Arginine Supply Impacts the Expression of Candidate microRNA Controlling Milk Casein Yield in Bovine Mammary Tissue

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Experiment
2.1.1. Experimental Design and Culture Medium
2.1.2. Cell Resuscitation and Treatment
2.1.3. Database Prediction and Result Acquisition
2.1.4. Detection of miRNA Expression by RT-qPCR
2.2. In Vivo Experiment
2.2.1. Experimental Animals and Management
2.2.2. Experimental Design
2.2.3. Samples and Data Collection
2.3. Statistical Analysis
3. Results
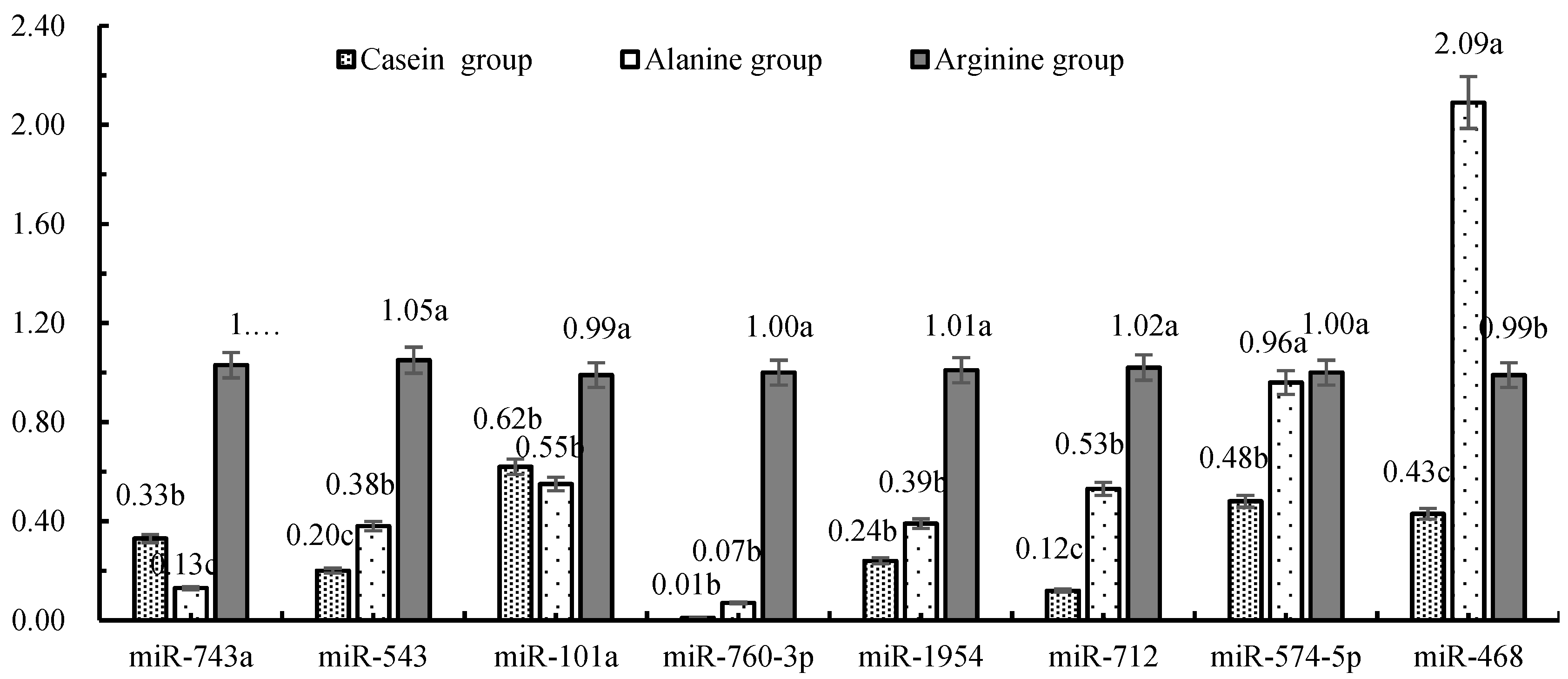
3.1. miRNA Expression in Mammary Cells In Vivo
3.2. Effects of Arginine Infusion on Milk Production in Lactating Cows
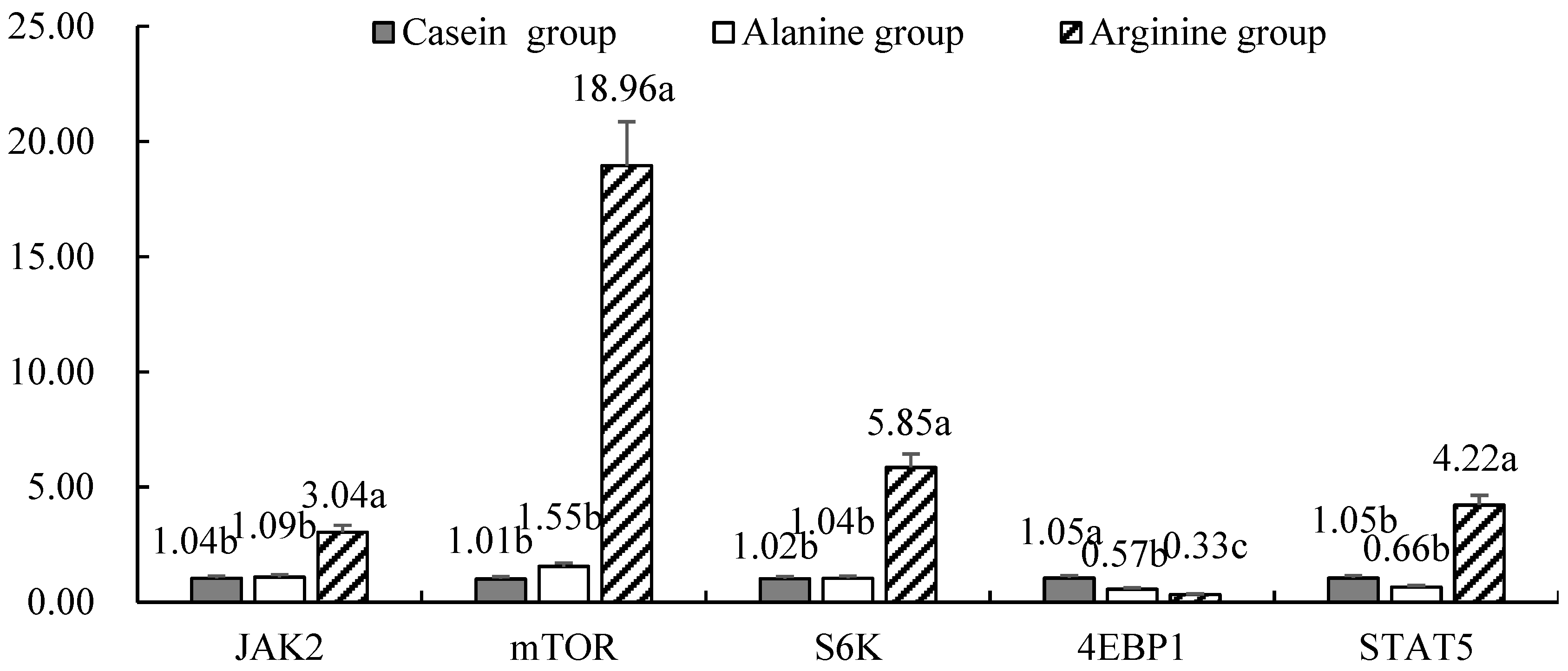
3.3. Effects of Arginine Infusion on Expression of mRNA and miRNA Involved in Milk Production
4. Discussion
4.1. Effects of Arginine Infusion on Milk Protein production
4.2. miRNA Regulation of Casein Gene Expression in Response to Arginine Supplementation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, S.W.; McPherson, R.L.; Wu, G. Dietary arginine supplementation enhances the growth of milk-fed young pigs. J. Nutr. 2004, 134, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Chew, B.P.; Eisenman, J.R.; Tanaka, T.S. Arginine infusion stimulates prolactin, growth hormone, insulin, and subsequent lactation in pregnant dairy cows. J. Dairy Sci. 1984, 67, 2507–2518. [Google Scholar] [CrossRef]
- Ma, Q.; Hu, S.; Bannai, M.; Wu, G. L-Arginine regulates protein turnover in porcine mammary epithelial cells to enhance milk protein synthesis. Amino Acids 2018, 50, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Wang, H.; Wu, T.; Ding, L.; Zhao, R.; Khas, E.; Wang, C.; Zhang, F.; Mi, F.; Wang, L.; et al. Milk protein responses to balanced amino acid and removal of Leucine and Arginine supplied from jugular-infused amino acid mixture in lactating dairy cows. J. Anim. Physiol. Anim. Nutr. (Berl.) 2017, 101, e278–e287. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Wu, G. Regulatory role for amino acids in mammary gland growth and milk synthesis. Amino Acids 2009, 37, 89–95. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Rhoads, J.; Satterfield, M.; Smith, S.B.; Spencer, T.E.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Xu, B.; Wang, H.; Bu, D.; Wang, J.; Loor, J.J. Effects of Arginine concentration on the in vitro expression of Casein and mTOR pathway related genes in mammary epithelial cells from dairy cattle. PLoS ONE 2014, 9, e95985. [Google Scholar] [CrossRef]
- Wang, C.; Li, Q. Identification of differentially expressed microRNAs during the development of Chinese murine mammary gland. J. Genet. Genom. 2007, 34, 966–973. [Google Scholar] [CrossRef]
- Wang, M.; Moisá, S.; Khan, M.J.; Wang, J.; Bu, D.; Loor, J.J. MicroRNA expression patterns in the bovine mammary gland are affected by stage of lactation 1. J. Dairy Sci. 2012, 95, 6529–6535. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Zhang, Y.; Wu, R.; Chen, X.; Zhang, Y.; Chen, X.; Zhu, D. miR-431 is involved in regulating cochlear function by targeting Eya4. Biochim. Biophys. Acta 2016, 1862, 2119. [Google Scholar] [CrossRef]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2005, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Haneda, S.; Imakawa, K.; Sakai, S.; Nagaoka, K. A microRNA, miR-101a, controls mammary gland development by regulating cyclooxygenase-2 expression. Differentiation 2009, 77, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Zidi, A.; Amills, M.; Tomás, A.; Vidal, O.; Ramírez, O.; Carrizosa, J.; Urrutia, B.; Serradilla, J.M.; Clop, A. Short communication: Genetic variability in the predicted microRNA target sites of caprine casein genes. J. Dairy Sci. 2010, 93, 1749–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.Z.; Ding, L.Y.; Wang, C.; Chen, L.M.; Loor, J.J.; Wang, H.R. Short communication: Arginase inhibition reduces the synthesis of casein in bovine mammary epithelial cells. J. Dairy Sci. 2017, 100, 4128–4133. [Google Scholar] [CrossRef] [PubMed]
- Vickery, H.B.; White, A. The basic amino acids of casein. J. Biol. Chem. 1933, 103, 413–415. [Google Scholar]
- Thomson, J.M.; Newman, M.; Parker, J.S.; Morinkensicki, E.M.; Wright, T.; Hammond, S.M. Extensive post-transcriptional regulation of microRNAs and its implications for cancer. Genes Dev. 2006, 20, 2202–2207. [Google Scholar] [CrossRef] [Green Version]
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; National Research Council, National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Bionaz, M.; Loor, J.J. Identification of reference genes for quantitative real-time PCR in the bovine mammary gland during the lactation cycle. Physiol. Genom. 2007, 29, 312–319. [Google Scholar] [CrossRef]
- Loor, J.J.; Dann, H.M.; Guretzky, N.A.; Everts, R.E.; Oliveira, R.; Green, C.A.; Litherland, N.B.; Rodriguez-Zas, S.L.; Lewin, H.A.; Drackley, J.K. Plane of nutrition prepartum alters hepatic gene expression and function in dairy cows as assessed by longitudinal transcript and metabolic profiling. Physiol. Genom. 2006, 27, 29–41. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△Ct method. Methods 2000, 25, 4. [Google Scholar]
- Hallén, E.; Wedholm, A.; Andrén, A.; Lundén, A. Effect of beta-casein, kappa-casein and beta-lactoglobulin genotypes on concentration of milk protein variants. J. Anim. Breed Genet. 2008, 125, 119–129. [Google Scholar] [CrossRef]
- Chen, L.M.; Li, Z.T.; Wang, M.Z.; Wang, H.R. Preliminary report of arginine on synthesis and gene expression of casein in bovine mammary epithelial cell. Int. Res. J. Agric. Sci. Soil Sci. 2013, 3, 17–23. [Google Scholar]
- Ding, L.Y.; Chen, L.M.; Wang, M.Z.; Zhang, J.; Loor, J.J.; Zhou, G.; Zhang, X.; Wang, H.R. Inhibition of arginase via jugular infusion of Nω-hydroxy-nor-l-arginine inhibits casein synthesis in lactating dairy cows. J. Dairy Sci. 2018, 101, 3514–3523. [Google Scholar] [CrossRef]
- Hu, L.Y.; Wu, J.C.; Wang, M.Z.; Li, P.F.; Feng, D.; Zhang, J. Effects of dietary arginine on mammary gland development and casein synthesis in Wistar rats after delivery. Chin. J. Anim. Nutr. 2015, 27, 1326–1332. [Google Scholar]
- Lorian, V. Differences between in vitro and in vivo studies. Antimicrob. Agents Chemother. 1988, 32, 1600–1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bionaz, M.; Hurley, W.; Loor, J. Milk protein synthesis in the lactating mammary gland: Insights from transcriptomics analyses. In Milk Proteinn; Hurley, W., Ed.; InTech North America: New York, NY, USA, 2012; pp. 285–324. [Google Scholar]
- Nichols, K.; Doelman, J.; Kim, J.J.M.; Carson, M.; Metcalf, J.A.; Cant, J.P. Exogenous essential amino acids stimulate an adaptive unfolded protein response in the mammary glands of lactating cows. J. Dairy Sci. 2017, 100, 5909–5921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cant, J.P.; Kim, J.J.M.; Cieslar, S.R.L.; Doelman, J. Symposium review: Amino acid uptake by the mammary glands: Where does the control lie? J. Dairy Sci. 2018, 101, 5655–5666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Q.; Gibson, G.E. Up-regulation of the mitochondrial malate dehydrogenase by oxidative stress is mediated by miR-743a. J. Neurochem. 2011, 118, 440–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, T.; Hebbel, R.P.; Kaul, D.K. Protective effect of arginine on oxidative stress in transgenic sickle mouse models. Free Radic. Biol. Med. 2006, 41, 1771–1780. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Yin, Y.; Kong, X.; Li, P.; Li, X.; Huang, R.; Wu, G. L-Arginine stimulates proliferation and prevents endotoxin-induced death of intestinal cells. Amino Acids 2010, 38, 1227–1235. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Dong, G.; Wang, B.; Gao, W.; Yang, Q. miR-543 promotes gastric cancer cell proliferation by targeting SIRT1. Biochem. Biophys. Res. Commun. 2016, 469, 15–21. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Du, Y.; Wang, L.; Liu, X.H.; Guo, J.; Weng, X.D. MiR-543 promotes cell proliferation and metastasis of renal cell carcinoma by targeting Dickkopf 1 through the Wnt/β-catenin signaling pathway. J. Cancer 2018, 9, 3660–3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Gan, L.; Zhang, J. miR-543 inhibites cervical cancer growth and metastasis by targeting TRPM7. Chem. Biol. Interact. 2019, 302, 83–92. [Google Scholar] [CrossRef]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; Ye, G.; Zhang, J.; Wang, L.; Wang, T.; Wang, Z.; Zhang, T.; Wang, G.; Guo, Z.; Luo, Y. Original article: miR-574-5p negatively regulates Qki6/7 to impact β-catenin/Wnt signalling and the development of colorectal cancer. Gut 2013, 62, 716–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Li, X.; Xiao, Q. MiR-574-5p mediates the cell cycle and apoptosis in thyroid cancer cells via Wnt/β-catenin signaling by repressing the expression of Quaking proteins. Oncol. Lett. 2018, 15, 5841–5848. [Google Scholar] [CrossRef]
- Wang, X.; Lu, X.; Geng, Z.; Yang, G.; Shi, Y. LncRNA PTCSC3/miR-574-5p Governs Cell Proliferation and Migration of Papillary Thyroid Carcinoma via Wnt/β-Catenin Signaling. J. Cell. Biochem. 2017, 118, 4745–4752. [Google Scholar] [CrossRef]
- Dong, J.S.; Kumar, S.; Takabe, W.; Chan, W.K.; Ni, C.W.; Albertsgrill, N.; Jang, I.H.; Kim, S.; Kim, W.; Kang, S.W. The atypical mechanosensitive microRNA-712 derived from pre-ribosomal RNA induces endothelial inflammation and atherosclerosis. Nat. Commun. 2013, 4, 3000. [Google Scholar]
- Kim, C.W.; Kumar, S.; Son, D.J.; Jang, I.H.; Griendling, K.K.; Jo, H. Prevention of abdominal aortic aneurysm by anti–microrna-712 or anti–microrna-205 in angiotensin ii–infused mice. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1412. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Amino Acids | Control Group | Casein Group | Arginine Group |
|---|---|---|---|
| Tyr | 3.25 | 3.25 | 3.25 |
| Ala | 2.76 | 2.76 | 2.76 |
| Gly | 0.57 | 0.57 | 0.57 |
| Glu | 15.94 | 15.94 | 15.94 |
| Ser | 5.09 | 5.09 | 5.09 |
| Cys | 0.35 | 0.35 | 0.35 |
| Phe | 3.57 | 3.57 | 3.57 |
| Leu | 11.75 | 11.75 | 11.75 |
| Ile | 4.24 | 4.24 | 4.24 |
| His | 1.38 | 1.38 | 1.38 |
| Lys | 4.39 | 4.39 | 4.39 |
| Thr | 4.14 | 4.14 | 4.14 |
| Met | 2.22 | 2.22 | 2.22 |
| Trp | 1.05 | 1.05 | 1.05 |
| Val | 4.84 | 4.84 | 4.84 |
| Pro | 7.53 | 7.53 | 7.53 |
| Asp | 3.38 | 3.38 | 3.38 |
| Arg | 0 | 1.60 | 3.20 |
| miRNAs | Target Gene Symbol | |||
|---|---|---|---|---|
| miR-325 | CSN1S1 | CSN3 | PRODH | ASL |
| miR-3062 | CSN1S1 | CSN3 | ||
| miR-471-3p | CSN1S1 | ARGⅡ | ||
| miR-186 | CSN1S1 | CSN3 | ||
| miR-465c-5p | CSN1S1 | OAT | ||
| miR-465b-5p | CSN1S1 | OAT | ||
| miR-465a-5p | CSN1S1 | OAT | ||
| miR-1942 | CSN1S1 | ARGⅡ | ||
| miR-743b-3p | CSN1S1 | OAT | ||
| miR-743a | CSN1S1 | OAT | ||
| miR-543 | CSN1S1 | ARGⅡ | OAT | |
| miR-362-3p | CSN1S1 | OAT | ||
| miR-329 | CSN1S1 | OAT | ||
| miR-694 | CSN1S1 | ARGⅠ | OAT | |
| miR-145 | CSN2 | |||
| miR-335-3p | CSN3 | ARG1 | ||
| miR-208a-5p | CSN3 | OAT | ||
| miR-3095-5p | CSN3 | OAT | ||
| miR-875-3p | CSN3 | OAT | ||
| miR-669b | CSN3 | ARG1 | ||
| miR-3071 | CSN3 | ARGⅡ | ||
| miR-574-5p | CSN3 | PRODH | ||
| miR-181d | CSN3 | OAT | ||
| miR-181a | CSN3 | OAT | ||
| miR-181b | CSN3 | OAT | ||
| miR-181c | CSN3 | OAT | ||
| miR-434-3p | CSN3 | ARGⅠ | ||
| miR-3067 | CSN3 | ARGⅡ | ||
| miR-374 | CSN3 | OAT | ||
| miR-384-3p | CSN3 | ARGⅠ | OAT | PRODH |
| miR-410 | CSN3 | OAT | ||
| miR-344d | CSN3 | OAT | ||
| miR-344e | CSN3 | OAT | ||
| miR-686 | CSN3 | ARGⅠ | ||
| miR-141 | CSN3 | GLS | ||
| miR-200a | CSN3 | GLS | ||
| miR-340-5p | CSN3 | ARGⅠ | ||
| miR-712 | CSN3 | ARGⅡ | ||
| miR-205 | CSN3 | ARGⅡ | ||
| miR-467f | CSN3 | ARGⅠ | ||
| miR-669m-3p | CSN3 | ARGⅠ | ||
| miR-409-3p | CSN3 | ARGⅡ | ||
| miR-101b | mTOR | ARGⅡ | ||
| miR-101a | mTOR | ARGⅡ | ||
| miR-466c-3p | ARGⅠ | OAT | ||
| miR-466p-3p | ARGⅠ | OAT | ||
| miR-466b-3p | ARGⅠ | OAT | ||
| miR-532-5p | ARGⅡ | OAT | ||
| miR-3065 | ARGⅡ | OAT | ||
| miR-5098 | ARGⅡ | OAT | ||
| miR-468 | ARGⅡ | OAT | ||
| miR-760-3p | OAT | PRODH | ||
| miR-1954 | OAT | PRODH | ||
| Ingredients | Percentage/% | Nutrients | Level/% |
|---|---|---|---|
| Alfalfa | 15.30 | NEL/(MJ/kg) 2) | 4.66 |
| Chinese wildrye | 10.47 | CP | 14.08 |
| Silage | 28.80 | NFC | 40.27 |
| Corn | 21.50 | NDF | 34.99 |
| Cottonseed meal | 6.10 | ADF | 21.09 |
| Soybean meal | 6.80 | EE | 3.96 |
| Distillers Dried Grains with Soluble | 9.40 | Ca | 0.91 |
| CaHPO4 | 0.60 | Total P | 0.59 |
| NaCl | 0.50 | ||
| Premix1) | 0.53 | ||
| Total | 100.00 |
| Amino Acids | Casein Group | Arginine Group | Alanine Group |
|---|---|---|---|
| Lys | 21.50 | 21.50 | 86.00 |
| Met | 13.92 | 13.92 | 13.92 |
| Phe | 12.88 | 12.88 | 12.88 |
| Ile | 4.75 | 4.75 | 4.75 |
| Arg | 0.00 | 9.42 | 0.00 |
| Ala | 0.00 | 0.00 | 19.31 |
| Gene | Accession No. | Gene Description | Sequence (5′ → 3′) | Sources | |
|---|---|---|---|---|---|
| CSN1S1 | BC109618 | αs1-casein | F | TCA CAG TAT GAA AGA GGG AA | Bos taurus |
| R | AGC CAA TAG GAT TAG GGA | ||||
| CSN1S2 | NM_174528.2 | αs2-casein | F | AGG AAC GCA AAT GAA GAG | Bos taurus |
| R | GGA GTA ATG GGA ACA GCA | ||||
| CSN2 | NM_181008 | β-casein | F | TGA GGA ACA GCA GCA AAC | Bos taurus |
| R | CAG AGG CAG AGG AAG GTG | ||||
| CSN3 | NM_174294 | κ-casein | F | CGC TGT GAG AAA GAT GAA | Bos taurus |
| R | AGA CCG CAG TTG AAG TAA | ||||
| mTOR | XM_001788228.1 | Mechanistic Target of Rapamycin | F | CAT GTG CGA ACA CAG CAA CA | Bos taurus |
| R | TGC CTT TCA CGT TCC TCT CC | ||||
| JAK2 | XM_002689603.1 | Janus kinase 2 | F | ACA GGG ATC TGG CAA CAA GG | Bos taurus |
| R | CGC ATA AAT TCC GCT GGT GG | ||||
| STAT5 | NM_001012673.1 | Signal transducer and activator of transcription 5 | F | CAA TGG ACA GTC TGG AGC CC | Bos taurus |
| R | CCT GCA CAC TGG GGA TTG TT | ||||
| S6K | NM_205816.1 | Ribosomal protein S6 kinase | F | CGG AAC AGT CAC ACA CAC CT | Bos taurus |
| R | TGG CTT CTT GCG TGA GGT AG | ||||
| 4EBP1 | BC120290.1 | EIF4E binding protein1 | F | CGG AAC TCA CCT GTG ACC AA | Bos taurus |
| R | AGG TGA TTC TGC CTG GCT TC | ||||
| GAPDH | XM_001252479.1 | Glyceraldehyde-3-phosphate dehydrogenase | F | CCC CGC GCT CTA ATG TTC A | Bos taurus |
| R | GAA GGG GTC ATT GAT GGC GA | ||||
| ACTB | NM_173979.3 | β-actin | F | ACT GTT AGC TGC GTT ACA CCC TT | Bos taurus |
| R | TGC TGT CAC CTT CAC CGT TCC | ||||
| miRNAs | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| Control | Casein | Arginine | |||
| miR-101b | 1.32 | 1.28 | 0.98 | 0.11 | 0.342 |
| miR-181b | 0.82 | 0.93 | 0.84 | 0.063 | 0.749 |
| miR-181d | 1.23 | 1.03 | 1.22 | 0.083 | 0.792 |
| miR-329 | 1.79 | 2.14 | 1.96 | 0.189 | 0.993 |
| miR-3062 | 2.27 | 1.74 | 2.47 | 0.17 | 0.463 |
| miR-3065 | 0.90 | 1.16 | 1.02 | 0.083 | 0.212 |
| miR-743a | 1.20b | 2.10ab | 2.73a | 0.213 | <0.001 |
| miR-543 | 0.68b | 0.61b | 1.60a | 0.103 | 0.012 |
| miR-101a | 0.22b | 0.37b | 3.99a | 0.157 | 0.003 |
| miR-760-3p | 1.76a | 0.82b | 2.35a | 0.127 | 0.034 |
| miR-1954 | 1.99b | 0.7c | 5.63a | 0.183 | <0.001 |
| miR-712 | 0.42b | 0.52b | 2.44a | 0.147 | 0.01 |
| miR-574-5p | 0.95b | 0.35b | 5.63a | 0.107 | <0.001 |
| miR-468 | 0.23b | 0.30b | 1.48a | 0.047 | 0.041 |
| miR-875-3p | 48.26a | 17.17b | 2.97c | 1.653 | 0.001 |
| Items | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| Casein | Alanine | Arginine | |||
| Average daily intake/(kg) | 22.82 | 21.64 | 22.3 | 1.625 | 0.461 |
| Milk protein/% | 3.04b | 3.11b | 3.17a | 0.077 | 0.046 |
| Milk yield/(kg/d) | 21.45b | 23.65a | 24.16a | 0.823 | 0.039 |
| Milk protein yield/(kg/d) | 0.65b | 0.74a | 0.77a | 0.027 | 0.04 |
| α-Casein/(g/L) | 11.70b | 16.37a | 17.24a | 1.417 | 0.008 |
| β-Casein/(g/L) | 8.56 | 7.27 | 9.09 | 0.523 | 0.985 |
| κ-Casein/(g/L) | 2.00b | 2.37b | 3.00a | 0.194 | 0.024 |
| Casein content/% | 2.23c | 2.60b | 2.93a | 0.167 | 0.031 |
| Casein yield/(kg/d) | 0.48c | 0.62b | 0.71a | 0.053 | 0.044 |
| Casein/milk protein/% | 73.18b | 83.85ab | 92.63a | 6.613 | 0.003 |
| αs1-casein | 0.96b | 1.00b | 1.54a | 0.113 | 0.028 |
| αs2-casein | 0.98b | 1.03b | 1.39a | 0.137 | 0.037 |
| β-casein | 1.03 | 1.43 | 1.22 | 0.117 | 0.832 |
| κ-casein | 1.00 | 1.03 | 1.36 | 0.103 | 0.327 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wang, Y.; Wang, M.; Zhou, G.; Chen, L.; Ding, L.; Bu, D.; Loor, J. Arginine Supply Impacts the Expression of Candidate microRNA Controlling Milk Casein Yield in Bovine Mammary Tissue. Animals 2020, 10, 797. https://doi.org/10.3390/ani10050797
Zhang X, Wang Y, Wang M, Zhou G, Chen L, Ding L, Bu D, Loor J. Arginine Supply Impacts the Expression of Candidate microRNA Controlling Milk Casein Yield in Bovine Mammary Tissue. Animals. 2020; 10(5):797. https://doi.org/10.3390/ani10050797
Chicago/Turabian StyleZhang, Xin, Yifan Wang, Mengzhi Wang, Gang Zhou, Lianmin Chen, Luoyang Ding, Dengpan Bu, and Juan Loor. 2020. "Arginine Supply Impacts the Expression of Candidate microRNA Controlling Milk Casein Yield in Bovine Mammary Tissue" Animals 10, no. 5: 797. https://doi.org/10.3390/ani10050797