1. Introduction
Antibiotics such as tetracycline have been used as feed additives to prevent diarrhea, promote animal growth and improve feed utilization in piglets. However, overusing antibiotics in animal feed results in residues in products and causes antibiotic resistance to pathogenic bacteria [
1]. Therefore, it is necessary to find proper alternatives as feed additives to the antibiotics currently used for improving intestinal health and relieving the weaning stress in piglets [
2,
3]. Plant extracts and essential oils might be used as alternatives to antibiotic growth promoters through improving gut health [
4,
5]. Tributyrin (TB) contains three butyric acid moieties esterified to a glycerol backbone [
6]. On a molar basis, when hydrolyzed by cellular lipases or esterase, TB could be3-fold more potent than butyrate, which could result in a higher butyrate level and keep longer in serum as compared with butyrate salts [
7,
8,
9]. TB is well tolerated orally and has been approved as a dietary additive in the United States and many other countries [
10]. Previous studies found that as a short-chain fatty acid (SCFA), butyric acid could be beneficial to intestine health by inhibiting the growth of bacteria, increasing mucosal cell proliferation, and improving intestinal cell development, as well as colonic defense work [
11]. Oregano is a perennial herbaceous plant of the genus
Oryx [
12]. As a plant extract, oregano oil is mainly composed of phenolic compounds including carvacrol, thymol, cymene and terpinene [
13]. Previous studies found that oregano oil could prevent diarrhea in weaned pigs caused by
Escherichia coli and improve overall body weight gain and health [
14]. Methyl salicylate is also an essential oil extracted from the leaves of holly and was found to have bactericidal and bacteriostatic effects in previous studies [
15]. However, little information was available on the effect of methyl salicylate on the intestine health and growth performance of weaning piglets.
Based on our previous in vitro studies on the effect of various combinations of tributyrin and essential oils against
Escherichia coli,
Salmonella and
Staphylococcus aureus [
16], two combinations were selected for the present animal feeding trial. This study was designed to test the hypothesis that dietary supplementation of tributyrin and oregano or tributyrin and methyl salicylate could exert beneficial effects on the intestine health of weaned piglets through both inhibiting
Escherichia coli and promoting mucosal cell proliferation. The major investigated effects included those on the ratio of villus height/crypt depth in the intestine, general health-related blood parameters, and intestinal microbiota and metabolites.
4. Discussion
Supplementation of tributyrinin the diet was able to provide butyric acid for the intestine tissuestodirectly use as nutrients. The fragrances of oregano and methyl salicylate can increase the appetite of the animals, activate their gastrointestinal receptors and stimulate the activity of digestive enzymes, thereby enhance the absorption of nutrients and promote the growth of the body [
29]. A previous study found that supplementation of 0.5% TB in their diet significantly improved daily weight gain and reduced the ratio of feed intake/weight gain in weaned piglets [
30]. However, in a different study it was found that TB at a dosage of 1% in their diet negatively affected the performance of weanling pigs [
31]. Additionally, previous research showed that treatment of oregano oil had no improvement effect on growth performance of nursery pigs [
32]. In this study, the dietary inclusion of OT and MT as well as AB had a numerical improvement on growth performance of the piglets, although the differences between these groups and the Con were not significant. Considering the current study was mainly designed to test the effect on gut health and microbiota and not for growth performance, more trials are needed to test their effect on growth performance in the future.
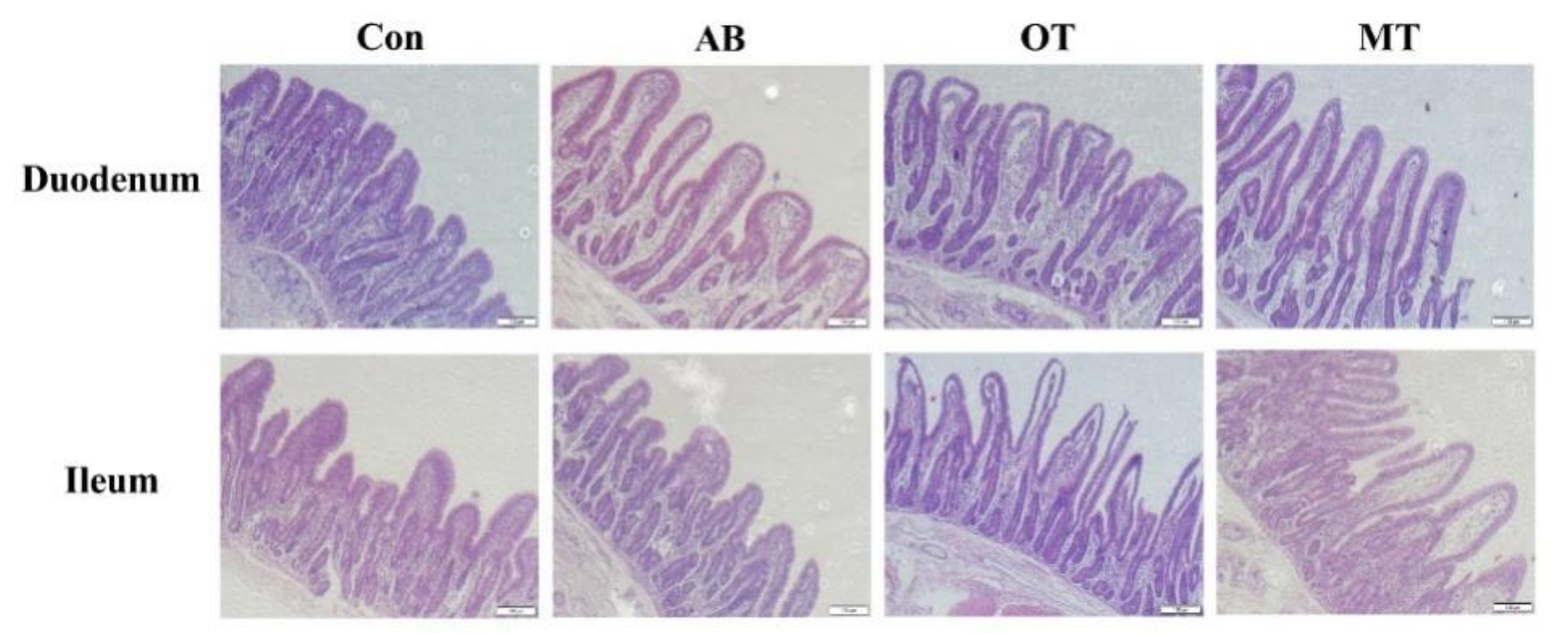
Intestinal morphology is an important factor reflecting intestinal health and function in pigs [
33]. Weaning could cause gastrointestinal tract dysfunction in piglets, which is often due to the contraction of small intestine villi, the increase of crypt depth and the decrease of enzyme activity. These changes in intestinal morphology can cause diarrhea, reduce feed intake and delay the animal’s growth [
34]. In this study, dietary inclusion of OT or MT significantly increased the villus height in the duodenum of piglets. A previous study found that dietary treatment with tributyrin in piglets with intrauterine growth retardation (IUGR) could significantly increase villus length and decrease crypt depth in the duodenum and jejunum [
4]. In different piglet studies it was found that dietary sodium butyrate increased the height of the duodenum villi and the depth of the crypt [
35], whereas inulin-coated sodium butyrate resulted in a 25% larger height of villi of the ileum compared to the control [
36]. Conversely, a study found oral administration of sodium butyrate decreased the jejunum mucosa thickness and villous height [
37]. Also, several studies have found that dietary sodium butyrate treatment has no effect on intestinal mucosal morphology or jejunal villus height in weaned piglets [
38,
39]. In general, our current study suggests that dietary inclusion of the combination of tributyrin and oregano or methyl salicylate effectively promotes the intestinal morphological structure with more healthy characteristics.
The beneficial effect of oregano may be attributed to carvacrol, which contributes to maintaining a good intestinal ecosystem in weaned piglets [
40]. Additionally, its components include carparinol and thyme, which are beneficial to intestinal mucosa and accelerate the regeneration rate of intestinal villi surface epithelial cells [
41]. Methyl salicylate is also an essential oil which has many healthy attributes including antioxidation and anti-inflammation effects. So far, little information is available on the possible effect of methyl salicylate on the intestinal structure of piglets. This study, that showed improving effects on piglet intestine health by dietary inclusion of a combination of tributyrin with oregano or methyl salicylate, confirmed our hypothesis on their beneficial functions.
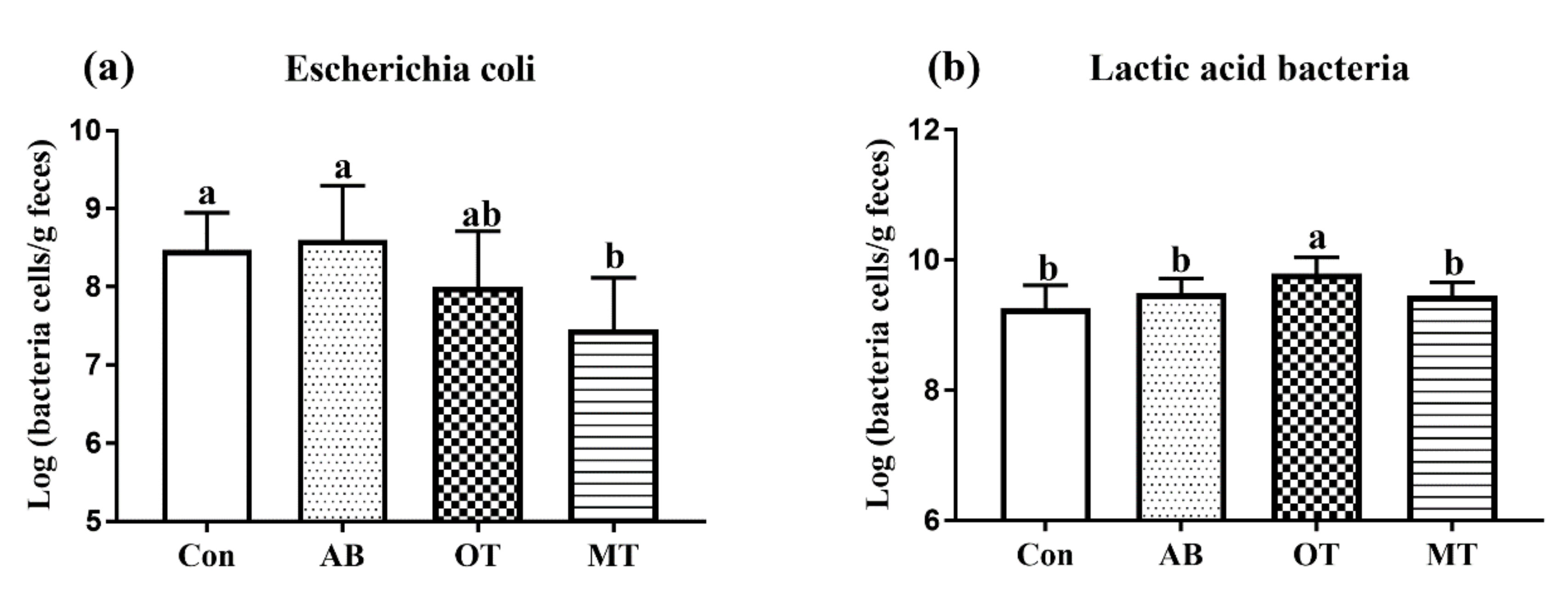
The dynamic balance of intestinal microbes is an important prerequisite for the health of piglets [
42]. When the pH is greater than 4, harmful bacteria in the digestive tract increase while the beneficial bacteria decrease; when the pH value is less than 4, the beneficial bacteria increase while the harmful bacteria decrease [
43]. The addition of tributyrin and essential oils to the feed of piglets reduced the pH of the gut, which might lead to well-balanced intestine microflora. In the bacterial cell, butyric acid is broken down into hydrogen and butyrate ions. The increased concentration of hydrogen ions negatively affects the growth of harmful bacteria such as
Escherichia coli and
Salmonella, but has little effect on beneficial bacteria such as
Lactobacillus, which is acid-resistant [
44]. A previous study found that inclusion of 0.3% butyric acid in the diet of early weaned piglets significantly increased the diversity of intestinal microorganisms [
45]. Another study found that supplementation of 0.4% or 0.6% sodium butyrate also reduced the pH in the stomach and duodenum, and resulted in a reduced number of harmful bacteria in the stomach and cecum [
46].
The essential oils contain bactericidal active components which have strong lipid solubility and surface activity. They can rapidly penetrate the cell membrane of pathogenic bacteria. On the other side, the essential oils might also inhibit the bacteria through a block respiratory oxidation process in mitochondria [
47]. A previous study found that a mixture of thyme and parsley cheese reduced the number of
Escherichia coli but increased that of
Lactobacillus in the feces of weaned piglets [
48], which was similar to the results found in the current study in the OT and MT groups.
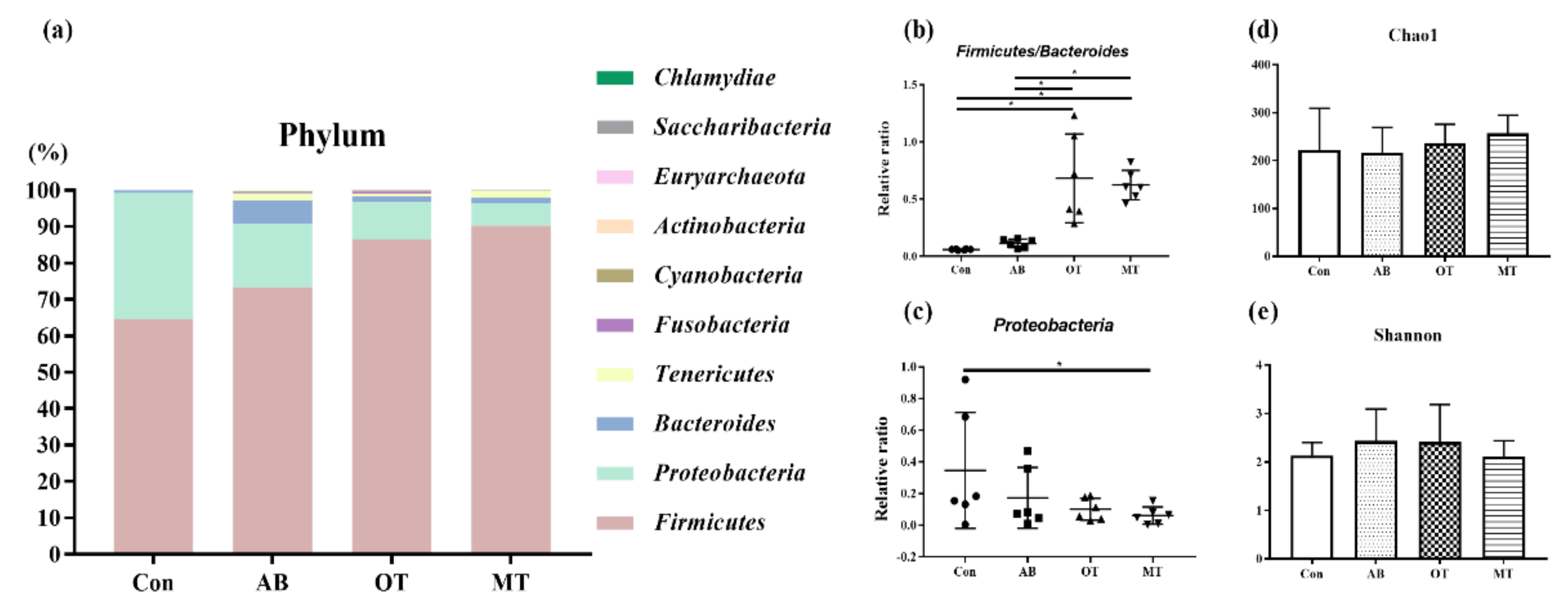
Studies have found that intestinal microbes are mainly anaerobic bacteria [
49]. Through the level analysis on the flora structure in pigs it was found that the dominant bacteria in the intestine were mainly
Bacteroides and
Firmicutes [
50,
51]. Intestinal
Firmicutes and
Bacteroides degrade polysaccharides and promote energy absorption, and therefore the ratio of
Firmicutes to
Bacteroidetes could be of significant relevance to gut microbiota status [
52].
Proteobacteria include many bacteria which might be harmful to intestinal health, such as
Escherichia coli,
Salmonella and
Helicobacter pylori. The abundance of
Proteobacteria is related to the intestinal microbial community and metabolism. Normally, a
Proteobacteria-dominated community is commonly observed in the intestinal tract of organisms with malnutrition and metabolic disorders. Moreover, a
Proteobacteria-dominated community is likely to cause immune disorders in the body which could further lead to intestinal inflammation [
53]. In the current study it was found that dietary tributyrate combined with oregano oil or methyl salicylate significantly increased the relative abundance of
Firmicutes and decreased the relative abundance of
Proteobacteria,
Actinobacillus and
Escherichia, which indicated that both combined treatments exerted a beneficial influence on the development of healthy intestinal flora.
Metabolomic studies on intestinal content may provide more information regarding the effect of dietary treatment on intestinal health and microbiota. The current study found that the dietary treatments resulted in significant changes in some important metabolites i.e., the OT treatment reduced carnitine and the MT treatment increased 3-aminoisobutyric acid. Carnitine is an important metabolite involved in fatty-acids metabolism [
54] whereas 3-aminoisobutyric acid provides energy for cellular metabolism [
55]. Lactic acid could form acetylated acid and enter mitochondria for oxidative phosphorylation. Moreover, it could promote the differentiation and development of bone cells [
56,
57]. The changes in the intestinal metabolites suggest the dietary treatments of OT and MT might affect nutrient digestion and metabolism as well as the profile of intestinal microbiota.
Microbial community diversity and metabolomics analysis may be new indicators of intestine stability and functions which are closely related to animal health; together, the analysis offers a promising approach to evaluate the functional status in the interaction between microbiota and the host intestine [
58]. In the current study, both the microbiota and metabolomics-based analysis were used to evaluate the effect of dietary tributyrin combined with oregano or methyl salicylate on the intestinal microbial society in weaned piglets. Through Pearson analysis on the flora and the major affected metabolites, the study found there were high positive correlations between amounts of
Firmicutes and dihydrocholesterol, as well as between amounts of
Proteobacteria and mannonic acid. Such results on the correlation analysis between intestinal flora and metabolites may provide more evidence that the intestinal flora plays important roles in animal health.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}