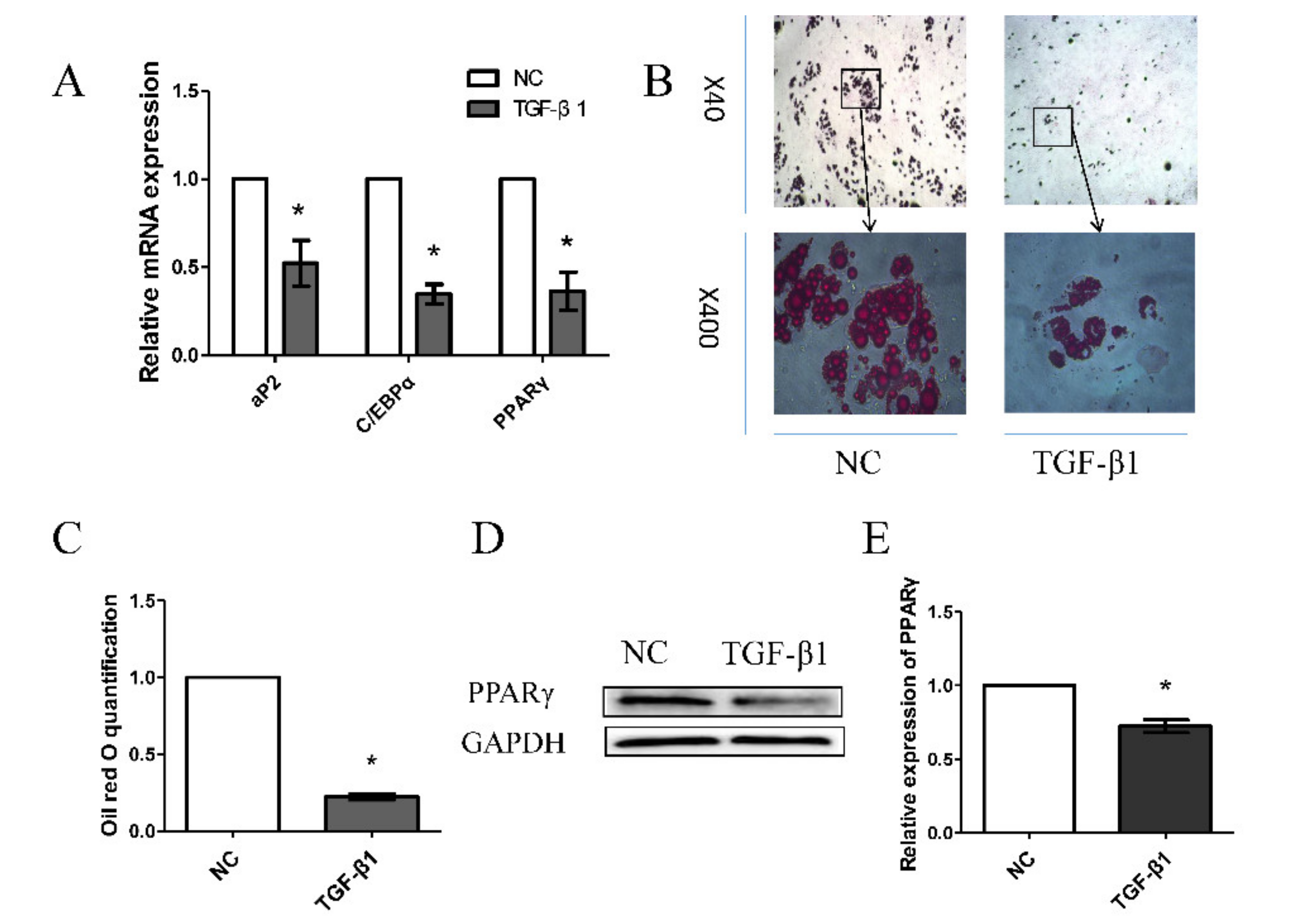{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Culture and Differentiation
2.3. RNA Isolation, Library Construction, and RNA-Seq
2.4. Transcriptome Assembly and Data Analysis
2.5. Pipeline to Identify LncRNAs
2.6. Enrichment Analysis
2.7. RT-qPCR and Western Blot Analysis
2.8. Oil Red O Staining and Triglyceride Detection
2.9. FISH
2.10. Statistical Analysis
3. Results
3.1. Effects of TGF-β1 on Porcine Preadipocyte Differentiation
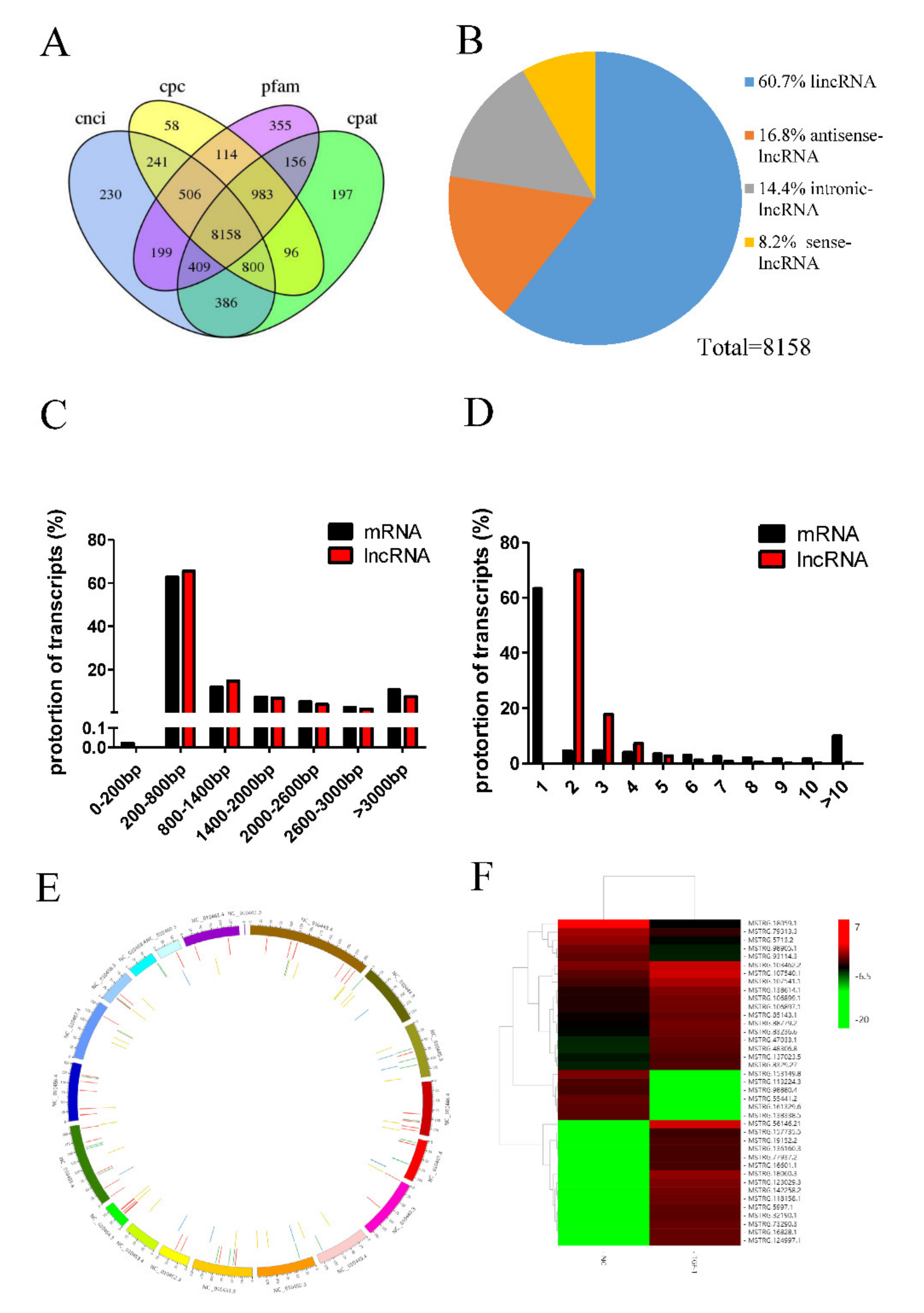
3.2. Overview of RNA-Seq
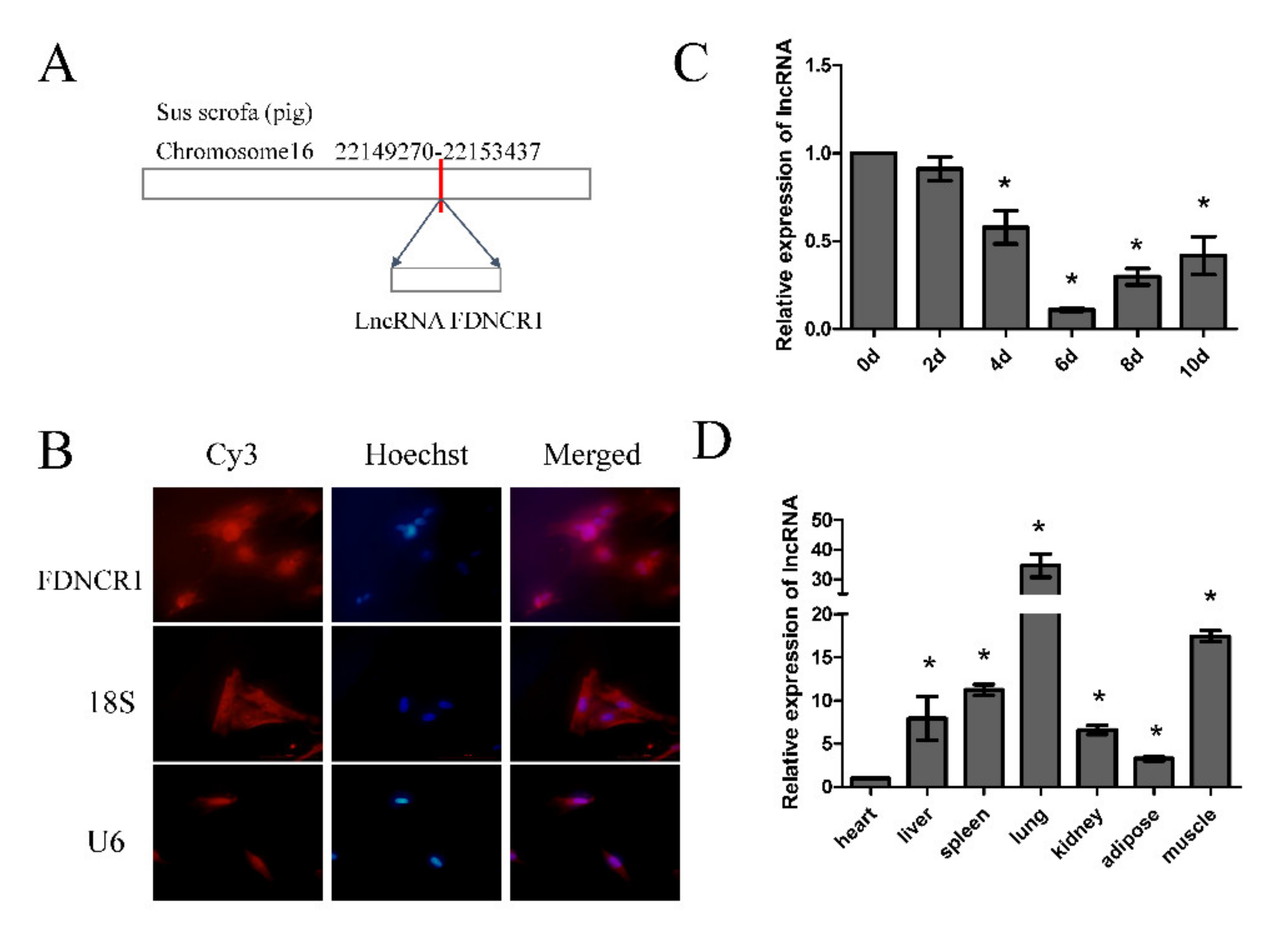
3.3. Identification of FDNCR1
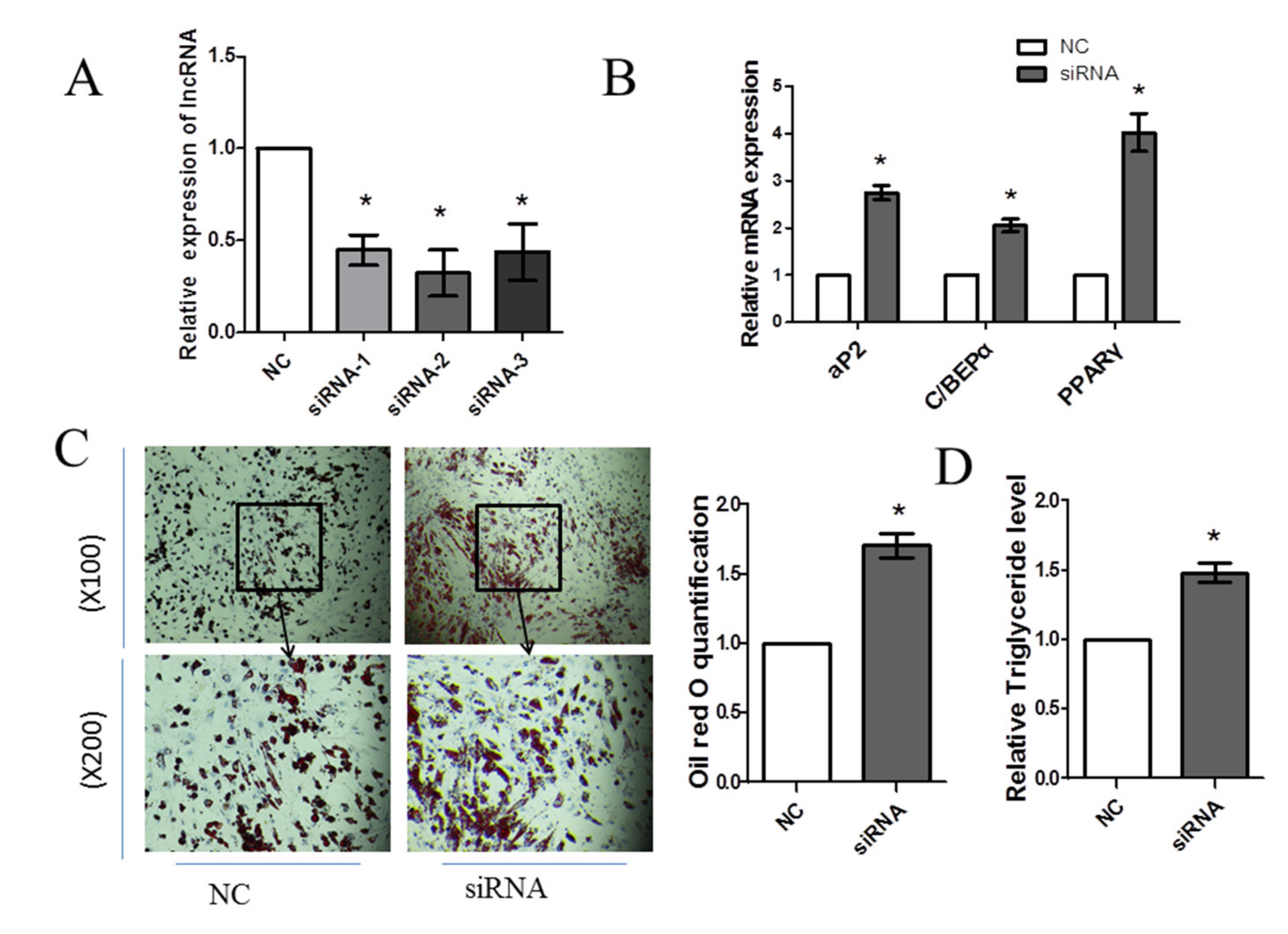
3.4. Knockdown of FDNCR1 Promotes Preadipocyte Differentiation
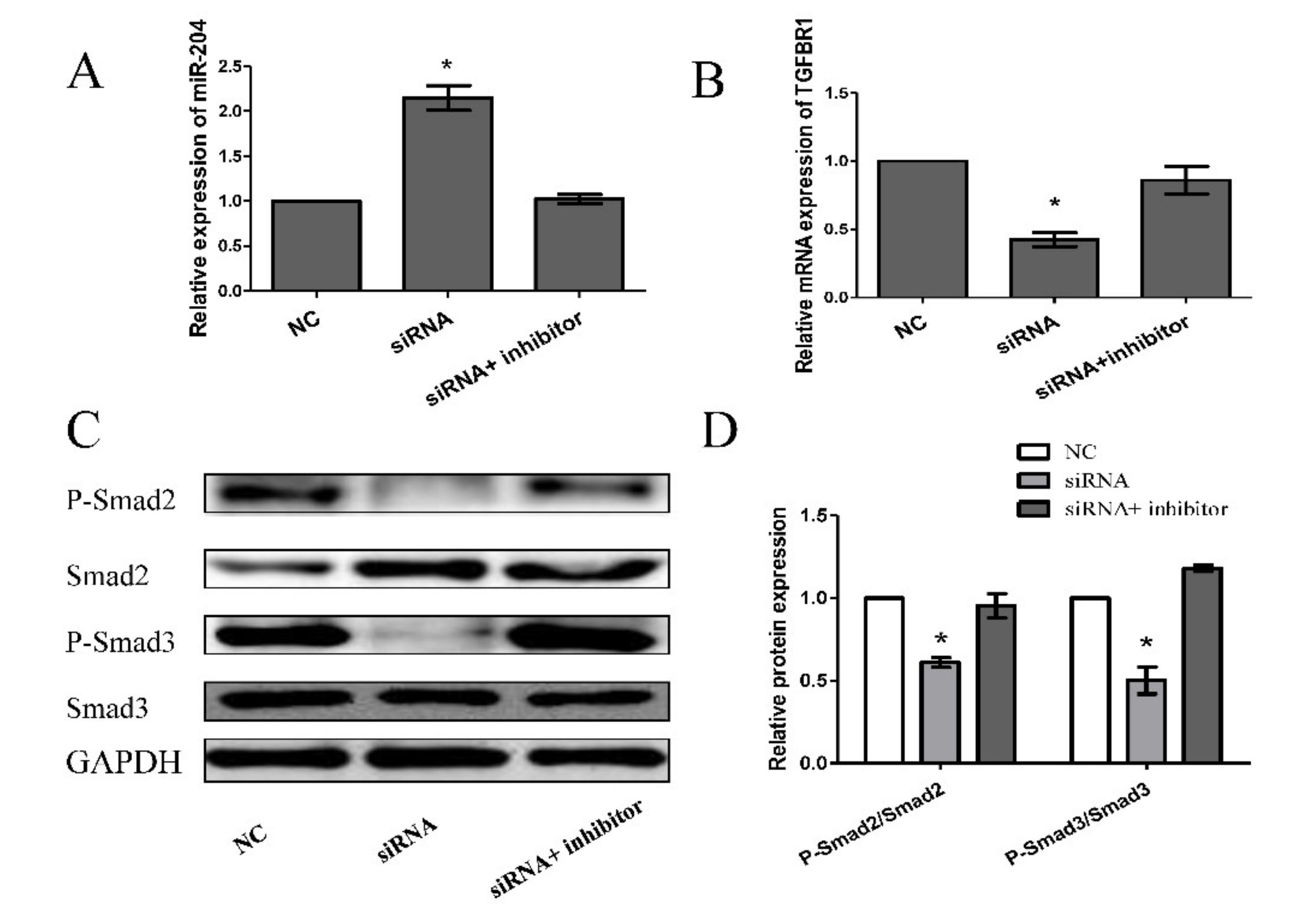
3.5. Knockdown of FDNCR1 Promotes Preadipocyte Differentiation through the TGF-β Signaling Pathway
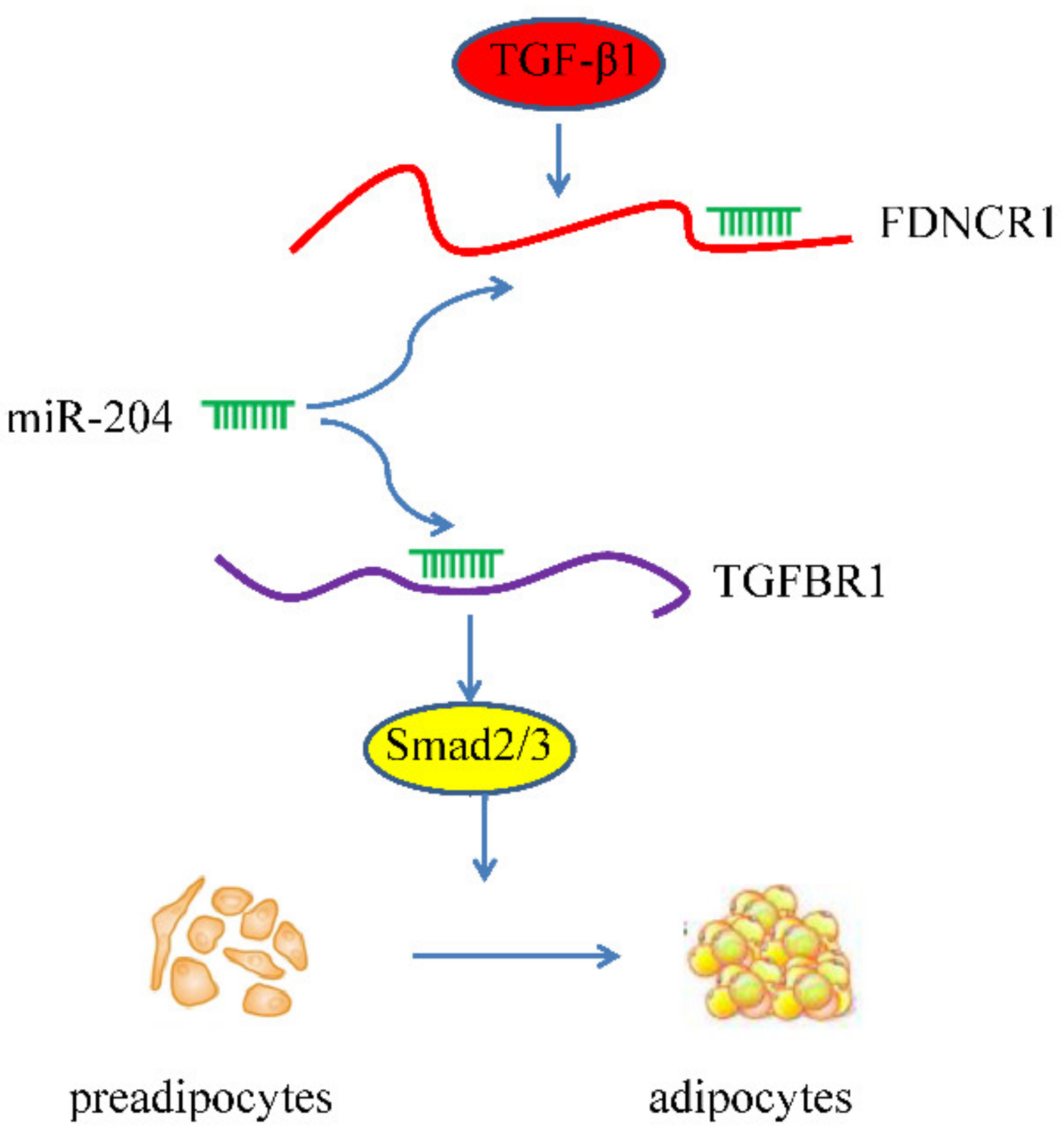
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Rosenvold, K.; Andersen, H.J. Factors of significance for pork quality-a review. Meat Sci. 2003, 64, 219–237. [Google Scholar] [CrossRef]
- Katsumata, M. Promotion of intramuscular fat accumulation in porcine muscle by nutritional regulation. Anim. Sci. J. 2011, 82, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Font-i-Furnols, M.; Tous, N.; Esteve-Garcia, E.; Gispert, M. Do all the consumers accept marbling in the same way? The relationship between eating and visual acceptability of pork with different intramuscular fat content. Meat Sci. 2012, 91, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Jaturasitha, S.; Lange, M.; Kohler, P.; Kreuzer, M.J.L.F. Quality of subcutaneous, intermuscular and intramuscular fat tissue using elevated quantities of medium chain fatty acids in pig fattening. Lipid/Fett 1996, 98, 149–156. [Google Scholar] [CrossRef]
- Fernandez, X.; Monin, G.; Talmant, A.; Mourot, J.; Lebret, B.J.M.s. Influence of intramuscular fat content on the quality of pig meat—2. Consumer acceptability of m. longissimus lumborum. Meat Sci. 1999, 53, 67–72. [Google Scholar] [CrossRef]
- Vuckovic, A.; Herber-Jonat, S.; Flemmer, A.W.; Ruehl, I.M.; Votino, C.; Segers, V.; Benachi, A.; Martinovic, J.; Nowakowska, D.; Dzieniecka, M.; et al. Increased TGF-beta: A drawback of tracheal occlusion in human and experimental congenital diaphragmatic hernia? Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L311–L327. [Google Scholar] [CrossRef] [Green Version]
- Fujio, K.; Komai, T.; Inoue, M.; Morita, K.; Okamura, T.; Yamamoto, K. Revisiting the regulatory roles of the TGF-beta family of cytokines. Autoimmun. Rev. 2016, 15, 917–922. [Google Scholar] [CrossRef]
- Goumans, M.-J.; Mummery, C. Functional analysis of the TGFb receptor/Smad pathway through gene ablation in mice. Int. J. Dev. Biol. 2000, 44, 253–265. [Google Scholar]
- Choy, L.; Skillington, J.; Derynck, R. Roles of Autocrine TGF-β Receptor and Smad Signaling in Adipocyte Differentiation. J. Cell Biol. 2000, 149, 667–682. [Google Scholar] [CrossRef]
- Souchelnytskyi, S.; Nakayama, T.; Nakao, A.; Morén, A.; Heldin, C.H.; Christian, J.L.; ten Dijke, P. Physical and Functional Interaction of Murine andXenopus Smad7 with Bone Morphogenetic Protein Receptors and Transforming Growth Factor-β Receptors. J. Biol. Chem. 1998, 273, 25364–25370. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Zhu, Y.; Nilsson, M.; Sundfeldt, K. TGF-beta isoforms induce EMT independent migration of ovarian cancer cells. Cancer Cell Int. 2014, 14, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, H.; Sakai, T. Biological Significance of Local TGF-beta Activation in Liver Diseases. Front. Physiol. 2012, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsurutani, Y.; Fujimoto, M.; Takemoto, M.; Irisuna, H.; Koshizaka, M.; Onishi, S.; Ishikawa, T.; Mezawa, M.; He, P.; Honjo, S.; et al. The roles of transforming growth factor-beta and Smad3 signaling in adipocyte differentiation and obesity. Biochem. Biophys. Res. Commun. 2011, 407, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.S.M.J.L. Discovery and annotation of long noncoding RNAs. Nat. Struct. Mol. Biol. 2015, 22, 5–7. [Google Scholar]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.A.; Aravin, A.A. Non-Coding RNAs in Transcriptional Regulation: The review for Current Molecular Biology Reports. Curr. Mol. Biol. Rep. 2015, 1, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Li, S. Expression of Concern: The functional role of long non-coding RNAs and epigenetics. Biol. Proced. Online 2016, 18, 12. [Google Scholar] [CrossRef] [Green Version]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 2015, 1253–1261. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, Y.; Xu, M.Q.; Wang, C.J.; Fu, X.H.; Liu, J.B.; Han, D.X.; Jiang, H.; Yuan, B.; Zhang, J.B. miR-181a regulate porcine preadipocyte differentiation by targeting TGFBR1. Gene 2019, 681, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Orom, U.A.; Derrien, T.; Beringer, M.; Gumireddy, K.; Gardini, A.; Bussotti, G.; Lai, F.; Zytnicki, M.; Notredame, C.; Huang, Q.; et al. Long noncoding RNAs with enhancer-like function in human cells. Cell 2010, 143, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Da Wei Huang, B.T.S.R.A.L. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Jaiswal, S.K.; Gupta, A.; Saxena, R.; Prasoodanan, V.P.K.; Sharma, A.K.; Mittal, P.; Roy, A.; Shafer, A.B.A.; Vijay, N.; Sharma, V.K. Genome Sequence of Peacock Reveals the Peculiar Case of a Glittering Bird. Front. Genet. 2018, 9, 392. [Google Scholar] [CrossRef]
- Shi, Q.; Wang, X.; Cai, C.; Yang, S.; Huo, N.; Liu, H. Association between TGF-beta1 Polymorphisms and Head and Neck Cancer Risk: A Meta-Analysis. Front. Genet. 2017, 8, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Niu, J.; Xi, G.; Niu, X.; Wang, Y.; Guo, M.; Yangzong, Q.; Yao, Y.; Sizhu, S.L.; Tian, J. TGF-beta1 resulting in differential microRNA expression in bovine granulosa cells. Gene 2018, 663, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Hills, C.; Price, G.W.; Wall, M.J.; Kaufmann, T.J.; Chi-Wai Tang, S.; Yiu, W.H.; Squires, P.E. Transforming Growth Factor Beta 1 Drives a Switch in Connexin Mediated Cell-to-Cell Communication in Tubular Cells of the Diabetic Kidney. Cell Physiol. Biochem. 2018, 45, 2369–2388. [Google Scholar] [CrossRef] [Green Version]
- Qi, R.; Yang, F.; Huang, J.; Peng, H.; Liu, Y.; Liu, Z. Supplementation with conjugated linoeic acid decreases pig back fat deposition by inducing adipocyte apoptosis. BMC Vet. Res. 2014, 10, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Cai, R.; Wang, Y.; Zhao, R.; Qin, J.; Pang, W. A Newly Identified LncRNA LncIMF4 Controls Adipogenesis of Porcine Intramuscular Preadipocyte through Attenuating Autophagy to Inhibit Lipolysis. Animals 2020, 10, 926. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, H.; Mao, R.; Yang, H.; Zhang, Y.; Zhang, Y.; Guo, P.; Zhan, D.; Xiang, B.; Liu, Y. The lncRNA RP11-142A22.4 promotes adipogenesis by sponging miR-587 to modulate Wnt5β expression. Cell Death Dis. 2020, 11, 475. [Google Scholar] [CrossRef]
- Cai, H.; Li, M.; Jian, W.; Song, C.; Huang, Y.; Lan, X.; Lei, C.; Chen, H. A novel lncRNA BADLNCR1 inhibits bovine adipogenesis by repressing GLRX5 expression. J. Cell. Mol. Med. 2020, 24, 7175–7186. [Google Scholar] [CrossRef]
- Ding, J.; Li, J.; Wang, H.; Tian, Y.; Xie, M.; He, X.; Ji, H.; Ma, Z.; Hui, B.; Wang, K.; et al. Long noncoding RNA CRNDE promotes colorectal cancer cell proliferation via epigenetically silencing DUSP5/CDKN1A expression. Cell Death Dis. 2017, 8, e2997. [Google Scholar] [CrossRef]
- Li, C.X.; Li, H.G.; Huang, L.T.; Kong, Y.W.; Chen, F.Y.; Liang, J.Y.; Yu, H.; Yao, Z.R. H19 lncRNA regulates keratinocyte differentiation by targeting miR-130b-3p. Cell Death Dis. 2017, 8, e3174. [Google Scholar] [CrossRef]
- Hall, J.R.; Messenger, Z.J.; Tam, H.W.; Phillips, S.L.; Recio, L.; Smart, R.C. Long noncoding RNA lincRNA-p21 is the major mediator of UVB-induced and p53-dependent apoptosis in keratinocytes. Cell Death Dis. 2015, 6, e1700. [Google Scholar] [CrossRef]
- Krishnan, J.; Mishra, R.K. Emerging trends of long non-coding RNAs in gene activation. FEBS J. 2014, 281, 34–45. [Google Scholar] [CrossRef]
- Weinberg, M.S.; Morris, K.V. Long Non-Coding RNA Targeting and Transcriptional De-Repression. Nucleic Acid Ther. 2013, 23, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouvenot, Y.; Poirier, F.; Jami, J.; Paldi, A. Biallelic transcription of Igf2 and H19 in individual cells suggests a post-transcriptional contribution to genomic imprinting. Curr. Biol. 1999, 9, 1199–1202. [Google Scholar] [CrossRef] [Green Version]
- Kouba, M.; Bonneau, M.; Noblet, J. Relative development of subcutaneous, intermuscular, and kidney fat in growing pigs with different body compositions. J. Anim. Sci. 1999, 77, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.W.; D’Agostino, R.B.; Sullivan, L.; Parise, H.; Kannel, W.B. Overweight and obesity as determinants of cardiovascular risk: The Framingham experience. Arch. Intern. Med. 2002, 162, 1867–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.C.; Jiao, Y.; Zhang, Y.Y.; Ning, J.; Zhang, Y.R.; Xu, J.; Wei, W.; Kang-Sheng, G. Lnc SMAD5-AS1 as ceRNA inhibit proliferation of diffuse large B cell lymphoma via Wnt/beta-catenin pathway by sponging miR-135b-5p to elevate expression of APC. Cell Death Dis. 2019, 10, 252. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, W.; Jin, M.; Chen, J.; Xu, W.; Kong, X. lncRNA MIAT functions as a competing endogenous RNA to upregulate DAPK2 by sponging miR-22-3p in diabetic cardiomyopathy. Cell Death Dis. 2017, 8, e2929. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, W.; Liu, J.B.; Wang, Y.; Hao, J.D.; Huang, Y.J.; Gao, Y.; Jiang, H.; Yuan, B.; Zhang, J.B. ssc-miR-204 regulates porcine preadipocyte differentiation and apoptosis by targeting TGFBR1 and TGFBR2. J. Cell. Biochem. 2020, 121, 609–620. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Meng, Y.; Gao, F.; Xiao, Y.; Zheng, Y.; Wang, H.-Q.; Gao, Y.; Jiang, H.; Yuan, B.; Zhang, J.-B. TGF-β1-Mediated FDNCR1 Regulates Porcine Preadipocyte Differentiation via the TGF-β Signaling Pathway. Animals 2020, 10, 1399. https://doi.org/10.3390/ani10081399
Zhang Z, Meng Y, Gao F, Xiao Y, Zheng Y, Wang H-Q, Gao Y, Jiang H, Yuan B, Zhang J-B. TGF-β1-Mediated FDNCR1 Regulates Porcine Preadipocyte Differentiation via the TGF-β Signaling Pathway. Animals. 2020; 10(8):1399. https://doi.org/10.3390/ani10081399
Chicago/Turabian StyleZhang, Zhe, Yu Meng, Fei Gao, Yue Xiao, Yi Zheng, Hao-Qi Wang, Yan Gao, Hao Jiang, Bao Yuan, and Jia-Bao Zhang. 2020. "TGF-β1-Mediated FDNCR1 Regulates Porcine Preadipocyte Differentiation via the TGF-β Signaling Pathway" Animals 10, no. 8: 1399. https://doi.org/10.3390/ani10081399