Evaluation of Dietary Organic and Inorganic Mercury Threshold Levels on Induced Mercury Toxicity in a Marine Fish Model


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
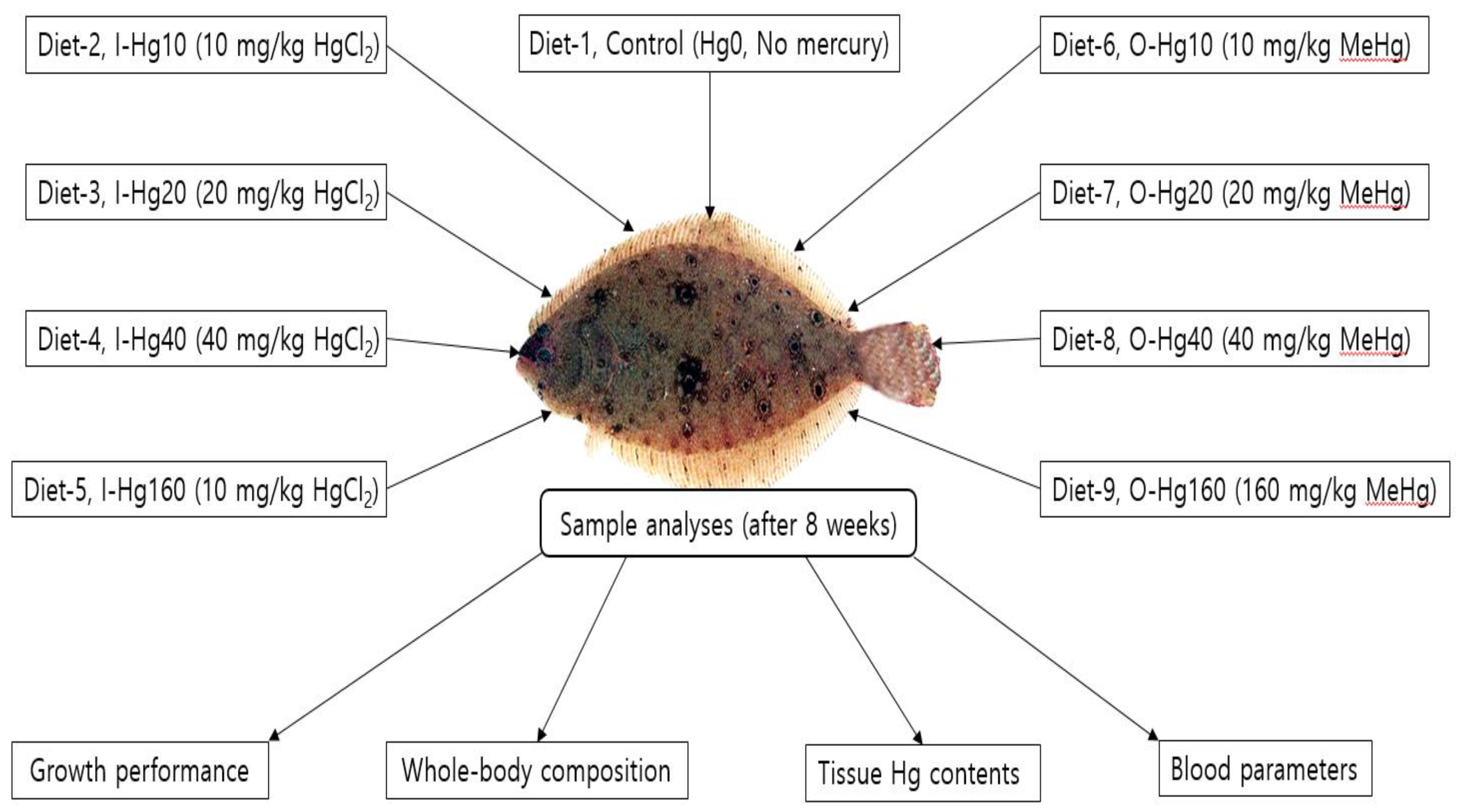
2. Materials and Methods
2.1. Diet Preparation
2.2. Fish and Husbandry Settings
2.3. Experimental Sampling and Analyses
2.4. Statistical Analysis
3. Results
3.1. Growth Performances
3.2. Proximate Composition
3.3. Mercury Concentration
3.4. Blood Biochemical Parameters
3.5. Broken Line Analysis
4. Discussions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gochfeld, M. Cases of mercury exposure, bioavailability, and absorption. Ecotoxicol. Environ. Saf. 2003, 56, 174–179. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Park, G.; Yun, H.; Lee, S.; Park, Y.; Bai, S.C. Synergistic effects of dietary vitamin E and selenomethionine on growth performance and tissue methylmercury accumulation on mercury induced toxicity in juvenile olive flounder, Paralichthys olivaceus (Temminck et Schlegel). Aquac. Res. 2017, 48, 570–580. [Google Scholar] [CrossRef]
- Agency for Toxic Substance and Disease Registry (ATSDR). Toxicological Profile for Mercury; US Department of Health and Humans Services, Public Health Human Services, Centers for Diseases Control: Atlanta, GA, USA, 2003.
- Myers, G.J.; Davidson, P.W.; Strain, J.J. Nutrient and Methyl Mercury Exposure from Consuming Fish. J. Nutri. 2007, 137, 2805–2808. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.; Malgorzata Korbas, M.; Pereiraa, V.; Cappello, T.; Maisano, M.; Canario, J.; Armando Almeida, A.; Pacheco, M. A multidimensional concept for mercury neuronal and sensory toxicity in fish—From toxicokinetics and biochemistry to morphometry and behavior. BBA-Gen. Sub. 2019, 1863, 129298. [Google Scholar] [CrossRef]
- Green, A.J.; Planchart, A. The neurological toxicity of heavy metals: A fish perspective. Comp. Biochem. Physiol. Part C 2018, 208, 12–19. [Google Scholar] [CrossRef]
- Gilmour, C.C.; Henry, E.A. Mercury methylation in aquatic systems affected by acid deposition. Environ. Pollut. 1991, 71, 131–169. [Google Scholar] [CrossRef]
- Bastos, W.R.; Dórea, J.G.; Bernardi, J.V.E.; Lauthartte, L.C.; Mussy, M.H.; Hauser, M.; Dória, C.R.C.; Malm, O. Mercury in muscle and brain of catfish from the Madeira river, Amazon, Brazil. Ecotoxicol. Environ. Saf. 2015, 118, 90–97. [Google Scholar] [CrossRef]
- Adams, D.H.; Sonne, C.; Basu, N.; Dietz, R.; Nam, D.H.; Leifsson, P.S.; Jensen, A.L. Mercury contamination in spotted seatrout, Cynoscion nebulosus: An assessment of liver, kidney, blood, and nervous system health. Sci. Total Environ. 2010, 408, 5808–5816. [Google Scholar] [CrossRef]
- Maulvaulta, A.L.; Custódio, A.; Anacleto, P.; Repolho, T.; Pousãoa, P.; Nunes, M.L.; Diniz, M.; Rosa, R.; Marques, A. Bioaccumulation and elimination of mercury in juvenile seabass (Dicentrarchus labrax) in a warmer environment. Environ. Res. 2016, 149, 77–85. [Google Scholar] [CrossRef]
- International Program on Chemical Safety (IPCS). Elemental Mercury and Inorganic Mercury Compounds; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- Ye, S.; Shin, J.; Lee, J.; Jung, E.M.; Lee, J.; Yun, E.; Kim, Y.; Oh, Y.; Ha, E.H. Systematic Review of Heavy Metal Concentrations in Fish and Shellfish in Korea. Ewha Med. J. 2018, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Niimi, A.J.; Kissoon, G.P. Evaluation of the critical body burden concept based on inorganic and organic mercury toxicity to rainbow trout (Oncorhynchus mykiss). Arch. Environ. Contam. Toxicol. 1994, 26, 169–178. [Google Scholar] [CrossRef]
- Cappello, T.; Brandão, F.; Guilherme, S.; Santos, M.A.; Maisano, M.; Mauceri, A.; João Canário, J.; Pacheco, M.; Pereira, P. Insights into the mechanisms underlying mercury-induced oxidative stress in gills of wild fish (Liza aurata) combining 1H NMR metabolomics and conventional biochemical assays. Sci. Total Environ. 2016, 548, 13–24. [Google Scholar] [CrossRef]
- Cappello, T.; Pereira, P.; Maisano, M.; Mauceri, A.; Pacheco, M.; Fasulo, S. Advances in understanding the mechanisms of mercury toxicity in wild golden grey mullet (Liza aurata) by 1H NMR-based metabolomics. Environ. Pollut. 2016, 219, 139–148. [Google Scholar] [CrossRef]
- Brandao, F.; Cappello, T.; Raimundo, J.; Santos, M.A.; Maisano, M.; Mauceri, A.; Pacheco, M.; Pereira, P. Unravelling the mechanisms of mercury hepatotoxicity in wild fish (Liza aurata) through a triad approach: Bioaccumulation, metabolomics profiles and oxidative stress. Metallomics 2015, 7, 1352–1363. [Google Scholar] [CrossRef]
- Regnell, O.; Tunlid, A. Laboratory study of chemical speciation of mercury in lake sediment and water under aerobic and anaerobic conditions. Appl. Environ. Microbiol. 1991, 57, 789–795. [Google Scholar] [CrossRef] [Green Version]
- Zheng, N.; Wang, S.; Dong, W.; Hua, X.; Li, Y.; Song, X.; Chu, Q.; Hou, S.; Li, Y. The toxicological effects of mercury exposure in marine fish. Bull. Environ. Contam. Toxicol. 2019, 102, 714–720. [Google Scholar] [CrossRef]
- Krabbenhoft, D.P.; Rickert, D.A. Mercury Contamination of Aquatic Ecosystems: U.S. Geological Survey Fact Sheet; 2009, FS-216-95. Available online: https://pubs.usgs.gov/fs/1995/fs216-95/ (accessed on 29 February 2020).
- Berntssen, M.H.G.; Aatland, A.; Handy, R.D. Chronic dietary mercury exposure causes oxidative stress, brain lesions, and altered behaviour in Atlantic salmon (Salmo salar) parr. Aquat. Toxicol. 2003, 65, 55–72. [Google Scholar] [CrossRef]
- Berntssen, M.H.G. Maximum limits of organic and inorganic mercury in fish feed. Aquac. Nutr. 2004, 10, 83–97. [Google Scholar] [CrossRef]
- Pereiraa, P.; Raimundo, J.; Araújo, O.; Canário, J.; Almeida, A.; Pacheco, M. Fish eyes and brain as primary targets for mercury accumulation—A new insight on environmental risk assessment. Sci. Total Environ. 2014, 494, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, O.; Puga, S.; Brandão, F.; Canário, J.; O’Driscoll, N.J.; Santos, M.A.; Pacheco, M.; Pereira, P. Oxidative stress profiles in brain point out a higher susceptibility of fish to waterborne divalent mercury compared to dietary organic mercury. Mar. Pollut. Bull. 2017, 122, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Baatrup, E. Structural & functional effects of heavy metals on the nervous system, including sense organs of fish. Comp. Biochem. Physiol. Part C. 1991, 100, 253–257. [Google Scholar]
- Harris, R.C.; Bodaly, R.A. Temperature, growth & dietary effects on fish mercury dynamics in two Ontario lakes. Biogeochemistry 1998, 40, 175–187. [Google Scholar]
- Rudd, J.W.; Furutani, A.; Turner, M.A. Mercury methylation by fish intestinal contents. Appl. Environ. Microbiol. 1980, 40, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Pedrero, Z.; Gentès, S.; Barre, J.; Renedo, M.; Tessier, E.; Berail, S.; Maury-Brachet, R.; Mesmer-Dudons, N.; Baudrimont, M.; et al. Specific pathways of dietary methylmercury and inorganic mercury determined by mercury speciation and isotopic composition in zebrafish (Danio rerio). Environ. Sci. Technol. 2015, 49, 12984–12993. [Google Scholar] [CrossRef]
- Harayashiki, C.A.Y.; Reichelt-Brushett, A.; Cowden, K.; Benkendorff, K. Effects of oral exposure to inorganic mercury on the feeding behaviour and biochemical markers in yellowfin bream (Acanthopagrus australis). Mar. Environ. Res. 2018, 134, 1–15. [Google Scholar] [CrossRef]
- Gentès, S.; Maury-Brachet, R.; Feng, C.; Pedrero, Z.; Tessier, E.; Legeay, A.; Mesmer-Dudons, N.; Baudrimont, M.; Maurice, L.; Amouroux, D.; et al. Specific Effects of Dietary Methylmercury and Inorganic Mercury in Zebrafish (Danio rerio) Determined by Genetic, Histological, and Metallothionein Responses. Environ. Sci. Technol. 2015, 49, 14560–14569. [Google Scholar] [CrossRef]
- Strungaru, S.A.; Robea, M.A.; Plavan, G.; Todirascu-Ciornea, E.; Ciobica, A.; Nicoara, M. Acute exposure to methylmercury chloride induces fast changes in swimming performance, cognitive processes and oxidative stress of zebrafish (Danio rerio) as reference model for fish community. J. Trace Elem. Med. Biol. 2018, 47, 115–123. [Google Scholar] [CrossRef]
- Mohseni, M.; Park, G.H.; Lee, J.H.; Okorie, O.E.; Browdy, C.; Bharadwaj, A.; Bai, S.C. Evaluation of toxicity of dietary chelated copper in juvenile olive flounder, Paralichthys olivaceus, based on growth and tissue copper concentration. J. World Aquac. Soc. 2012, 43, 548–559. [Google Scholar] [CrossRef]
- Moon, H.B.; Kim, S.J.; Park, H.; Jung, Y.S.; Lee, S.; Kim, Y.H.; Choi, M. Exposure assessment for methyl and total mercury from seafood consumption in Korea, 2005–2008. J. Environ. Monit. 2011, 13, 2400–2405. [Google Scholar] [CrossRef]
- Ikingura, J.R.; Akagi, H. Total mercury and methylmercury levels in fish from hydroelectric reservoirs in Tanzania. Sci. Total Environ. 2003, 304, 355–368. [Google Scholar] [CrossRef]
- Wang, W.X.; Raymond, S.K.; Wong, R.S.K. Bioaccumulation kinetics and exposure pathways of inorganic mercury and methylmercury in a marine fish, the sweetlips Plectorhinchus gibbosus. Mar. Ecol. Prog. Ser. 2003, 261, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Pickhardt, P.C.; Stepanova, M.; Nicholas, S.F. Contrasting Uptake Routes and Tissue Distributions of Inorganic And Methylmercury In Mosquitofish (Gambusia Affinis) and Redear Sunfish (Lepomis Microlophus). Environ. Toxicol. Chem. 2006, 25, 2132–2142. [Google Scholar] [CrossRef]
- Morcillo, P.; Esteban, M.A.; Cuesta, A. Mercury and its toxic effects on fish. AIMS Environ. Sci. 2017, 4, 386–402. [Google Scholar] [CrossRef]
- Poopal, R.K.; Ramesh, M.; Dinesh, B. Short-term mercury exposure on Na+/K+-ATPase activity and ionoregulation in gill and brain of an Indian major carp, Cirrhinus mrigala. J. Trace Elem. Med. Biol. 2013, 27, 70–75. [Google Scholar] [CrossRef]
- Korbas, M.; MacDonald, T.C.; Pickering, I.J.; George, G.N.; Krone, P.H. Chemical form matters: Differential accumulation of mercury following inorganic and organic mercury exposures in Zebrafish Larvae. ACS Chem. Biol. 2012, 7, 411–420. [Google Scholar] [CrossRef]
- Lee, J.W.; Riu, N.D.; Lee, S.; Bai, S.C.; Moniello, G.; Hung, S.S.O. Effects of dietary methylmercury on growth performance and tissue burden in juvenile green (Acipenser medirostris) and white sturgeon (A. transmontanus). Aquat. Toxicol. 2011, 105, 227–234. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis of the Association of Official Analytical Chemists, 17th ed.; Horwitz, W., Ed.; AOAC: Arlington, VA, USA, 2000. [Google Scholar]
- Lee, J.H.; Moniruzzaman, M.; Yun, H.; Lee, S.; Park, Y.; Bai, S.C. Dietary vitamin C reduced mercury contents in the tissues of juvenile olive flounder (Paralichthys olivaceus) exposed with and without mercury. Environ. Toxicol. Pharmacol. 2016, 45, 8–14. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis of the Association of Official Analytical Chemists, 16th ed.; Cunniff, P., Ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1995. [Google Scholar]
- Parrino, V.; Cappello, T.; Costa, G.; Cannavà, C.; Sanfilippo, M.; Fazio, F.; Fasulo, S. Comparative study of haematology of two teleost fish (Mugil cephalus and Carassius auratus) from different environments and feeding habits. Eur. Zool. J. 2018, 85, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Nambi, R.W.; Won, S.H.; Katya, K.; Bai, S.C. Dietary selenium requirement and toxicity levels in juvenile Nile tilapia, Oreochromis niloticus. Aquaculture 2016, 464, 153–158. [Google Scholar] [CrossRef]
- Robbins, K.R.; Norton, H.W.; Baker, D.H. Estimation of nutrient requirements from growth data. J. Nutr. 1979, 109, 1710–1714. [Google Scholar] [CrossRef]
- Rodgers, D.W.; Beamish, F.W.H. Dynamics of dietary methyl mercury in rain-bow trout, Salmo gairdneri. Aquat. Toxicol. 1982, 2, 271–290. [Google Scholar] [CrossRef]
- Houck, A.; Cech, J.J. Effects of dietary methylmercury on juvenile Sacramento blackfish bioenergetics. Aquat. Toxicol. 2004, 69, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Gharaei, A.; Esmaili-Sari, A.; Jafari-shamoshaki, V.; Ghaffari, M. Beluga (Huso huso Brandet 1869) bioenergetics under dietary methylmercury. Fish Physiol. Biochem. 2008, 34, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Wobeser, G. Prolonged oral administration of methyl mercury chloride to rainbow trout (Salmo gairdneri) fingerlings. J. Fish. Res. Board Can. 1975, 32, 2015–2032. [Google Scholar] [CrossRef]
- Friedmann, A.; Watzin, M.; Brinckjohnsen, T.; Leiter, J. Low levels of dietary methylmercury inhibit growth and gonadal development in juvenile walleye (Stizostedion vitreum). Aquat. Toxicol. 1996, 35, 265–728. [Google Scholar] [CrossRef]
- Zalups, R.K.; Lash, L.H. Advances in understanding the renal transport and toxicity of mercury. J. Toxicol. Environ. Health 1994, 42, 1–44. [Google Scholar] [CrossRef]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Rowland, I.R.; Robinson, R.D.; Doherty, R.A. Effects of diet on mercury metabolism and excretion in mice given methylmercury: Role of gut flora. Arch. Environ. Health 1984, 39, 401–408. [Google Scholar] [CrossRef]
- Simon, O.; Boudou, A. Direct and Trophic Contamination of the Herbivorous Carp Ctenopharyngodon idella by Inorganic Mercury and Methylmercury. Ecotoxicol. Environ. Saf. 2001, 50, 48–59. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ingredients | % In Diet |
|---|---|
| Casein 1 | 27.0 |
| Fish meal 2 | 32.0 |
| Wheat flour 3 | 18.0 |
| Dextrin 1 | 4.20 |
| Corn starch 2 | 3.50 |
| Fish oil 4 | 6.10 |
| Vitamin premix 5 | 3.00 |
| Mineral premix 6 | 3.00 |
| Cellulose 1 | 3.20 |
| Hg-premix (5000 ppm) | 0.00 |
| Diets | WG (%) 2 | SGR (%/day) 3 | FE (%) 4 | PER 5 | SR (%) 6 |
|---|---|---|---|---|---|
| Hg0 | 288.4 ± 18.5 a | 2.77 ± 0.10 a | 109.8 ± 10.6 a | 2.33 ± 0.22 a,b | 81.7 ± 7.6 a,b |
| I-Hg10 | 285.3 ± 17.1 a | 2.75 ± 0.09 a | 106.4 ± 6.6 a | 2.29 ± 0.14 a,b | 83.3 ± 5.8 a,b |
| I-Hg20 | 266.9 ± 19.9 b | 2.65 ± 0.11 a,b | 102.1 ± 7.2 a,b | 2.16 ± 0.15 a,b,c | 85.0 ± 5.0 a,b |
| I-Hg40 | 232.1 ± 9.06 c,d | 2.45 ± 0.1 b,c,d | 95.8 ± 11.7 a,b | 2.06 ± 0.25 a,b,c | 78.3 ± 2.8 a,b |
| I-Hg160 | 207.6 ± 6.33 d,e | 2.29 ± 0.1 c,d | 77.4 ± 3.8 c | 1.65 ± 0.08 d | 75.0 ± 5.0 a,b |
| O-Hg10 | 282.9 ± 10.2 a | 2.74 ± 0.05 a | 110.2 ± 7.9 a | 2.34 ± 0.17 a | 81.7 ± 2.8 a,b |
| O-Hg20 | 243.8 ± 12.9 b,c | 2.52 ± 0.08 a,b,c | 94.4 ± 6.2 a,b | 1.99 ± 0.13 b,c | 86.7 ± 5.8 a |
| O-Hg40 | 200.4 ± 3.9 e | 2.24 ± 0.03 d | 87.0 ± 13.9 b,c | 1.87 ± 0.3 c,d | 73.3 ± 14.4 b |
| O-Hg160 | 7.3 ± 20.8 f | 0.12 ± 0.38 e | 4.1 ± 11.1 d | 0.07 ± 0.25 e | 13.3 ± 10.4 c |
| Diets | Moisture (%) | Crude Protein (%) | Crude Lipid (%) | Ash (%) |
|---|---|---|---|---|
| Hg0 | 76.3 ± 0.8 | 70.5 ± 1.1 | 11.7 ± 1.4 a | 12.7 ± 1.2 |
| I-Hg10 | 77.7 ± 0.8 | 70.2 ± 1.6 | 11.1 ± 2.1 a,b | 13.3 ± 0.7 |
| I-Hg20 | 77.5 ± 0.4 | 69.4 ± 2.2 | 11.2 ± 1.2 a,b | 14.1 ± 4.5 |
| I-Hg40 | 78.3 ± 1.0 | 69.1 ± 1.4 | 10.7 ± 0.42 a,b | 14.1 ± 1.5 |
| I-Hg160 | 77.7 ± 0.9 | 69.0 ± 2.2 | 9.9 ± 2.5 a,b | 14.4 ± 0.7 |
| O-Hg10 | 77.8 ± 1.8 | 70.0 ± 1.4 | 11.0 ± 1.4 a,b | 13.4 ± 1.4 |
| O-Hg20 | 77.5 ± 1.0 | 69.7 ± 1.4 | 9.1 ± 0.7 a,b | 14.2 ± 1.6 |
| O-Hg40 | 77.3 ± 3.0 | 69.1 ± 1.6 | 8.8 ± 1.3 b | 15.2 ± 0.7 |
| O-Hg160 | ND | ND | ND | ND |
| Diets | Gill (μg/g) | Kidney (μg/g) | Liver (μg/g) |
|---|---|---|---|
| Hg0 | 1.83 ± 0.51 d | 2.02 ± 0.96 e | 1.38 ± 0.31 e |
| I-Hg10 | 1.63 ± 0.37 d | 2.30 ± 0.31 e | 1.52 ± 0.37 e |
| I-Hg20 | 1.77 ± 0.17 d | 2.88 ± 0.51 e | 3.12 ± 0.64 d,e |
| I-Hg40 | 2.55 ± 0.29 d | 5.09 ± 0.95 d,e | 7.55 ± 5.13 c |
| I-Hg160 | 3.37 ± 0.30 c,d | 13.8 ± 1.79 c | 17.1 ± 2.18 b |
| O-Hg10 | 5.32 ± 0.87 b,c | 8.44 ± 0.56 d | 6.69 ± 1.68 c,d |
| O-Hg20 | 7.58 ± 1.32 b | 17.74 ± 2.18 b | 14.53 ± 2.39 b |
| O-Hg40 | 13.82 ± 3.64 a | 28.29 ± 4.59 a | 24.20 ± 2.53 a |
| O-Hg160 | ND | ND | ND |
| Diets | Total Protein 2 | AST 3 | ALT 4 | Cholesterol 5 |
|---|---|---|---|---|
| Hg0 | 1.8 ± 0.42 | 242 ± 59.6 | 15 ± 8.72 | 141 ± 23.4 |
| I-Hg10 | 2.2 ± 0.36 | 297 ± 52.1 | 11.7 ± 0.58 | 160 ± 13.1 |
| I-Hg20 | 2.2 ± 0.25 | 224 ± 49.24 | 9.0 ± 1.73 | 155 ± 0.0 |
| I-Hg40 | 2.0 ± 0.38 | 268 ± 52.1 | 12.3 ± 1.53 | 150 ± 19.8 |
| I-Hg160 | 1.7 ± 0.40 | 225 ± 38.4 | 10.7 ± 2.52 | 140 ± 13.4 |
| O-Hg10 | 1.9 ± 0.10 | 224 ± 51.9 | 11.0 ± 2.65 | 167 ± 4.51 |
| O-Hg20 | 1.7 ± 0.40 | 254 ± 121.9 | 11.7 ± 6.66 | 142 ± 27.5 |
| O-Hg40 | 1.7 ± 0.35 | 232 ± 48.8 | 11.3 ± 3.21 | 152 ± 14.5 |
| O-Hg160 | ND | ND | ND | ND |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raihan, S.M.; Moniruzzaman, M.; Park, Y.; Lee, S.; Bai, S.C. Evaluation of Dietary Organic and Inorganic Mercury Threshold Levels on Induced Mercury Toxicity in a Marine Fish Model. Animals 2020, 10, 405. https://doi.org/10.3390/ani10030405
Raihan SM, Moniruzzaman M, Park Y, Lee S, Bai SC. Evaluation of Dietary Organic and Inorganic Mercury Threshold Levels on Induced Mercury Toxicity in a Marine Fish Model. Animals. 2020; 10(3):405. https://doi.org/10.3390/ani10030405
Chicago/Turabian StyleRaihan, Said Majdood, Mohammad Moniruzzaman, Youngjin Park, Seunghan Lee, and Sungchul C. Bai. 2020. "Evaluation of Dietary Organic and Inorganic Mercury Threshold Levels on Induced Mercury Toxicity in a Marine Fish Model" Animals 10, no. 3: 405. https://doi.org/10.3390/ani10030405