May Salivary Chromogranin A Act as a Physiological Index of Stress in Transported Donkeys? A Pilot Study

 , , , and
, , , and 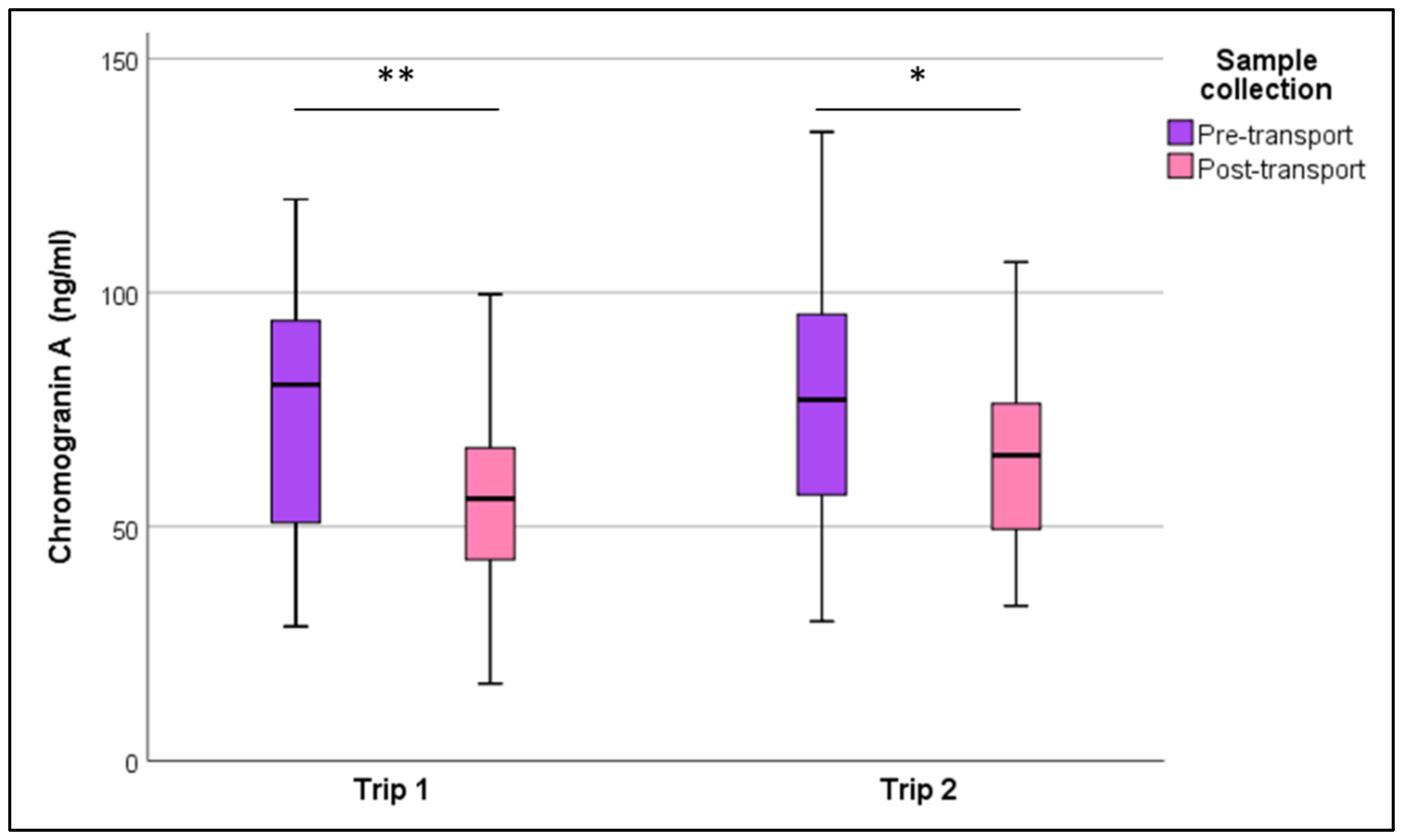{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations, Statistics Division. Forestry Production and Trade. Available online: http://www.fao.org/faostat/en/#data/FO (accessed on 4 April 2019).
- Dai, F.; Costa, E.D.; Murray, L.M.A.; Canali, E.; Minero, M. Welfare Conditions of Donkeys in Europe: Initial Outcomes from On-Farm Assessment. Animals 2016, 6, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrales-Hernández, A.; Mota-Rojas, D.; Guerrero-Legarreta, I.; Roldan-Santiago, P.; Rodríguez-Salinas, S.; Yáñez-Pizaña, A.; De La Cruz, L.; Gonzalez-Lozano, M.; Mora-Medina, P. Physiological responses in horses, donkeys and mules sold at livestock markets. Int. J. Vet. Sci. Med. 2018, 6, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Weeks, C.; McGreevy, P.; Waran, N.K. Welfare issues related to transport and handling of both trained and unhandled horses and ponies. Equine Vet. Educ. 2011, 24, 423–430. [Google Scholar] [CrossRef]
- Stull, C.; Rodiek, A.V. Physiological responses of horses to 24 hours of transportation using a commercial van during summer conditions. J. Anim. Sci. 2000, 78, 1458–1466. [Google Scholar] [CrossRef]
- Fazio, E.; Medica, P.; Cravana, C.; Ferlazzo, A. Comparative Exploration of the β-endorphin, Adrenocorticotrophic Hormone, and Cortisol Patterns’ Response to Simulated and Conventional Transportations of Stallions (Equus caballus). J. Equine Vet. Sci. 2016, 44, 74–81. [Google Scholar] [CrossRef]
- Padalino, B.; Hall, E.; Raidal, S.L.; Celi, P.; Knight, P.; Jeffcott, L.; Muscatello, G. Health Problems and Risk Factors Associated with Long Haul Transport of Horses in Australia. Animals 2015, 5, 1296–1310. [Google Scholar] [CrossRef]
- Fazio, E.; Prachi, S.; David, S.; Elizabeth, H. Total and free iodothyronine changes in response to transport of Equidae (Equus asinus and Equus caballus). Vet. Ital. 2017, 53, 55–60. [Google Scholar] [CrossRef]
- Fazio, E.; Ferlazzo, A. Evaluation of stress during transport. Vet. Res. Commun. 2003, 27, 519–524. [Google Scholar] [CrossRef]
- Fazio, E.; Medica, P.; Cravana, C.; Aveni, F.; Ferlazzo, A. Comparative endocrinological responses to short transportation of Equidae (Equus asinus and Equus caballus). Anim. Sci. J. 2012, 84, 258–263. [Google Scholar] [CrossRef]
- Minka, N.S.; Ayo, J.O. Physiological responses of food animals to road transportation stress. Afr. J. Biotechnol. 2009, 8, 7415–7427. [Google Scholar]
- Bonelli, F.; Rota, A.; Aurich, C.; Ille, N.; Camillo, F.; Panzani, D.; Sgorbini, M. Determination of Salivary Cortisol in Donkey Stallions. J. Equine Vet. Sci. 2019, 77, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Lensen, R.C.M.M.; Moons, C.P.H.; Diederich, C. Physiological stress reactivity and recovery related to behavioral traits in dogs (Canis familiaris). PLoS ONE 2019, 14, e0222581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, B.; Dillon, E.; Mills, P.J.; Ziegler, M.G. Catecholamines in human saliva. Life Sci. 2001, 69, 87–99. [Google Scholar] [CrossRef]
- Forhead, A.J.; Smart, D.; Smith, R.; Dobson, H. Transport-Induced stress responses in fed and fasted donkeys. Res. Vet. Sci. 1995, 58, 144–151. [Google Scholar] [CrossRef]
- Lee, T.; Shimizu, T.; Iijima, M.; Obinata, K.; Yamashiro, Y.; Nagasawa, S. Evaluation of psychosomatic stress in children by measuring salivary chromogranin A. Acta Paediatr. 2006, 95, 935–939. [Google Scholar] [CrossRef]
- Kanai, K.; Hino, M.; Hori, Y.; Nakao, R.; Hoshi, F.; Itoh, N.; Higuchi, S. Circadian variations in salivary chromogranin a concentrations during a 24-Hour period in dogs. J. Vet. Sci. 2008, 9, 421–423. [Google Scholar] [CrossRef] [Green Version]
- Akiyoshi, H.; Aoki, M.; Shimada, T.; Noda, K.; Kumagai, D.; Saleh, N.; Sugii, S.; Ohashi, F. Measurement of plasma chromogranin A concentrations for assessment of stress responses in dogs with insulin-Induced hypoglycemia. Am. J. Vet. Res. 2005, 66, 1830–1835. [Google Scholar] [CrossRef]
- Nakane, H.; Asami, O.; Yamada, Y.; Harada, T.; Matsui, N.; Kanno, T.; Yanaihara, N. Salivary Chromogranin A As An Index Of Psychosomatic Stress Response. Biomed. Res. 1998, 19, 401–406. [Google Scholar] [CrossRef] [Green Version]
- Casal i Plana, N. Identification of New Physiological Parameters for Monitoring Chronic Stress in Growing Pigs: Hair Cortisol and Chromogranin A. Ph.D. Thesis, Universitat Autònoma de Barcelona, Barcelona, Spain, 2016. [Google Scholar]
- Sato, F.; Ishida, N.; Hasegawa, T.; Katayama, Y.; Kanno, T.; Yanaihara, N.; Ohki, H.; Iwanaga, T.; Mukoyama, H. Expression of Equine Chromogranin A. In Chromogranins: Functional and Clinical Aspects; Helle, K.B., Aunis, D., Eds.; Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 2002; pp. 137–142. [Google Scholar]
- Sato, F.; Hasegawa, T.; Katayama, Y.; Iwanaga, T.; Yanaihara, N.; Kanno, T.; Ishida, N. Molecular Cloning of Equine Chromogranin A and Its Expression in Endocrine and Exocrine Tissues. J. Vet. Med. Sci. 2000, 62, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Tateo, A.; Padalino, B.; Boccaccio, M.; Maggiolino, A.; Centoducati, P. Transport stress in horses: Effects of two different distances. J. Vet. Behav. 2012, 7, 33–42. [Google Scholar] [CrossRef]
- Fazio, E.; Medica, P.; Cravana, C.; Ferlazzo, A. Pituitary-Adrenocortical adjustments to transport stress in horses with previous different handling and transport conditions. Vet. World 2016, 9, 856–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escribano, D.; Soler, L.; Montes, A.; Martínez-Subiela, S.; Cerón, J. Measurement of chromogranin A in porcine saliva: Validation of a time-Resolved immunofluorometric assay and evaluation of its application as a marker of acute stress. Animal 2012, 7, 640–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, S.; Soler, L.; Moons, C.; Kashiha, M.; Bähr, C.; Vandermeulen, J.; Janssens, S.; Montes, A.; Escribano, D.; Cerón, J.; et al. Different stressors elicit different responses in the salivary biomarkers cortisol, haptoglobin, and chromogranin A in pigs. Res. Vet. Sci. 2014, 97, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Escribano, D.; Montes, A.; Fuentes-Rubio, M.; Cerón, J. Saliva chromogranin A in growing pigs: A study of circadian patterns during daytime and stability under different storage conditions. Vet. J. 2014, 199, 355–359. [Google Scholar] [CrossRef]
- Escribano, D.; Ko, H.-L.; Chong, Q.; Llonch, L.; Manteca, X.; Llonch, P. Salivary biomarkers to monitor stress due to aggression after weaning in piglets. Res. Vet. Sci. 2019, 123, 178–183. [Google Scholar] [CrossRef]
- Fujimoto, S.; Nomura, M.; Niki, M.; Motoba, H.; Ieishi, K.; Mori, T.; Ikefuji, H.; Ito, S. Evaluation of stress reactions during upper gastrointestinal endoscopy in elderly patients: Assessment of mental stress using chromogranin A. J. Med. Investig. 2007, 54, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Yamakoshi, T.; Park, S.-B.; Jang, W.-C.; Kim, K.; Yamakoshi, Y.; Hirose, H. Relationship between salivary Chromogranin-A and stress induced by simulated monotonous driving. Med. Biol. Eng. 2009, 47, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Helle, K.B.; Corti, A.; Metz-Boutigue, M.-H.; Tota, B. The endocrine role for chromogranin A: A prohormone for peptides with regulatory properties. Cell. Mol. Life Sci. 2007, 64, 2863–2886. [Google Scholar] [CrossRef]
- Mahata, S.K. Catestatin—The Catecholamine Release Inhibitory Peptide: A Structural and Functional Overview. Available online: https://www.ingentaconnect.com/content/ben/cmciema/2004/00000004/00000003/art00010 (accessed on 15 April 2020).
- Mahapatra, N.R. Catestatin is a novel endogenous peptide that regulates cardiac function and blood pressure. Cardiovasc. Res. 2008, 80, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Dev, N.B.; Gayen, J.R.; O’Connor, D.T.; Mahata, S.K. Chromogranin a and the autonomic system: Decomposition of heart rate variability and rescue by its catestatin fragment. Endocrinology 2010, 151, 2760–2768. [Google Scholar] [CrossRef] [PubMed]
- Srithunyarat, T. Chromogranin A Epitopes Catestatin and Vasostatin. Ph.D. Thesis, Acta Universitatis Agriculturae Sueciae, Sveriges Lantbruksuniv, Uppsala, Sweden, 2017; pp. 1652–6880. [Google Scholar]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, F.; Dalla Costa, E.; Cannas, S.; Heinzl, E.U.L.; Minero, M.; Mazzola, S.M. May Salivary Chromogranin A Act as a Physiological Index of Stress in Transported Donkeys? A Pilot Study. Animals 2020, 10, 972. https://doi.org/10.3390/ani10060972
Dai F, Dalla Costa E, Cannas S, Heinzl EUL, Minero M, Mazzola SM. May Salivary Chromogranin A Act as a Physiological Index of Stress in Transported Donkeys? A Pilot Study. Animals. 2020; 10(6):972. https://doi.org/10.3390/ani10060972
Chicago/Turabian StyleDai, Francesca, Emanuela Dalla Costa, Simona Cannas, Eugenio Ugo Luigi Heinzl, Michela Minero, and Silvia Michela Mazzola. 2020. "May Salivary Chromogranin A Act as a Physiological Index of Stress in Transported Donkeys? A Pilot Study" Animals 10, no. 6: 972. https://doi.org/10.3390/ani10060972