Identification and Functional Annotation of Genes Related to Horses’ Performance: From GWAS to Post-GWAS

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
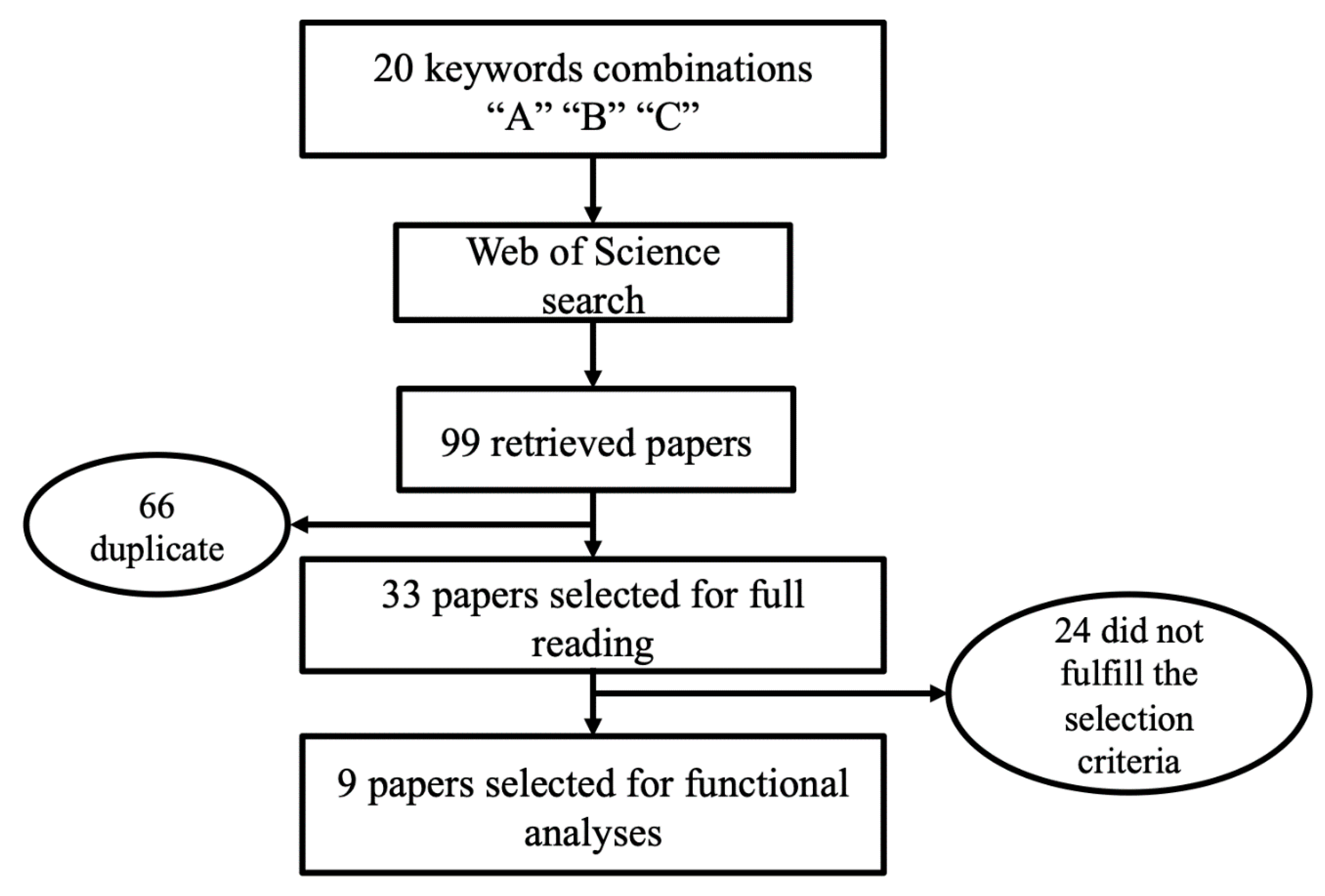
2.1. Identification of Candidate Genes—Systematic Review
2.2. Functional Annotation—Gene Networks
3. Results
3.1. Systematic Review
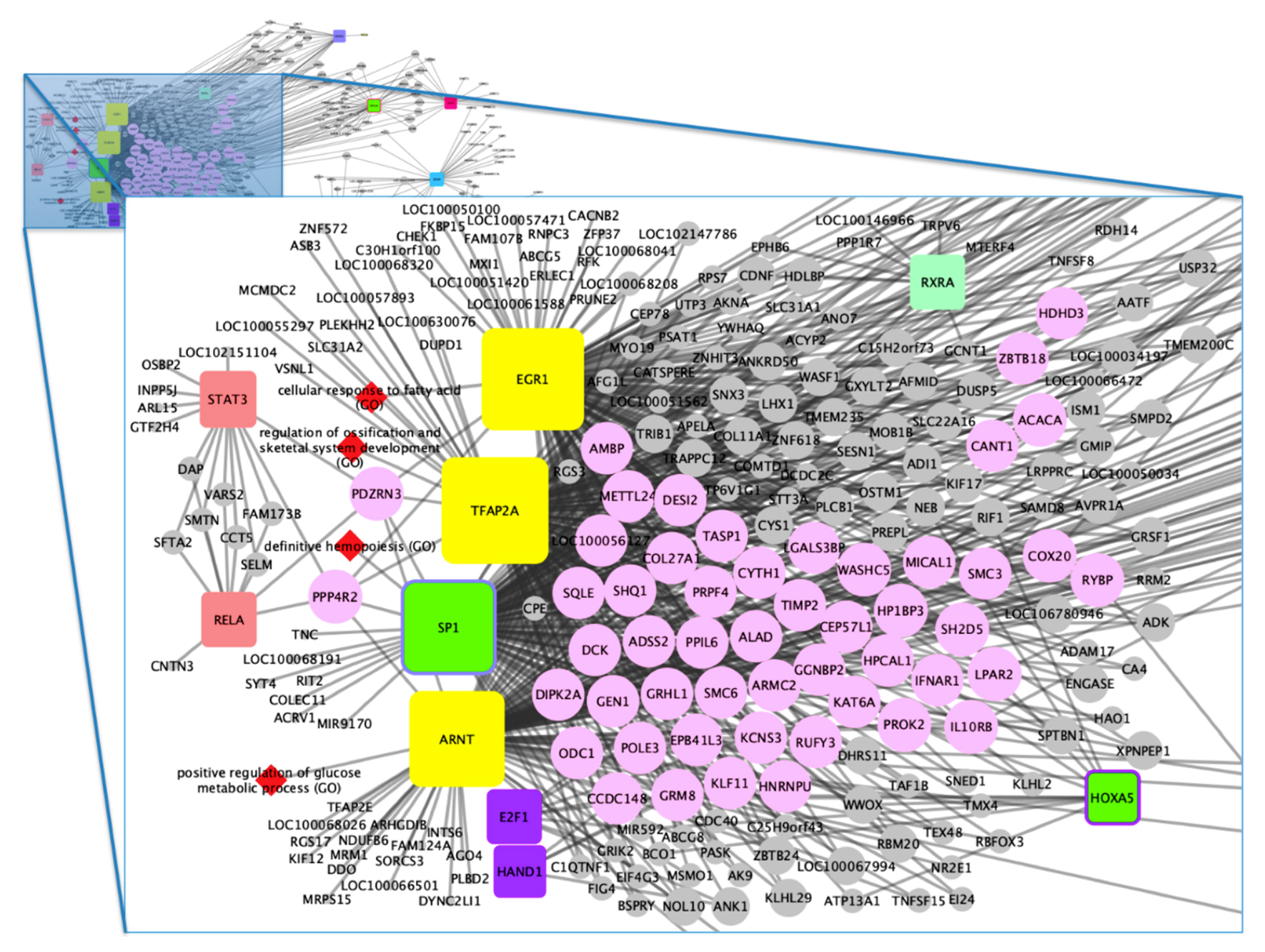
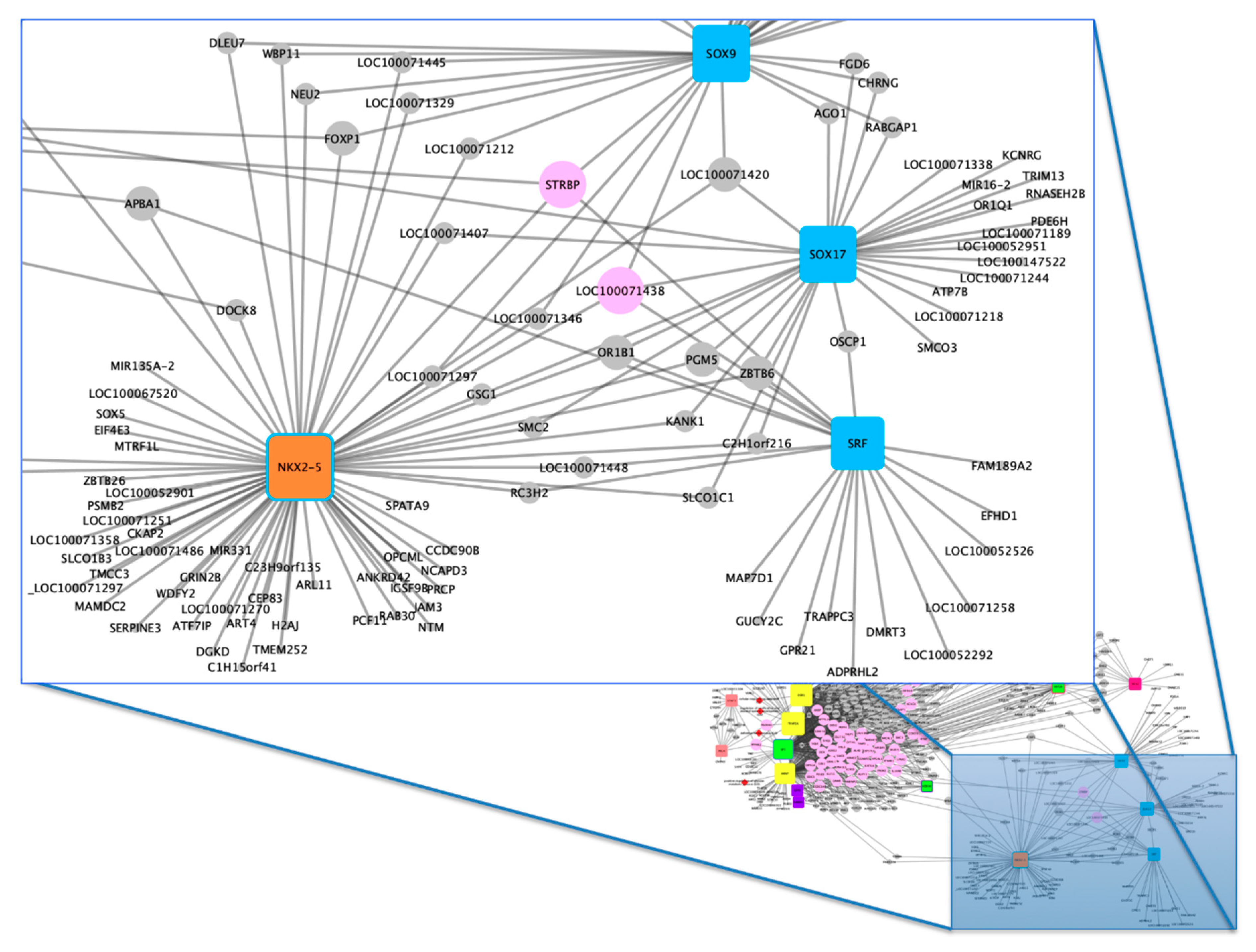
3.2. Functional Analysis
4. Discussion
4.1. Gene–Biological Process Network
4.2. Gene–TF Network
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Levine, M.A. Domestication and early history of the horse. In The Domestic Horse: The Evolution, Development, and Management of Its Behaviour; Mills, D., McDonell, S., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 5–22. [Google Scholar]
- FAOSTAT. Production statistics of the Food and Agricultural Organization of the United Nations. 2017. Available online: http://www.fao.org/faostat/en/#data/QA (accessed on 19 December 2019).
- Cross, P. Global Horse statistics internal 02 2019. HPBA 2019, 1–19. [Google Scholar]
- Pereira, L.P.; Regatieri, I.C.; Ferraz, G.C.; Queiroz Neto, A.; Curi, R.A. Perspectivas do uso de marcadores moleculares no melhoramento genético de equinos de corrida da raça Quarto de Milha. Vet. Zootec. 2015, 22, 347–369. [Google Scholar]
- Fortes, M.R.S.; Reverter, A.; Zhang, Y.; Collis, E.; Nagaraj, S.H.; Jonsson, N.N.; Prayaga, K.C.; Barris, W.; Hawken, R.J. Association weight matrix for the genetic dissection of puberty in beef cattle. Proc. Natl. Acad. Sci. USA 2010, 107, 13642–13647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verardo, L.L.; Silva, F.F.; Lopes, M.S.; Madsen, O.; Bastiaansen, J.W.M.; Knol, E.F.; Kelly, M.; Varona, L.; Lopes, P.S.; Guimarães, S.E.F. Revealing new candidate genes for reproductive traits in pigs: Combining Bayesian GWAS and functional pathways. Genet. Sel. Evol. 2016, 48, 9. [Google Scholar] [CrossRef] [Green Version]
- Soares, A.C.C.; Guimarães, S.E.F.; Kelly, M.J.; Fortes, M.R.S.; Silva, F.F.; Verardo, L.L.; Mota, R.R.; Moore, S. Multiple-trait genomewide mapping and gene network analysis for scrotal circumference growth curves in Brahman cattle. J. Anim. Sci. 2017, 95, 3331–3345. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGo: A Cystoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome. Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Tonon, L.; Touzet, H.; Varré, J.S. TFM-Explorer: Mining cis-regulatory regions in genomes. Nucleic Acids Res. 2010, 38, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Sandelin, A.; Alkema, W.; Engstrom, P.; Wasserman, W.W.; Lenhard, B. JASPAR: An open-access database for eukaryotic transcription factor binding profiles. Nucleic Acids Res. 2004, 32, 91–94. [Google Scholar] [CrossRef] [Green Version]
- Touzet, H.; Varré, J.-S. Efficient and accurate P-value computation for Position Weight Matrices. Algorithm Mol. Biol. 2007, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A Cytoscape plugin to assess overrepresentation of Gene Ontology categories in Biological Networks. Bionformatics 2005, 21, 3448–3449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meira, C.T.; Farah, M.M.; Fortes, M.R.S.; Moore, S.S.; Pereira, G.L.; de Vasconcelos Silva, J.A., II; Mota, M.D.S.; Curi, R.A. A genome-wide association study for morphometric traits in Quarter Horse. J. Equine Vet. Sci. 2014, 34, 1028–1031. [Google Scholar] [CrossRef] [Green Version]
- Meira, C.T.; Fortes, M.M.; Farah, M.R.S.; Porto-Neto, L.R.; Kelly, M.; Moore, S.S.; Pereira, G.L.; Chardulo, L.A.L.; Curi, R.A. Speed index in the racing Quarter Horse: A genome-wide association study. J. Equine Vet. Sci. 2014, 34, 1263–1268. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.-H.; Lee, J.W.; Park, J.-E.; Choi, I.-Y.; Oh, H.-S.; Kim, H.J.; Kim, H. Multiple genes related to muscle identified through a joint analysis of two-stage genome-wide association study for racing performance of 1,156 Thoroughbreds. Asian Australas. J. Anim. Sci. 2015, 28, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Ricard, A.; Robert, C.; Blouin, C.; Baste, F.; Torquet, G.; Morgenthaler, C.; Rivière, J.; Mach, N.; Mata, X.; Schibler, L.; et al. Endurance exercise ability in the horse: A trait with complex polygenic determinism. Front. Genet. 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velie, B.D.; Fegraeus, K.J.; Solé, M.; Rosengren, M.K.; Røed, K.H.; Ihler, C.-F.; Strand, E.; Lindgren, G. A genome-wide association study for harness racing success in the Norwegian-Swedish coldblooded trotter reveals genes for learning and energy metabolism. BMC Genet. 2018, 19, 1–13. [Google Scholar] [CrossRef]
- Pereira, G.L.; Chardulo, L.A.; de Vasconcelos Silva, J.A., II; Faria, R.; Curi, R.A. Genomic regions associated with performance in racing line of Quarter Horses. Livest. Sci. 2018, 211, 42–51. [Google Scholar] [CrossRef] [Green Version]
- McGivney, B.A.; Hernandez, B.; Katz, L.M.; MacHugh, D.E.; McGovern, S.P.; Parnell, A.C.; Wiencko, H.L.; Hill, E.W. A genomic prediction model for racecourse starts in the Thoroughbred horse. Anim. Genet. 2019, 50, 347–357. [Google Scholar] [CrossRef]
- Gmel, A.I.; Druml, T.; Niederhäusern, R.; Leeb, T.; Neuditschko, M. Genome-wide association studies based on equine joint angle measurements reveal new QTL affecting the conformation of horses. Genes 2019, 10, 370. [Google Scholar] [CrossRef] [Green Version]
- Bussiman, F.O.; Santos, B.A.; Silva, B.C.A.; Mamani, G.C.M.; Grigoletto, L.; Pereira, G.L.; Ferraz, G.C.; Ferraz, J.B.S.; Mattos, E.C.; Eler, J.P.; et al. Genome-wide association study: Understanding the genetic basis of the gait type in Brazilian Mangalarga Marchador horses, a preliminary study. Livest. Sci. 2020, 231, 1–11. [Google Scholar] [CrossRef]
- Landry-Truchon, K.; Houde, N.; Boucherat, O.; Joncas, F.-H.; Dasen, J.S.; Philippidou, P.; Mansfield, J.H.; Jeannotte, L. HOXA5 plays tissue-specific roles in the developing respiratory system. Co. Biol. 2017, 144, 3547–3561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, S.D.; Rundqvist, H.; Papandreou, I.; Duh, R.; McNulty, W.J.; Howlett, R.A.; Olfert, I.M.; Sundberg, C.J.; Denko, N.C.; Poellinger, L.; et al. HIF-1∂ in endurance training: Suppression of oxidative metabolism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, 2059–2069. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Suzuki, R.; Lee, K.; Tran, T.; Gunton, J.E.; Saha, A.K.; Patti, M.-E.; Goldfine, A.; Ruderman, N.B.; Gonzalez, F.J.; et al. Ablation of ARNT/HIF1β in liver alters gluconeogenesis, lipogenic gene expression, and serum ketones. Cell Metab. 2009, 9, 428–439. [Google Scholar] [CrossRef] [Green Version]
- Wenke, A.-K.; Bosserhoff, A.K. Roles of AP-2 transcription factors in the regulation of cartilage and skeletal development. FEBS J. 2010, 277, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.S.; Degnin, C.; Fiddler, T.A.; Stauffer, D.; Thayer, M.J. Regulation of MyoD activity and muscle cell differentiation by MDM2, pRb, and Sp1. J. Biol. Chem. 2003, 278, 22615–22622. [Google Scholar] [CrossRef] [Green Version]
- Gokey, N.G.; Lopez-Anido, C.; Gillian-Daniel, A.L.; Svaren, J. Early growth response 1 (Egr1) regulates cholesterol biosynthetic gene expression. J. Biol. Chem. 2011, 286, 29501–29510. [Google Scholar] [CrossRef] [Green Version]
- George, R.M.; Firulli, A.B. Hand factors in cardiac development. Anat. Rec. 2019, 302, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Denechaud, P.-D.; Fajas, L.; Giralt, A. E2F1, a novel regulator of metabolism. Front. Endocrinol. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Begue, G.; Douillard, A.; Galbes, O.; Rossano, B.; Vernus, B.; Candau, R.; Py, G. Early activation of rat skeletal muscle IL-6/STAT1/STAT3 dependent gene expression in resistance exercise linked to hypertrophy. PLoS ONE 2013, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Fukai, A.; Mabuchi, A.; Ikeda, T.; Yano, F.; Ohba, S.; Nishida, N.; Akune, T.; Yoshimura, N.; Nakagawa, T.; et al. Transcriptional regulation of endochondral ossification by HIF-2∂ during skeletal growth and osteoarthritis development. Nat. Med. 2010, 16, 678–687. [Google Scholar] [CrossRef]
- Lefebvre, V.; Angelozzi, M.; Haseeb, A. SOX9 in cartilage development and disease. Curr. Opin. Cell Biol. 2019, 61, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Saba, R.; Kitajima, K.; Rainbow, L.; Engert, S.; Uemura, M.; Ishida, H.; Kokkinopoulos, I.; Shintani, Y.; Miyagawa, S.; Kanai, Y.; et al. Endocardium differentiation through Sox17 expression in endocardium precursor cells regulates heart development in mice. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Guerci, A.; Lahoute, C.; Hébrard, S.; Collard, L.; Graindorge, D.; Favier, M.; Cagnard, N.; Batonnet-Pichon, S.; Précigout, G.; Garcia, L.; et al. Srf-dependent paracrine signals produced by miofibers control satellite cell-mediated skeletal muscle hypertrophy. Cell Metab. 2011, 15, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, S.; Kume, T. Forkhead transcription factors, Foxc1 and Foxc2, are required for the morphogenesis of the cardiac outflow tract. Dev. Biol. 2006, 296, 421–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gontan, C.; Munck, A.; Vermeij, M.; Grosveld, F.; Tibboel, D.; Rottier, R. Sox2 is important for two crucial processes in lung development: Branching morphogenesis and epithelial cell differentiation. Dev. Biol. 2008, 317, 296–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merki, E.; Zamora, M.; Raya, A.; Kawakami, Y.; Wang, J.; Zhang, X.; Burch, J.; Kubalak, S.W.; Kaliman, P.; Belmonte, J.C.I.; et al. Epicardial retinoid X receptor ∂ is required for myocardial growth and coronary artery formation. Proc. Natl. Acad. Sci. USA 2005, 102, 18455–18460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKeever, K.H.; Hinchcliff, K.W. Neuroendocrine control of blood volume, blood pressure and cardiovascular function in horses. Equine Vet. J. 1995, 18, 77–81. [Google Scholar] [CrossRef]
- Hew-Butler, T. Arginine vasopressin, fluid balance and exercise. Sports Med. 2010, 40, 459–479. [Google Scholar] [CrossRef]
- Hew-Butler, T.; Noakes, T.D.; Soldin, S.J.; Verbalis, J.G. Acute changes in endocrine and fluid balance markers during high-intensity, steady-state, and prolonged endurance running: Unexpected increases in oxytocin and brain natriuretic peptide during exercise. Eur. J. Endocrinol. 2008, 159, 729–737. [Google Scholar] [CrossRef] [Green Version]
- Peake, J.; Della Gatta, P.; Suzuki, K.; Nieman, D. Cytokine expression and secretion by skeletal muscle cells: Regulatory mechanisms and exercise effects. Exerc. Immunol. Rev. 2015, 21, 8–25. [Google Scholar] [PubMed]
- Pedersen, B.K. Muscles and their myokines. J. Exp. Biol. 2011, 214, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capomaccio, S.; Cappelli, K.; Barrey, E.; Felicetti, M.; Silvestrelli, M.; Verini-Supplizi, A. Microarray analysis after strenuous exercise in peripheral blood mononuclear cells of endurance horses. Anim. Genet. 2010, 41, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Tiburcio-Félix, R.; Cisneros, B.; Hernández-Kelly, L.C.R.; Hernández-Contreras, M.A.; Luna-Herrera, J.; Rea-Hernández, I.; Jiménez-Aguilar, R.; Olivares-Bañuelos, T.N.; Ortega, A. Neuronal nitric oxide synthase in cultured cerebellar Bergmann glia: Glutamate-dependent regulation. ACS Chem. Neurosci. 2019, 10, 2668–2675. [Google Scholar] [CrossRef]
- Coussen, F. Molecular determinants of kainate receptor trafficking. Neuroscience 2009, 158, 25–35. [Google Scholar] [CrossRef] [PubMed]
- NCBI. National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 13 May 2020).
- Carobrez, A.P. Transmissão pelo glutamate como alvo molecular na ansiedade. Rev. Bras. Psiquiatr. 2003, 25, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.; Arkins, S. Equine learning behaviour. Behav. Process. 2007, 76, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Marinier, S.L.; Alexander, A.J. The use of maze in testing learning and memory in horses. Appl. Anim. Behav. Sci. 1994, 39, 177–182. [Google Scholar] [CrossRef]
- Girard, I.; Garland, T., Jr. Plasma corticosterone response to acute and chronic voluntary exercise in female house mice. J. Appl. Physiol. 2002, 92, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Eide, D.J. The SLC39 family of metal ion transporters. Pflügers Arch. 2004, 447, 796–800. [Google Scholar] [CrossRef]
- Clarkson, P.M.; Thompson, H.S. Antioxidants: What a role do they play in physical activity and health? Am. J. Clin. Nutr. 2000, 72, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Koury, J.C.; Donangelo, C.M. Zinco, estresse oxidativo e atividade física. Rev. Nutr. 2003, 16, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Goldfarb, A.H. Nutritional antioxidants as therapeutic and preventive modalities in exercise-induced muscle damage. Can. J. Appl. Physiol. 1999, 24, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Ngan, E.S.W.; Tam, P.K.H. Prokineticin-signaling pathway. Int. J. Biochem. Cell Biol. 2008, 40, 1679–1684. [Google Scholar] [CrossRef]
- Silva, A.S.; Zanesco, A. Exercício físico, receptores β-adrenérgicos e resposta vascular. J. Vasc. Bras. 2010, 9, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Marlin, D.; Nankervis, K. Equine Exercise Physiology, 1st ed.; Blackwell Science: London, UK, 2002; pp. 1–296. [Google Scholar]
- Silveira, L.R.; Pinheiro, C.H.J.; Zoppi, C.C.; Hirabara, S.M.; Vitzel, K.F.; Bassit, R.A.; Leandro, C.G.; Barbosa, M.R.; Sampaio, I.H.; Melo, I.H.P.; et al. Regulação do metabolismo de glicose e ácido graxo no músculo esquelético durante exercício físico. Arq. Bras. Endocrinol. Metab. 2011, 55, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Tabrizian, T.; Hataway, F.; Murray, D.; Shariat-Madar, Z. Prolylcarboxypeptidase gene expression in the heart and kidney: Effects of obesity and diabetes. Cardiovasc. Hematol. Agents Med. Chem. 2015, 13, 113–123. [Google Scholar] [CrossRef]
- Maier, C.; Schadock, I.; Haber, P.K.; Wysocki, J.; Ye, M.; Kanwar, Y.; Flask, C.A.; Yu, X.; Hoit, B.D.; Adams, G.N.; et al. Prolylcarboxypeptidase deficiency is associated with increased bloos pressure, glomerular lesions, and cardiac dysfunction independent of altered circulating and cardiac angiotensin II. J. Mol. Med. 2017, 95, 473–486. [Google Scholar] [CrossRef]
- Moreno, M.; Ortega, F.; Xifra, G.; Ricart, W.; Fernández-Real, J.M.; Moreno-Navarrete, J.M. Cytosolic aconitase activity sustains adipogenic capacity of adipose tissue connecting iron metabolism and adipogenesis. FASEB J. 2015, 29, 1529–1539. [Google Scholar] [CrossRef] [Green Version]
- Machado, L.P.; Kohayagawa, A.; Yonezawa, L.A.; Silveira, V.F.; Saito, M.E. Metabolismo do ferro em equinos atletas. Cienc. Rural 2010, 40, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Minini, R.A.B.; Laposy, C.B.; Neto, H.B.; Melchert, A.; Giuffrida, R.; Rossi, H.; Valle, H.F.D. Concentrações de ferro, cobre, zinco e manganês em equinos da raça Puro-sangue Lusitano, antes e após exercício. Pesq. Vet. Bras. 2013, 33, 1045–1048. [Google Scholar] [CrossRef] [Green Version]
- Ramnanan, C.J.; Edgerton, D.S.; Kraft, G.; Cherrington, A.D. Physiologic action of glucagon on liver glucose metabolism. Diabetes Obes. Metab. 2011, 13, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Canali, E.S.; Kruel, L.F.M. Respostas hormonais ao exercício. Rev. Paul. Educ. Fís. 2001, 15, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Khoo, E.Y.H.; Wallis, J.; Tsintzas, K.; Macdonald, I.A.; Mansell, P. Effects of exenatide on circulating glucose, insulin, glucagon, cortisol and catecholamines in healthy volunteers during exercise. Diabetologia 2010, 53, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nottoli, T.; Hagopian-Donaldson, S.; Zhang, J.; Perkins, A.; Williams, T. AP-2-null cells disrupt morphogenesis of the eye, face, and limbs in chimeric mice. Proc. Natl. Acad. Sci. USA 1998, 95, 13714–13719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Wilke, T.; Ashique, A.M.; Narvey, M.; Zerucha, T.; Savino, E.; Williams, T.; Richman, J.M. Chicken transcription factor AP-2: Cloning, expression and its role in outgrowth of facial prominences and limb buds. Dev. Biol. 1997, 188, 248–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, M.B.; Abu-Seida, A.M.; El-Glil, A.I.A. Radiographical, ultrasonographic and arthroscopic findings of osteochondrosis dissecans of the tarsocrural joint of horses. Res. Opin. Anim. Vet. Sci. 2014, 4, 318–322. [Google Scholar]
- Mostafa, M.B.; Abu-Seida, A.M.; El-Glil, A.I.A. Septic tarsitis in horses: Clinical, radiological, ultrasonographic, arthroscopic and bacteriological findings. Res. Opin. Anim. Vet. Sci. 2014, 4, 30–34. [Google Scholar]
- Barcelos, K.M.C.; Rezende, A.S.C.; Biggi, M.; Lana, A.M.Q.; Maruch, S.; Faleiros, R.R. Prevalence of tarsal diseases in champion Mangalarga Marchador horses in the marcha picada modality and its association with tarsal angle. J. Equine Vet. Sci. 2016, 47, 25–30. [Google Scholar] [CrossRef]
- Foss, M.A.; Wickler, S.J. Veterinary aspects of endurance riding. In Equine Sports Medicine and Surgery, 1st ed.; Hinchcliff, K., Kaneps, A.J., Geor, R.J., Eds.; Saunders Elsevier: Philadelphia, PA, USA, 2004; pp. 1105–1117. [Google Scholar]
- Moreira, D.O.; Marques, M.M.; Leão, N.F.; Viana, W.S.; Oliveira, F.P.L.; Faleiros, R.R.; Silveira, G.H.A. Concentrações de proteínas totais, glicose, cálcio, fósforo, lactate, ureia e creatinina em equinos de cavalaria militar antes e após trabalho de patrulhamento urbano. Cienc. Anim. Bras. 2015, 16, 73–80. [Google Scholar] [CrossRef]
- Padalino, B.; Rubino, G.; Centoducati, P.; Petazzi, F. Training versus Overtraining: Evaluation of two protocols. J. Equine Vet. Sci. 2007, 27, 28–31. [Google Scholar] [CrossRef]
- Brand, E.; Herrman, S.-M.; Nicaud, V.; Evans, A.; Ruidavets, J.-B.; Arvelier, D.; Luc, G.; Cambien, F.; Soubrier, F. Identification of two polymorphisms in the early growth response protein-1 gene: Possible association with lipid variables. J. Mol. Med. 2000, 78, 81–86. [Google Scholar] [CrossRef]
- Noleto, P.G.; Santos, J.B.F.; Rocha, F.M.; Fasano, P.E.; Guimarães, E.C.; Mundim, A.V. Effect of a 130 km endurance ride on the serum biochemical profiles of Mangalarga Marchador horses. J. Equine Vet. Sci. 2016, 39, 7–11. [Google Scholar] [CrossRef]
- Onmaz, A.C.; Van Den Hoven, R.; Gunes, V.; Cinar, M.; Kucuk, O. Oxidative stress in horses after a 12-hours transport period. Revue. Méd. Vét. 2011, 162, 213–217. [Google Scholar]
- Ferraz, G.C.; Teixeira-Neto, A.R.; Pereira, M.C.; Linardi, R.L.; Lacerda-Neto, J.C.; Queiroz-Neto, A. Influência do treinamento aeróbio sobre o cortisol e glicose plasmáticos em equinos. Arq. Bras. Med. Vet. Zootec. 2010, 62, 23–29. [Google Scholar] [CrossRef]
- Vyroubal, P.; Chiarla, C.; Giovannini, I.; Hyspler, R.; Ticha, A.; Hrnciarikova, D.; Zadak, Z. Hypocholesterolemia in clinically serious conditions–Review. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2008, 152, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Irrcher, I.; Hood, D.A. Regulation of Egr-1, and Sp1 mRNA expression in contracting skeletal muscle cells. J. Appl. Physiol. 2004, 97, 2207–2213. [Google Scholar] [CrossRef] [Green Version]
- Hood, D.A. Invited review: Contractile activity-induced mitochondrial biogenesis in skeletal muscle. J. Appl. Physiol. 2001, 90, 1137–1157. [Google Scholar] [CrossRef]
- Snow, D.H.; Guy, P.S. Muscle fibre type composition of a number of limb muscles in different types of horse. Res. Vet. Sci. 1980, 54, 137–144. [Google Scholar] [CrossRef]
- Hyytiäinen, H.K.; Mykkänen, A.K.; Hielm-Björkman, A.K.; Stubbs, N.C.; McGowan, C.M. Muscle fiber type distribution of the thoracolumbar and hindlimb regions of horses: Relating fibre type and functional role. Acta Vet. Scand. 2014, 56, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Runx2, a multifunctional transcription factor in skeletal development. J. Cell. Biochem. 2002, 87, 1–8. [Google Scholar] [CrossRef]
- Chen, H.; Ghori-Javed, F.Y.; Rashid, H.; Adhami, M.D.; Serra, R.; Gutierrez, S.E.; Javed, A. Runx2 regulates endochondral ossification through control of chondrocyte proliferation and differentiation. J. Bone Miner. Res. 2014, 29, 2653–2665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffcott, L.B.; Henson, F.M.D. Studies on growth cartilage in the horse and their application to aetiopathogenesis of dyschondroplasia (osteochondrosis). Vet. J. 1998, 156, 177–192. [Google Scholar] [CrossRef]
- Distl, O. Genomics of skeletal disorders. In Equine Genomics, 1st ed.; Chowdhary, B.P., Ed.; Texas A&M University: College Station, TX, USA, 2013; pp. 187–198. [Google Scholar]
- Thomas, H.S. The Horse Conformation Handbook, 1st ed.; Storey Publishing: North Adams, MA, USA, 2005. [Google Scholar]
- Hill, E.W.; McGivney, B.A.; Gu, J.; Whiston, R.; MacHugh, D.E. A genome-wide SNP-association study confirms a sequence variant (g.66493737C>T) in the equine myostatin (MSTN) gene as the most powerful predictor of optimum racing distance for Thoroughbred racehorses. BMC Genom. 2010, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosio, Y.; Berto, G.; Camera, P.; Bianchi, F.; Ambrogio, C.; Claus, P.; Di Cunto, F. PPP4R2 regulates reuronal cell differentiation and survival, functionally cooperating with SMN. Eur. J. Cell Biol. 2012, 91, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Sewduth, R.N.; Jaspard-Vinassa, B.; Peghaire, C.; Guillabert, A.; Franzl, N.; Larrieu-Lahargue, F.; Moreau, C.; Fruttiger, M.; Dufourcq, P.; Couffinhal, T.; et al. The ubiquin ligase PDZRN3 is required for vascular morphogenesis through Wnt/planar cell polarity signaling. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.-A.; Kimura, Y.; Matsuura, K.; Yamamoto, H.; Gondo, T.; Inui, M. PDZRN3 (LNK3, SEMCAP3) is required for the differentiation of C2C12 myoblasts into myotubes. J. Cell Sci. 2006, 119, 5106–5113. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.F.; Shim, S.-O.; Strittmatter, S.M. Release of MICAL autoinhibition by Semaphorin-plexin signaling promotes interaction with collapsin response mediator protein. J. Neurosci. 2008, 28, 2287–2297. [Google Scholar] [CrossRef]
- Hung, R.-J.; Yazdani, U.; Yoon, J.; Wu, H.; Yang, T.; Gupta, N.; Huang, Z.; van Berkel, W.J.H.; Terman, J.R. Mical links semaphorins to F-actin disassembly. Nature 2010, 463, 823–827. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, R.; Holmes, K.C. Actin structure and function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef] [Green Version]
- Giridharan, S.S.P.; Rohn, J.L.; Naslavsky, N.; Caplan, S. Differential regulation of actin microfilaments by human MICAL proteins. J. Cell Sci. 2012, 125, 614–624. [Google Scholar] [CrossRef] [Green Version]
- Grozdanov, P.N.; Roy, S.; Kittur, N.; Meier, T. SHQ1 is required prior to NAF1 for assembly of H/ACA small nucleolar and telomerase RNPs. RNA J. 2009, 15, 1188–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, M.S.; Carlson, M.E.; Conboy, I.M. Differentiation rather than aging of muscle stem cells abolishes their telomerase activity. Biotechnol. Prog. 2009, 25, 1130–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleischer, J.; Breer, H.; Strotmann, J. Mammalian olfactory receptors. Front. Cell. Neurosci. 2009, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drutel, G.; Arranj, J.M.; Wisnewsky, C.; Schwartz, K.; Schwartz, J.C. Cloning of OL1, a putative olfactory receptor and its expression in the developing rat heart. Recept. Channels 1995, 3, 33–40. [Google Scholar] [PubMed]
- Feingold, E.A.; Penny, L.A.; Nienhuis, A.W.; Forget, B.G. An olfactory receptor gene is located in the extended human β-globin gene cluster and is expressed in erythroid cells. Genomics 1999, 61, 15–23. [Google Scholar] [CrossRef]
- Weber, M.; Pehl, U.; Breer, H.; Strotmann, J. Olfactory receptor expressed in ganglia of the autonomic nervous system. J. Neurosci. Res. 2002, 68, 176–184. [Google Scholar] [CrossRef]
- Griffin, C.A.; Kafadar, K.A.; Pavlath, G.K. MOR23 promotes muscle regeneration and regulates cell adhesion and migration. Dev. Cell 2009, 17, 649–661. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Article (Publication Year) | Group | Trait | Material | Breed | Country |
|---|---|---|---|---|---|
| Meira et al. (2014a) [14] | 1 | Morphometric traits (weight, rump length and body length) | Blood | Quarter horse | Brazil |
| Meira et al. (2014b) [15] | 2 | Speed index (SI) | Blood | Quarter horse | Brazil |
| Shin et al. (2015) [16] | 3 | Estimated breeding value (EBV) of race time | Blood | Thoroughbred | Korea |
| Ricard et al. (2017) [17] | 4 | Total race distance, average race speed and finishing status (qualified, eliminated or retired) | Blood | Arabian and crossed Arabian | France |
| Velie et al. (2018) [18] | 5 | Harness racing success (career earnings, best km time (s) and number of gallops) | Hair and/or blood | Norwegian-Swedish coldblood trotter | Sweden |
| Pereira et al. (2018) [19] | 6 | Maximum speed index | Blood | Quarter Horse | Brazil |
| McGivney et al. (2019) [20] | 7 | Racecourse starts, durability traits | Hair and/or blood | Thoroughbred | 16 countries |
| Gmel et al. (2019) [21] | 8 | Joint angles (poll, elbow, carpal, fetlock (front and hind), hip, stifle and hock) | Blood and/or sperm | Franches-Montagnes and Lipizzan | Switzerland, Austria, Croatia, Slovakia, and Hungary |
| Bussiman et al. (2020) [22] | 9 | Gait type | Blood | Mangalarga Marchador | Brazil |
| TF | Group | Biological Process | Literature Review * |
|---|---|---|---|
| HOXA5 | 1 and 2 | Respiratory system process and cartilage morphogenesis | Lung regulatory signaling pathways and tracheal cartilage patterning [23] |
| HIF1A | 1 and 8 | Response to oxidative stress and response to hypoxia | Skeletal muscle’s response to endurance training [24] |
| ARNT | 1, 4, 5, 6 and 8 | Positive regulation of glucose metabolic process and regulation of vascular endothelial growth factor production | Role in gluconeogenesis and lipogenic gene expression [25] |
| TFAP2A | 1, 2, 6, and 9 | Skeletal system development and regulation of bone mineralization | Regulation of cartilage and skeletal development [26] |
| SP1 | 1 and 6 | Definitive hemopoiesis | Muscle cell differentiation [27] |
| EGR1 | 1, 6 and 9 | Cellular response to fatty acids and cellular response to lipids | Regulation of cholesterol biosynthetic gene expression [28] |
| HAND1 | 2 | Cardiac ventricle development and cardiac chamber development | Cardiomyocyte proliferation and cardiac development [29] |
| E2F1 | 2 | Response to lipids and to fatty acids | Muscle oxidative metabolism [30] |
| STAT3 | 3 | Response to cytokine stimulus and inflammatory response | Muscle hypertrophy following resistance exercise [31] |
| RELA | 3 | Regulation of cartilage development and negative regulation of protein catabolic process | Transcriptional regulation during skeletal growth and osteoarthritis development [32] |
| SOX9 | 5 | Cartilage development and regulation of skeletal muscle fiber development | Regulation of chondrogenesis [33] |
| SOX17 | 5 | Cardiac endothelium cell differentiation and heart formation | Specification of endocardium cells and heart development [34] |
| SRF | 5 | Regulation of muscle contraction and regulation of muscle system process | Skeletal muscle hypertrophy [35] |
| FOXC1 | 6 | Cardiac muscle cell proliferation and heart growth | Morphogenesis of the cardiac outflow tract [36] |
| SOX2 | 8 | Lung morphogenesis and detection of mechanical stimulus | Branching morphogenesis and epithelial cell differentiation [37] |
| RXRA | 9 | Cardiac muscle tissue growth and heart growth | Myocardial growth and coronary artery formation [38] |
| Article (Publication Year) | Group | TF | Gene |
|---|---|---|---|
| Meira et al. (2014a) [14] | 1 | ARNT, TFAP2A, SP1, EGR1, HIF1A | HP1BP3, SH2D5, IL10RB |
| ARNT, TFAP2A, SP1, EGR1, HIF1A, HOXA5 | LPAR2 | ||
| TFAP2A, SP1, EGR1, HOXA5 | IFNAR1 | ||
| Meira et al. (2014b) [15] | 2 | TFAP2A, E2F1, HOXA5, HAND1 | GRM8, CCDC148, KAT6A |
| Shin et al. (2015) [16] | 3 | STAT3, RELA | PDZRN3, PPP4R2 |
| Ricard et al. (2017) [17] | 4 | ARNT | - |
| Velie et al. (2018) [18] | 5 | ARNT, NKX2-5, SOX9, SRF | STRBP |
| ARNT, NKX2-5 | RYBP | ||
| ARNT, SOX17 | PROK2 | ||
| NKX2-5, SOX9, SOX17, SRF | LOC100071438 | ||
| Pereira et al. (2018) [19] | 6 | ARNT, TFAP2A, SP1, FOXC1 | HDHD3 |
| ARNT, TFAP2A, EGR1, FOXC1 | ZBTB18 | ||
| ARNT, TFAP2A, SP1, EGR1, FOXC1 | ACACA | ||
| ARNT, TFAP2A, SP1, EGR1 | EPB41L3, ARMC2, CEP57L1, PPIL6, CYTH1, GEN1, SMC6, SHQ1, RYBP, PROK2, DIPK2, ODC1, HPCAL1, KLF11, GRHL1, PPP4R2, PRPF4, ALAD, POLE3 | ||
| ARNT | GGNBP2, PDZRN3 | ||
| Pereira et al. (2018) [19] | 6 | ARNT, TFAP2A, SP1, EGR1 | COL27A1, DCK, SQLE, DESI2, ADSS2, HNRNPU, METTL24, TIMP2, TASP1, LOC100056127, KCNS3, RUFY3, LGAL53BP, AMBP |
| ARNT, TFAP2A, SP1, EGR1, FOXC1 | COX20, WASHC5 | ||
| TFAP2A, SP1, EGR1, FOXC1 | MICAL1, SMC3 | ||
| FOXC1 | CANT1 | ||
| McGivney et al. (2019) [20] | 7 | NKX2-5 | - |
| Gmel et al. (2019) [21] | 8 | ARNT, HIF1A, SOX2 | - |
| Bussiman et al. (2020) [22] | 9 | TFAP2A, EGR1, RXRA | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Littiere, T.O.; Castro, G.H.F.; Rodriguez, M.d.P.R.; Bonafé, C.M.; Magalhães, A.F.B.; Faleiros, R.R.; Vieira, J.I.G.; Santos, C.G.; Verardo, L.L. Identification and Functional Annotation of Genes Related to Horses’ Performance: From GWAS to Post-GWAS. Animals 2020, 10, 1173. https://doi.org/10.3390/ani10071173
Littiere TO, Castro GHF, Rodriguez MdPR, Bonafé CM, Magalhães AFB, Faleiros RR, Vieira JIG, Santos CG, Verardo LL. Identification and Functional Annotation of Genes Related to Horses’ Performance: From GWAS to Post-GWAS. Animals. 2020; 10(7):1173. https://doi.org/10.3390/ani10071173
Chicago/Turabian StyleLittiere, Thayssa O., Gustavo H. F. Castro, Maria del Pilar R. Rodriguez, Cristina M. Bonafé, Ana F. B. Magalhães, Rafael R. Faleiros, João I. G. Vieira, Cassiane G. Santos, and Lucas L. Verardo. 2020. "Identification and Functional Annotation of Genes Related to Horses’ Performance: From GWAS to Post-GWAS" Animals 10, no. 7: 1173. https://doi.org/10.3390/ani10071173