This study shows that the in-vitro digestibility of the three fiber sources significantly increased in DM% compared to the control group. Cornstalk showed a higher value of digestibility compared to alfalfa and rice straw. Greater digestibility of corn stalk than rice straw and alfalfa in these results may be due to the lower crude fiber content in corn stalk compared to alfalfa and rice straw. Similarly, Park et al. [
26] observed that in-vitro digestibility significantly increased in corn and wheat treatments. The levels of fiber in pig’s diet were reviewed as an important element affecting digestibility, and fiber-containing diets have a potential to increase fecal output [
27]. In the current study, in-vitro digestibility results for both methods used did not show any effect on the control, while a positive correlation was observed between alfalfa and corn stalk treatment (
p < 0.01). However, there was a negative correlation between rice straw and cornstalk treatments.
A higher response was observed in all treatment groups in the intestinal digesta method in comparison to the 3-step procedure, which is likely due to the extent of feed residues that were available in the intestinal digesta. Similar results were observed by Chen et al. [
28] when the 2-step in-vitro gas production technique (2 + IVGPT) and 3 steps were used, the 2 + IVGPT method presented a significantly higher dietary fiber digestibility compared to the 3-step method. The 3-step method stimulated the hydrolysis of intestinal microbes, which was induced by fibrolytic enzymes mainly and not from fermentation [
28,
29]. In-vitro fermentation of different fiber treatments resulted in the production of short-chain fatty acids (SCFA), including acetic, propionic, and n-butyric acids that were measured. Rice straw showed a higher concentration of acetic, propionic, and n- butyric acids compared to alfalfa and cornstalk treatments and control. Similarly, Fernando et al. [
30] suggested that all probiotic strains on rice fiber formed SCFA, especially acetate, at twice the level of the untreated control. Several studies have demonstrated that different fiber sources are fermented at different rates and produce different amounts of SCFA [
31]. Our results show that the fermentation rates are clearly dependent on the fiber source; hence, the concentration rates and peak areas differ. Acetic acid concentration and peak area were significantly higher compared to other SCFA; however, there no significant difference detected between different treatments The higher concentration of SCFA in dietary fiber shows that the substrate in rice straw contains more fermentable material than other treatments [
30]. In the current study, it was observed that peak areas were significantly higher in the controls than treatment groups in all measured SCFA. The alfalfa treatment demonstrated a significant decrease in SCFA concentrations and peak areas in contrast to findings by Liu et al. [
32], who indicated that alfalfa meal treatment could produce more SCFA and improve piglet gut health, which was associated with less diarrhea. This may be dependent on the age of the pig and the level of feeding. All treatments and untreated controls showed less effect on n-butyric concentrations and peak areas. Previously, butyric acid had a high response on organic acids when used as feed additives. Gut microbiota contains numerous and diverse microbial populations [
33], and in the past, the identification of porcine microbiota has been made using culture-dependent techniques.
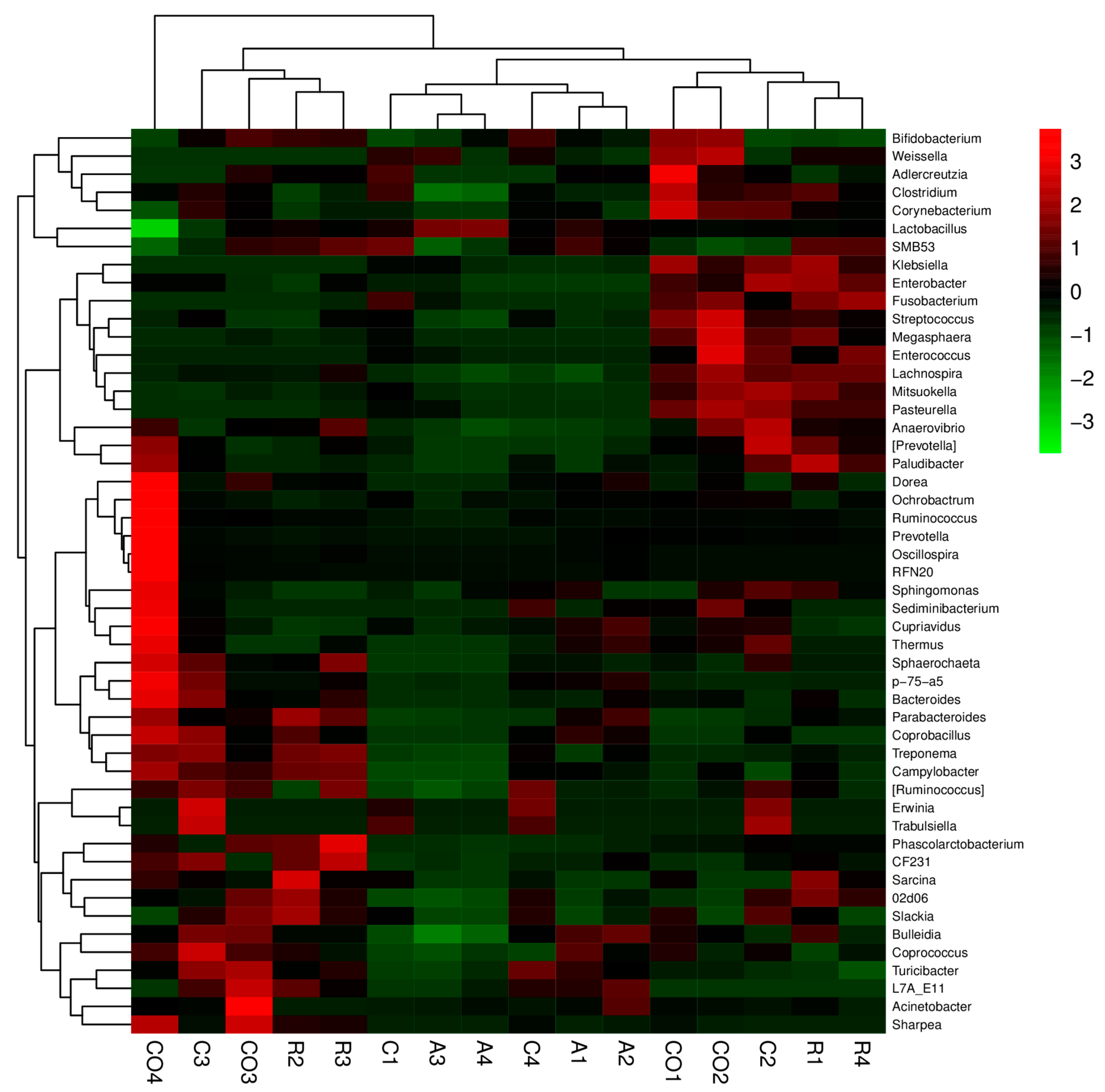
However, the knowledge of microbiota has advanced over the years with the advent of next-generation sequencing technology and bioinformatics [
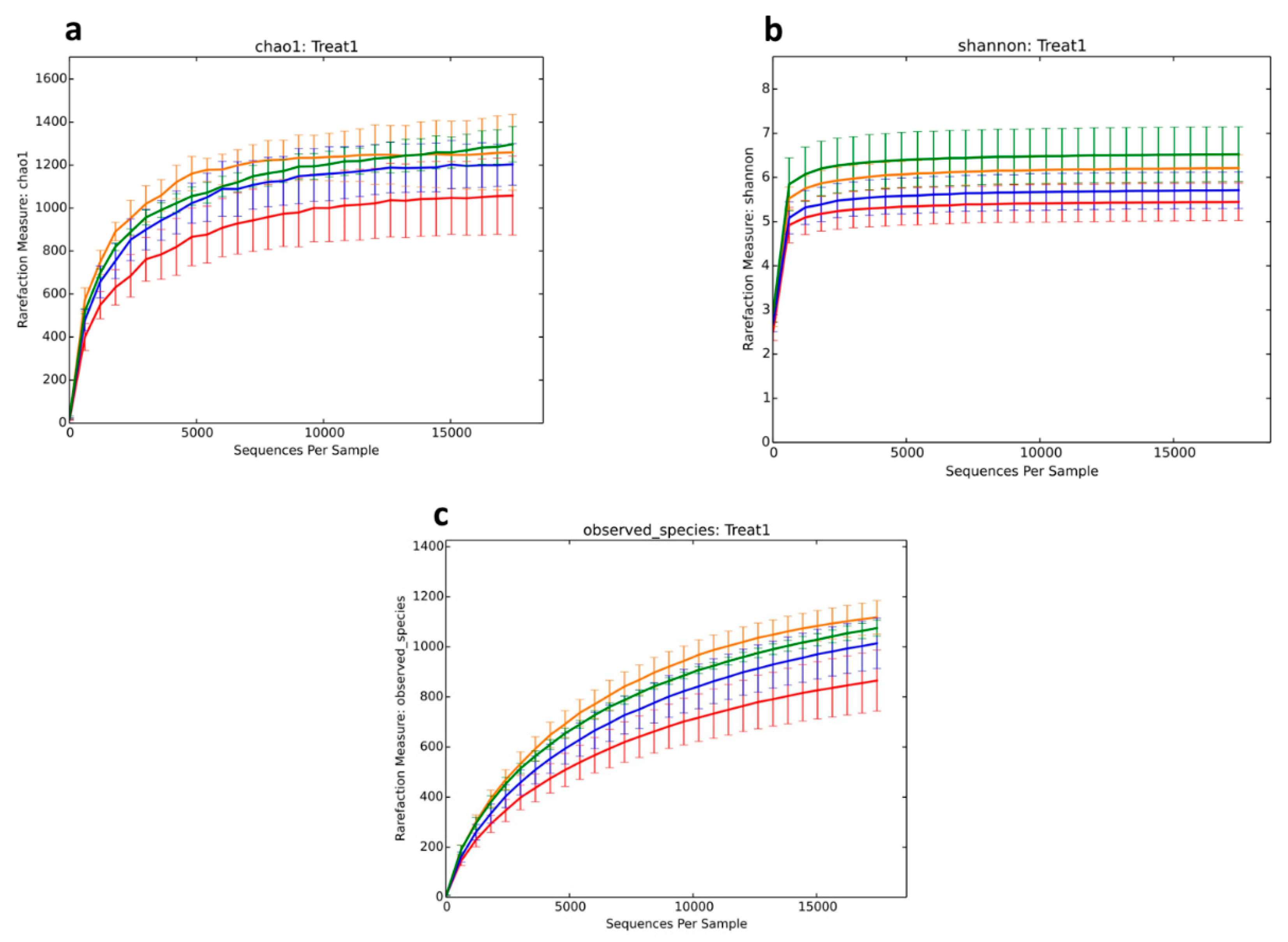
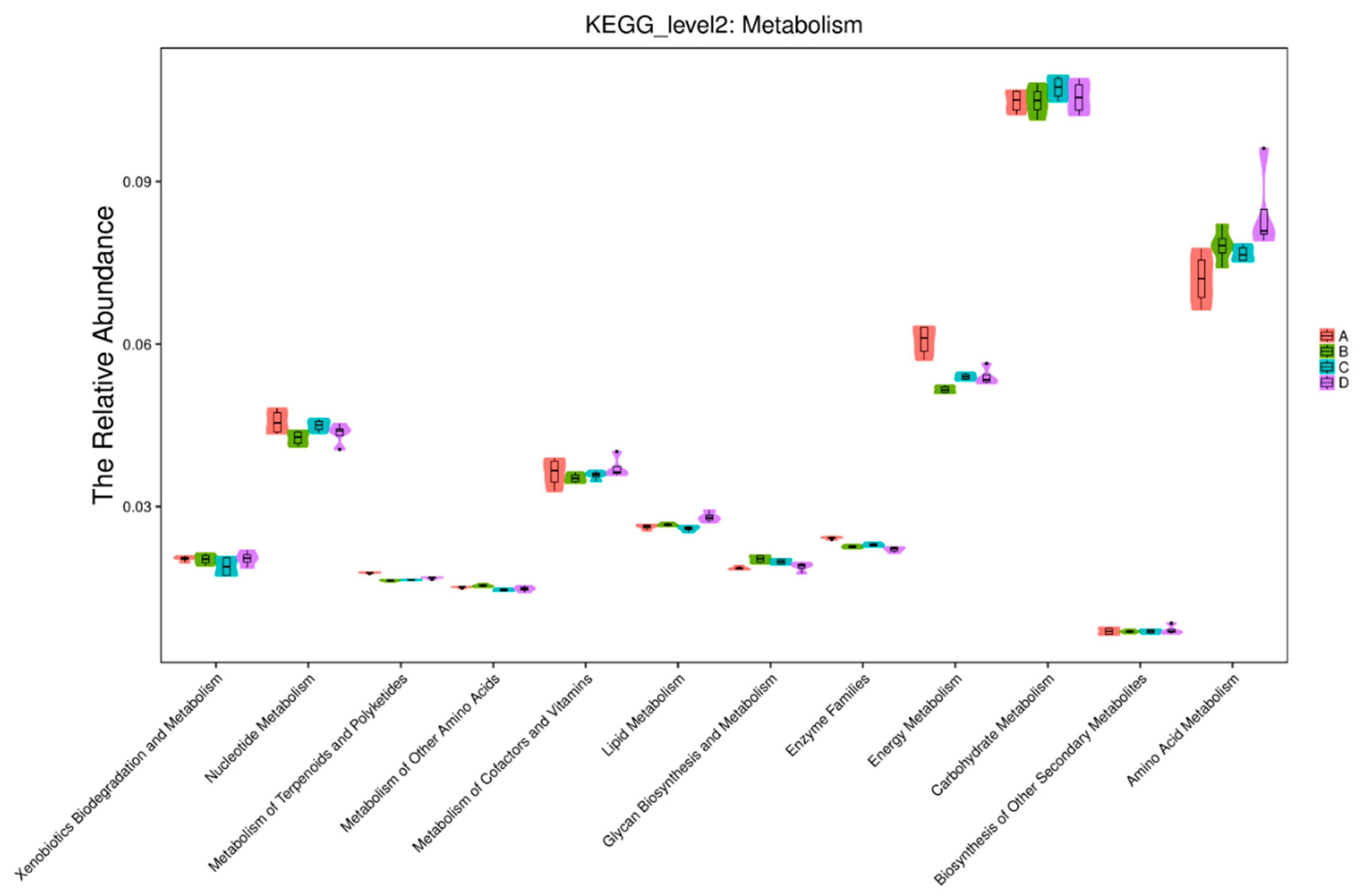
34]. The effect of dietary fibers on bacterial composition during fermentation was investigated using 16S rRNA based methods and illumina MiSeq sequencing. OTU analysis in this study revealed microbial diversity in cecum digesta in all treatment groups. We found that alpha diversity indices differed based on treatments in terms of species richness and evenness. The Chao1 index showed that there was less microbial diversity among treatments compared to controls, whereas microbial richness and species diversity increased among the treatments, as shown by the Shannon diversity index. In agreement with Yu et al. [
35], who showed similar results in the cecum microbial population with chitosen supplementation. Results of the speccacum species curve in
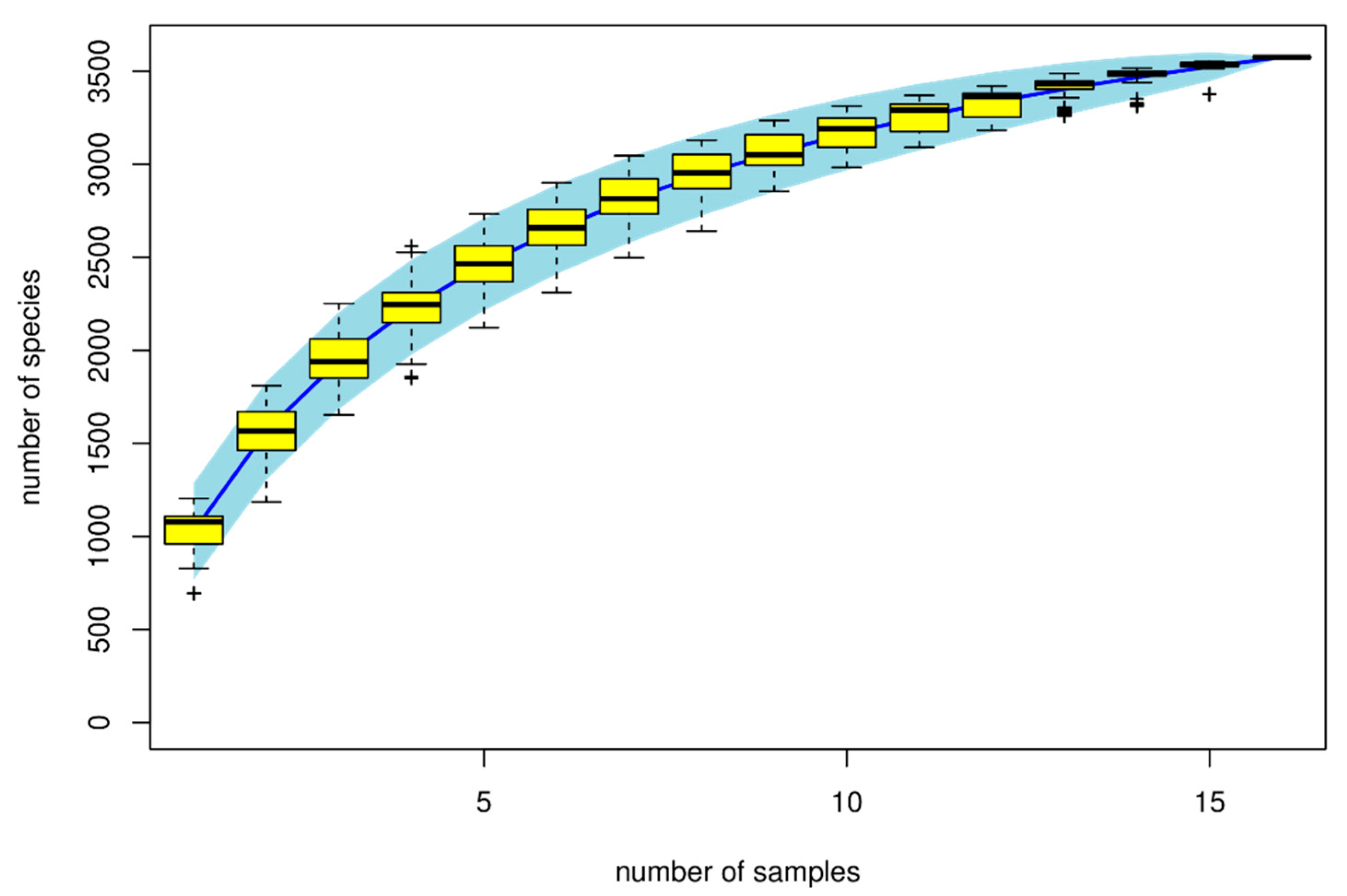
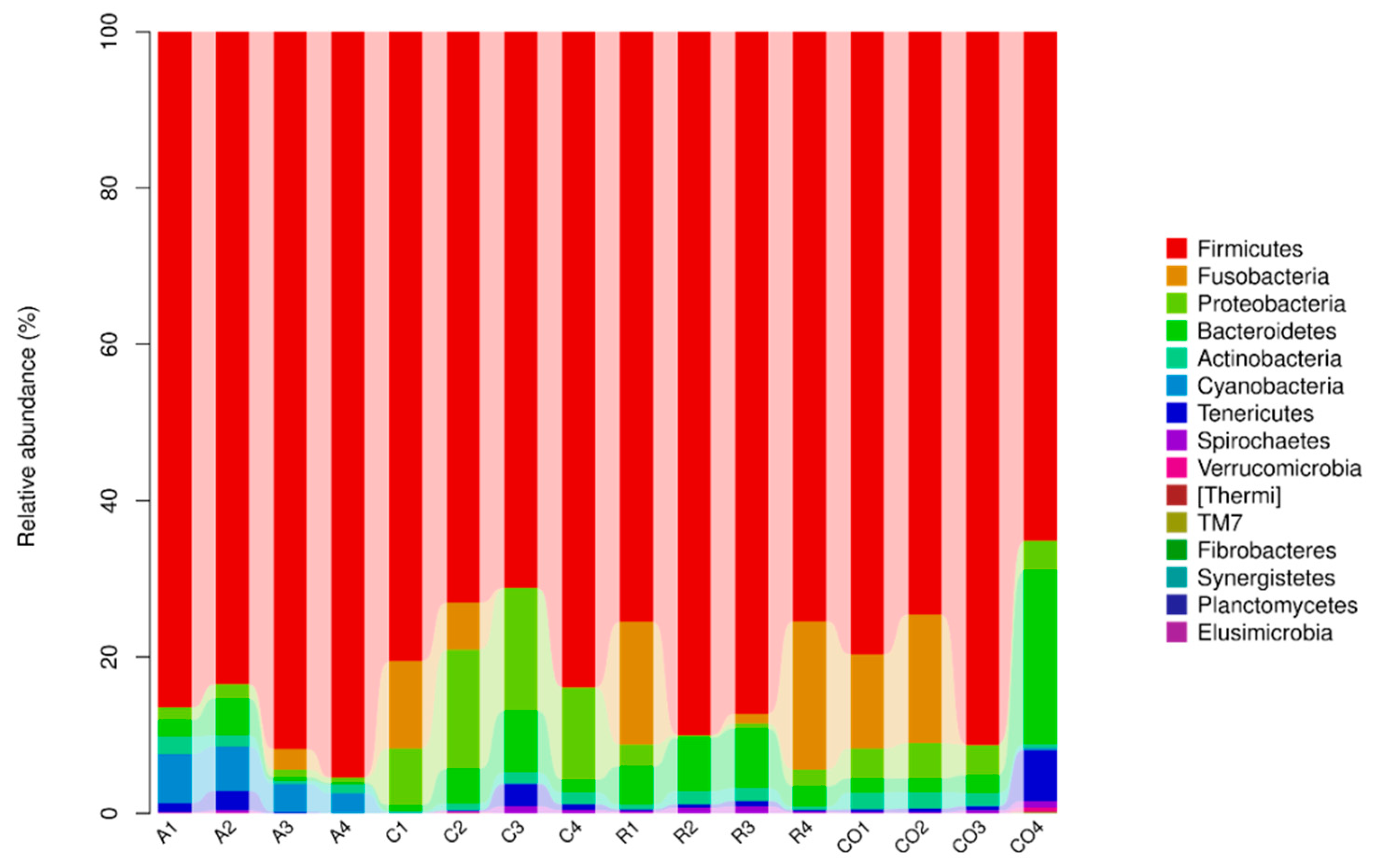
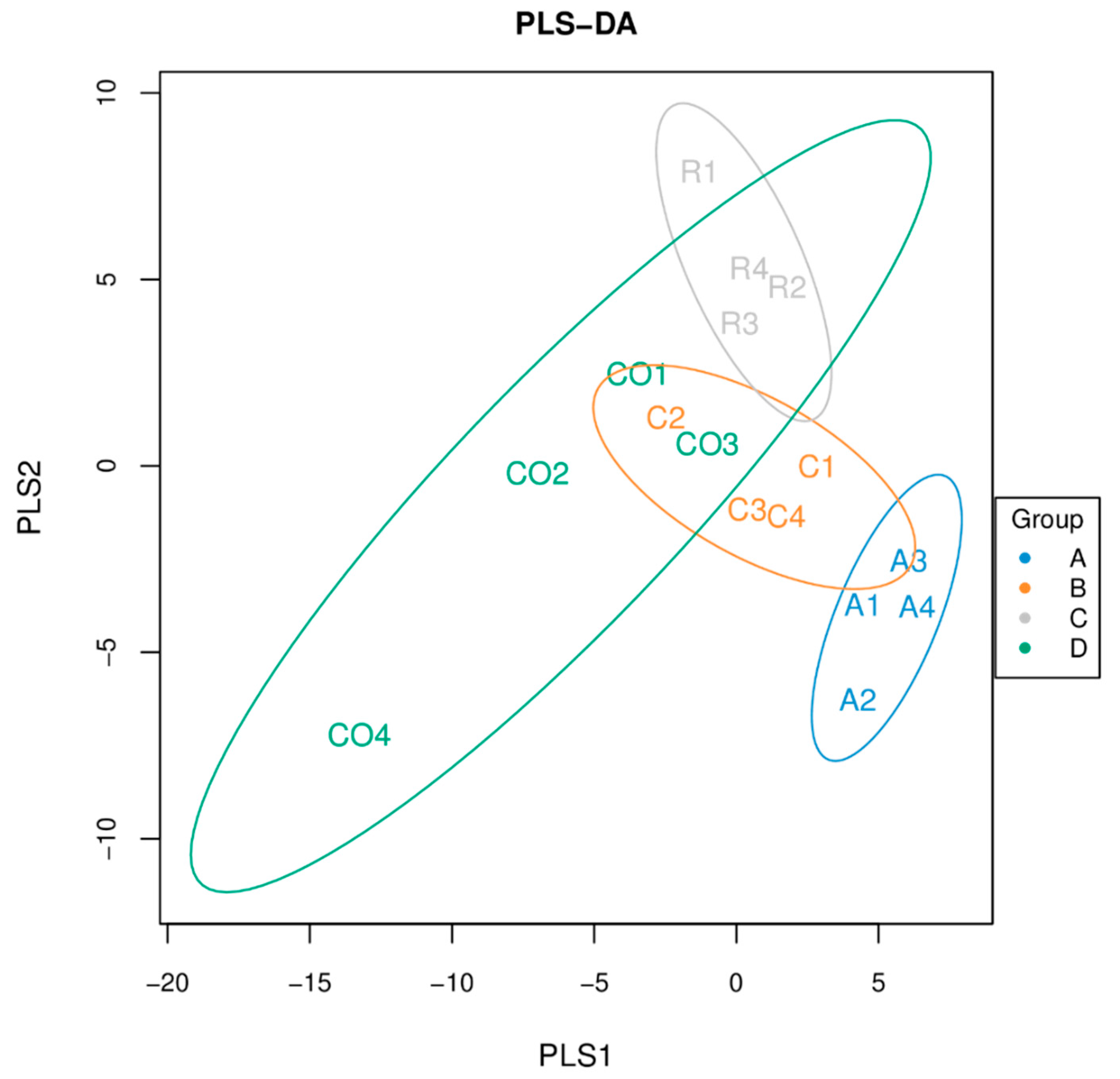
Figure 3 show that the number of microbial species increases as sample size increases, until they reach optimal level. Microbial composition classification on each classification level showed a high number of bacterial species at the genus level and Lactobacilli were the most abundant bacterial genus.
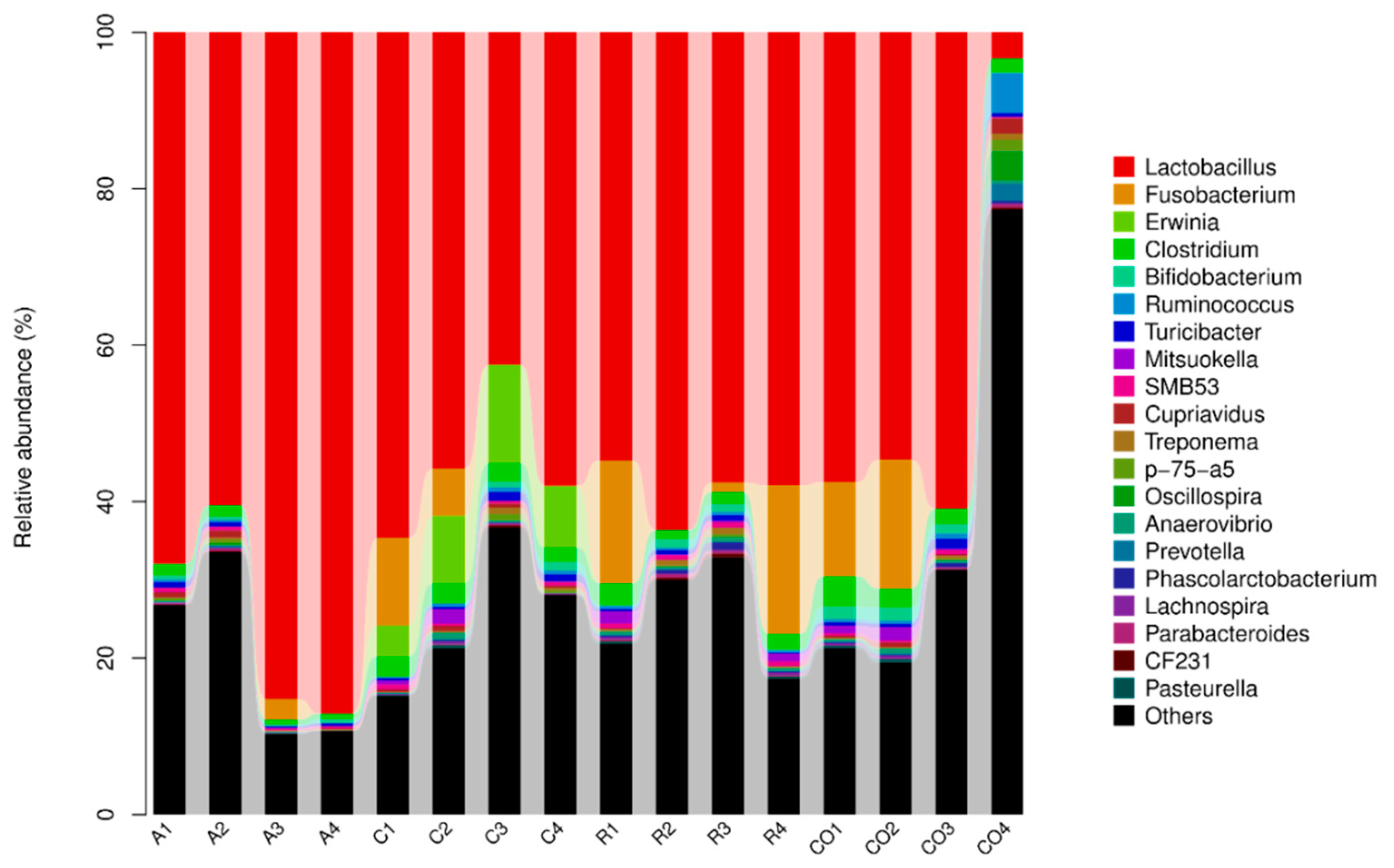
Lactobacillus are considered probiotics and have been associated with the promotion of gastrointestinal tract health [
36]. Lactobacilli prevent infections and colonization of the gut by pathogens and produce antimicrobial factors, including bacteriocins and lactic acids [
37]. Niu et al. [
38] found that Lactobacillus was the abundant genera accounting for more than 84% of total sequences. This study revealed that
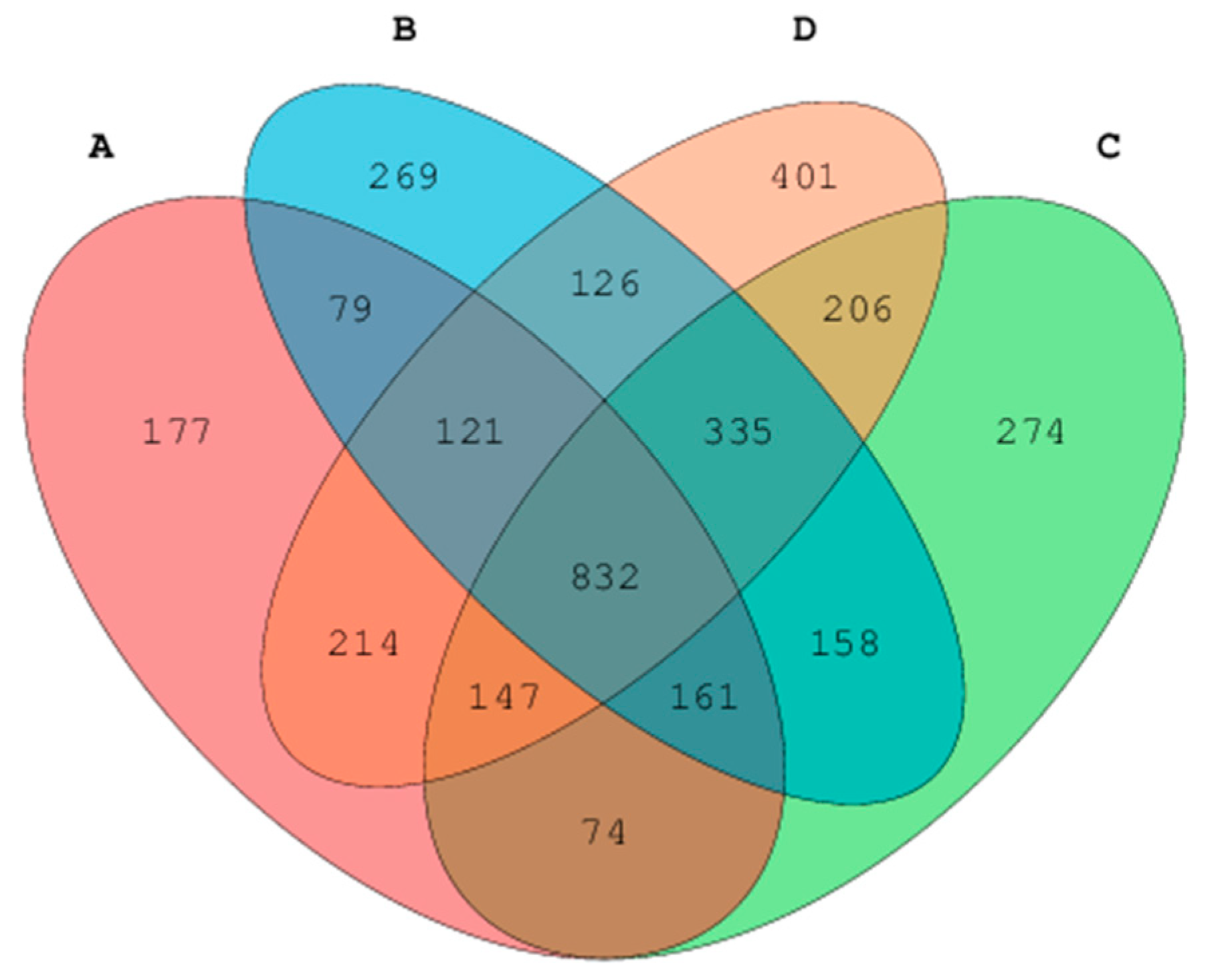
Firmicutes,
Bacteriodetes,
Proteobacteria,
Cynobacteria, and
Actinobacteria were the dorminant bacterial phyla. Furthermore, alfalfa treatment was associated with significant Firmicutes abundance, which accounted for 89.2% sequences. Alfalfa diet has been associated with the ability to decrease the number of pathogenic bacteria and an increase in the number of bacteria that promote alfalfa fiber hydrolysis [
39]. In this study, all treatments and the control showed less effect on the abundance of
Prevotella,
Phascolarctobacteroides,
Parabacteroides,
Ochrobactru, and unclassified
Commenadaceae genera. Previously, other studies revealed that
Prevotella and
Bacteroidetes genera and phylum were abundant in the gut when fibrous diets were used [
40]. In this study, only
Bacteroidetes were abundant, while marginal levels of
Prevotella were observed. However, our results concur with Gorvitovskaia et al. [
41], who showed that there was an inverse correlation between of Bacteriodes and Prevotella: when
Bacteroides numbers were high in a sample,
Prevotella was less abundant and vice versa. Liu et al. [
42] stated that
Prevotella species promote fiber, pectin, protein, and hemicellulose utilization processes and enhance low pH in the ruminal environment caused by sub-acute ruminal acidosis and reduced numbers of
Prevotella. There was a significant difference between all taxonomic units. However,
Gammaproteobacteria,
Enterobacteriales,
Enterobacteriaceae,
Proteobactera, and
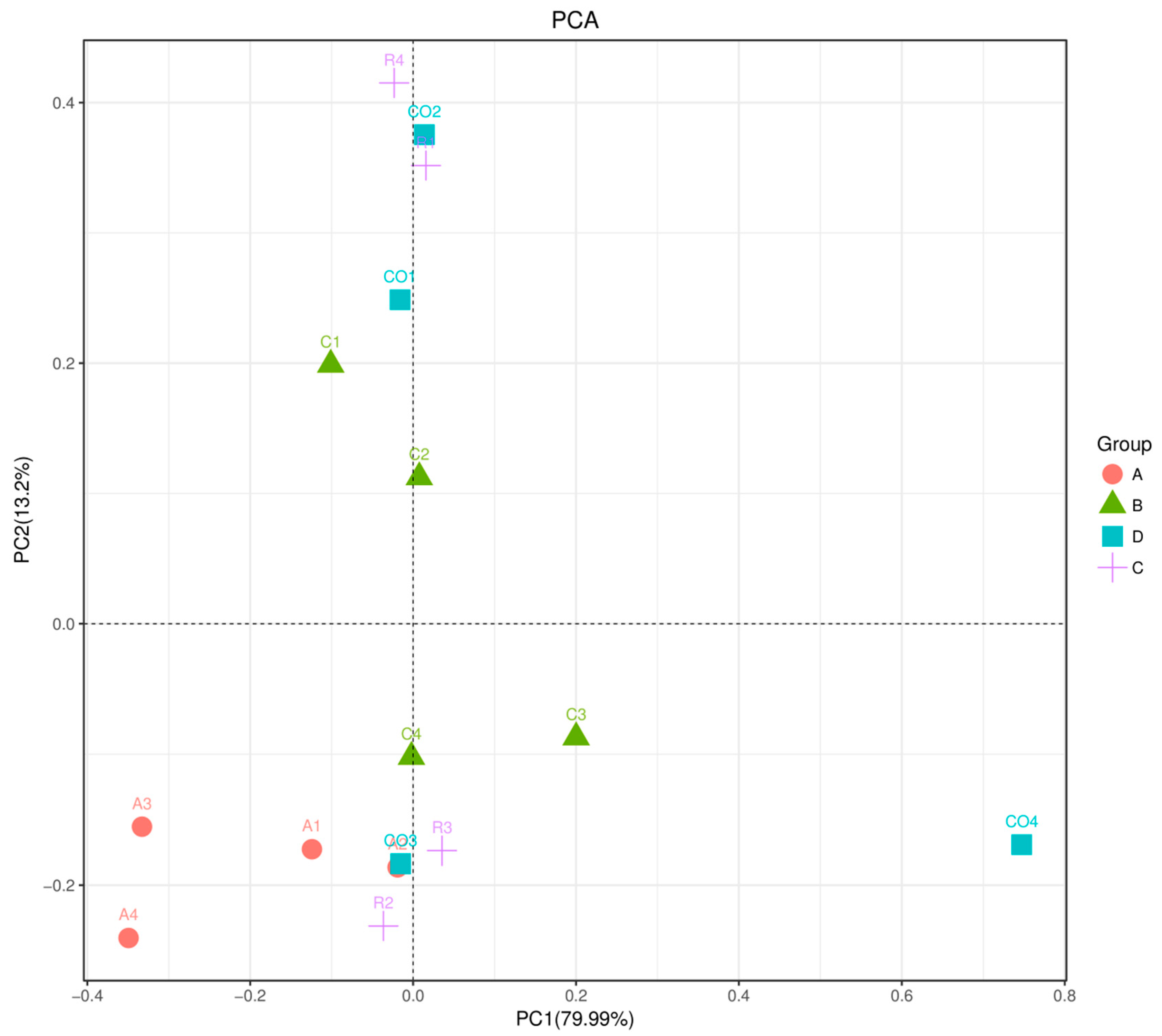
Erwina showed high significance with a 4.7 to 4.8 LDA score (log10) for cornstalk treatment. Principal component analysis (PCA) of microbial evenness and diversity revealed a significant decrease in microbial community diversity among all dietary fiber treatments and marginal diversity in the controls. The three treatments were effective in assessing microbial diversity. Furthermore, rice straw treatment group showed microbial community diversity compared to alfalfa and cornstalk treatment groups, and microbial functions were more available on alfalfa treatment in comparison to other treatments. Nevertheless, the results were based on a limited sample size, and larger well-designed studies are expected to confirm these preliminary findings. Thus, further investigations should be conducted in this line using higher sample sizes. Several factors, such as age, genetic factors, development of diseases, and diet affect the gut microbial community. In this study, the age, breed, previous diet fed, and sex of pigs were not considered.

 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}