Identification of Metabonomics Changes in Longissimus Dorsi Muscle of Finishing Pigs Following Heat Stress through LC-MS/MS-Based Metabonomics Method

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagent
2.2. Animal Treatment and Collection of Samples
2.3. UPLC-ESI-QTOF-MS Analysis
2.4. Data Analysis
2.5. Quantitative Verification of Muscle L-Carnitine Content
2.6. Determination of Intramuscular Fat Content
3. Results
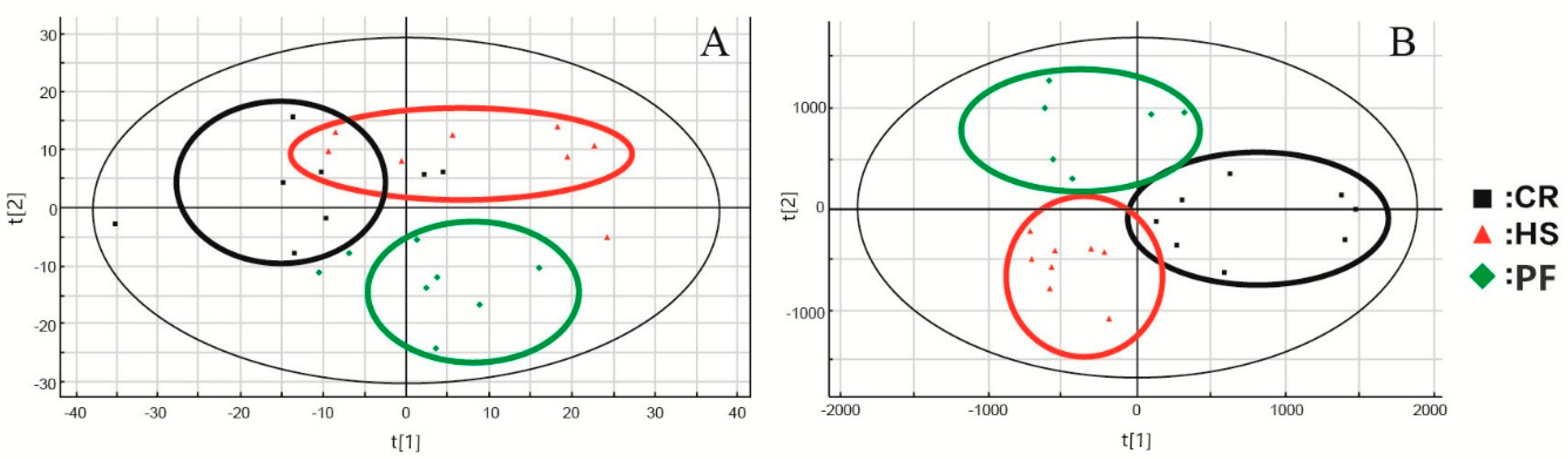
3.1. MDA of Data Matrices
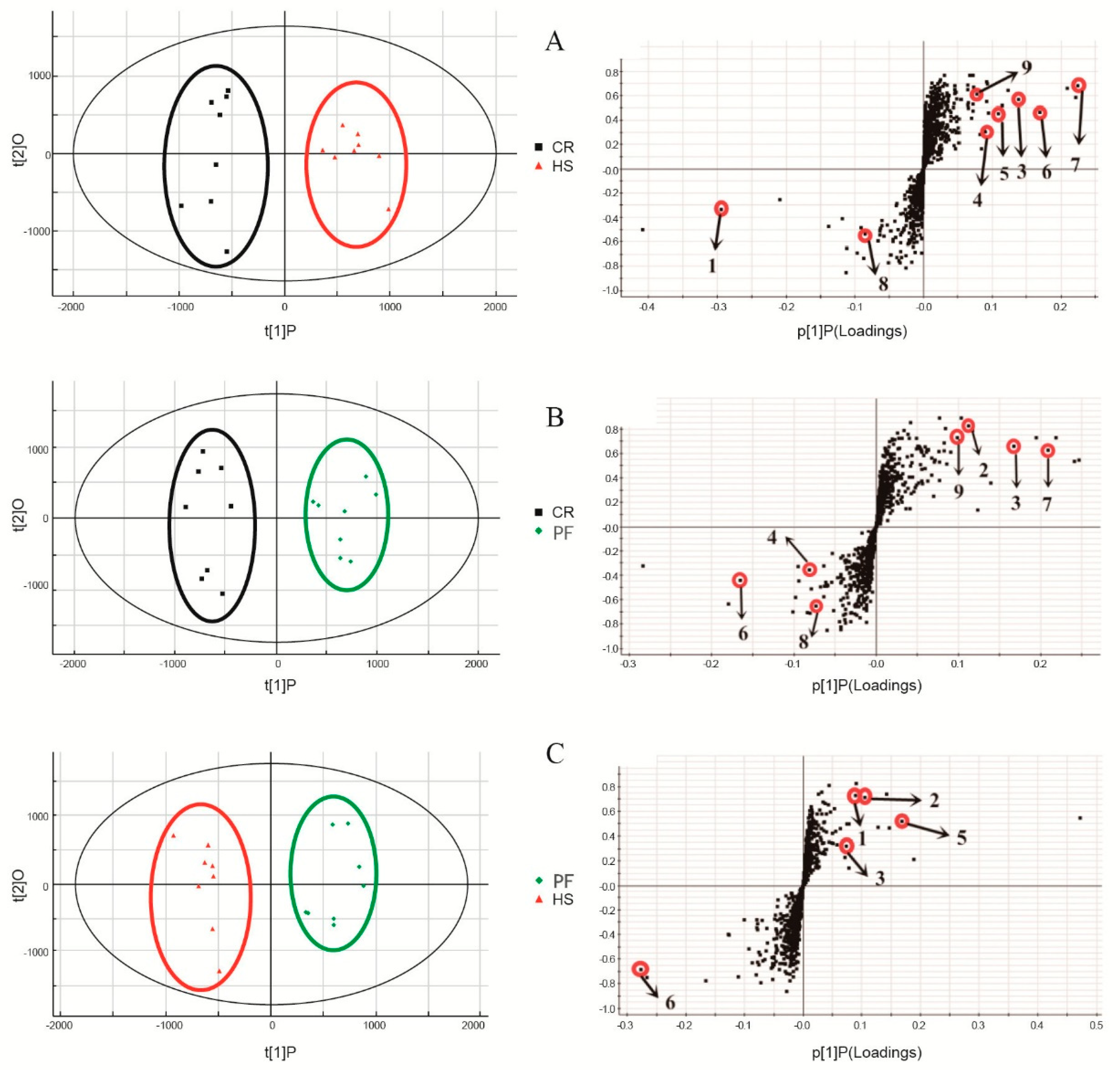
3.2. Differential Metabolites in LD Muscles of Different Treatments
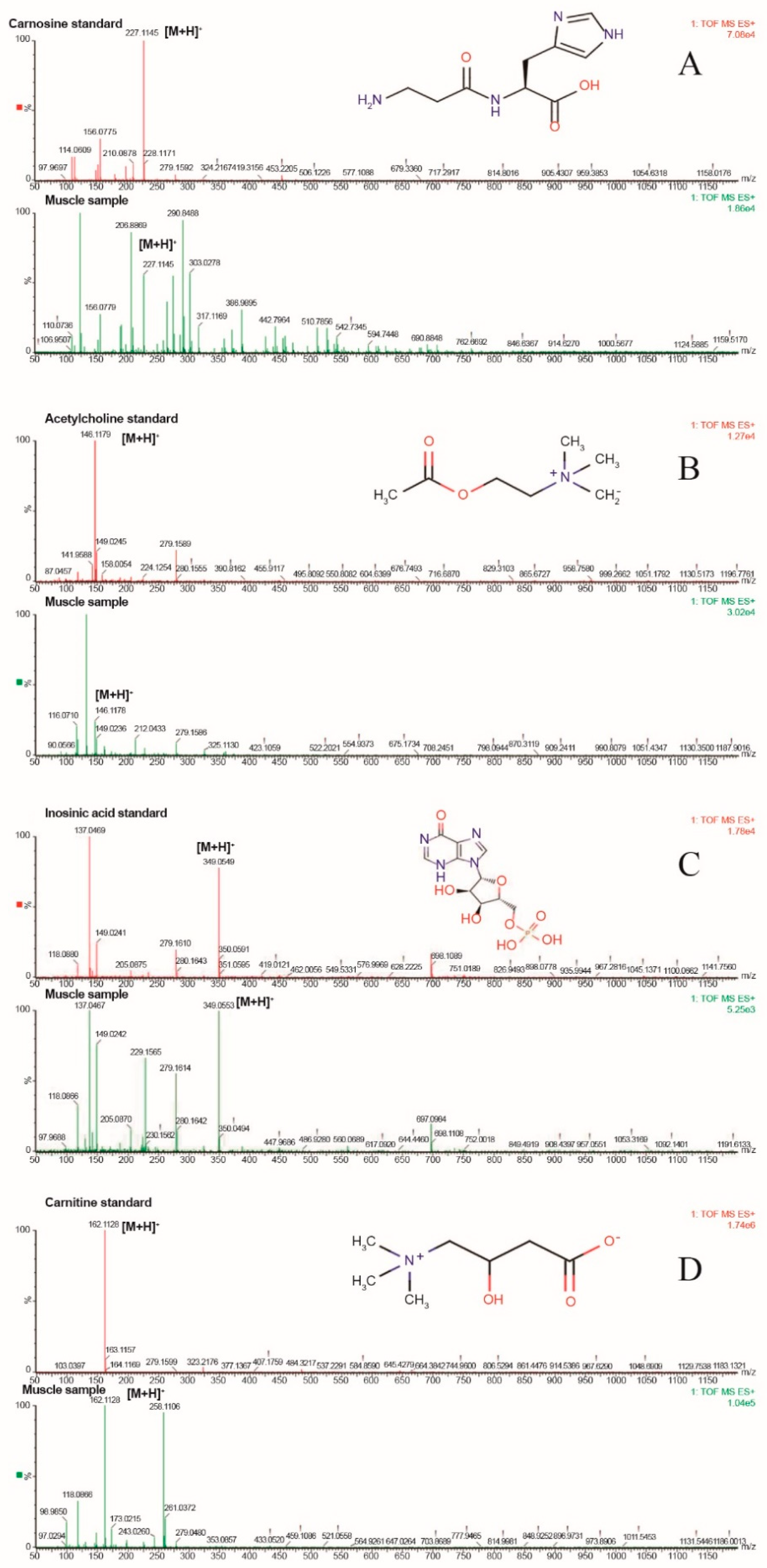
3.3. Qualitative Validation of Differential Metabolites
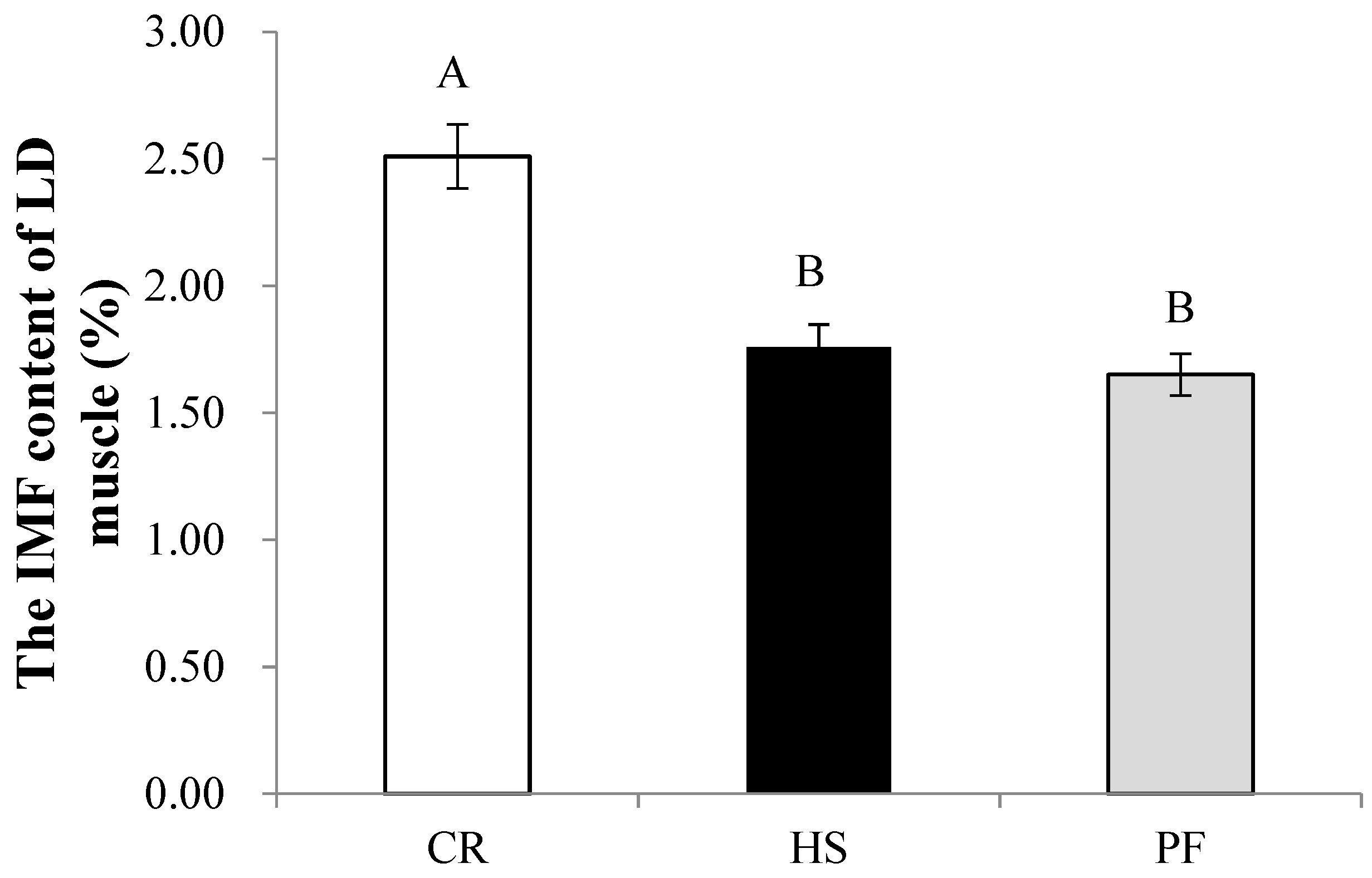
3.4. Quantitative Determination of Muscle Differential Metabolites and IMF
3.5. Correlation Among Metabolites Content and Meat Quality Index
3.6. Correlation Among Metabolites Content and Meat Oxidation Index
4. Discussion
4.1. The Effect of Heat Stress on the Contents of Meat Quality-Related Substances
4.2. The Effects of Heat Stress on Muscle Metabolites Related to Anti-Stress and Anti-Oxidative Damage Ability
4.3. Analysis of the Effect of Heat Stress on Neuromodulatory Function in Muscle Tissue
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Vinoth, A.; Thirunalasundari, T.; Tharian, J.A.; Shanmugam, M.; Rajkumar, U.; Murugesan, S. Effect of thermal manipulation during embryogenesis on liver heat shock protein expression in chronic heat stressed colored broiler chickens. J. Therm. Biol. 2015, 53, 162–171. [Google Scholar] [CrossRef]
- He, X.F.; Lu, Z.; Ma, B.B.; Zhang, L.; Li, J.L.; Jiang, Y.; Zhou, G.H.; Gao, F. Chronic Heat Stress Damages Small Intestinal Epithelium Cells Associated with the Adenosine 5′-Monophosphate-Activated Protein Kinase Pathway in Broilers. J. Agric. Food Chem. 2018, 66, 7301–7309. [Google Scholar] [CrossRef]
- Tang, S.; Yu, J.M.; Zhang, M.; Bao, E.D. Effects of different heat stress periods on various blood and meat quality parameters in young Arbor Acer broiler chickens. Can. J. Anim. Sci. 2013, 93, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Huynh, T.; Aarnink, A.; Gerrits, W.; Heetkamp, M.; Canh, T.; Spoolder, H.; Kemp, B.; Verstegen, M. Thermal behaviour of growing pigs in response to high temperature and humidity. Appl. Anim. Behav. Sci. 2005, 91, 1–16. [Google Scholar] [CrossRef]
- Ganesan, S.; Summers, C.M.; Pearce, S.C.; Gabler, N.K.; Valentine, R.J.; Baumgard, L.H.; Rhoads, R.P.; Selsby, J.T. Short-term heat stress causes altered intracellular signaling in oxidative skeletal muscle. J. Anim. Sci. 2017, 95, 2438–2451. [Google Scholar] [CrossRef] [PubMed]
- Volodina, O.; Ganesan, S.; Pearce, S.C.; Gabler, N.K.; Baumgard, L.H.; Rhoads, R.P.; Selsby, J.T. Short-term heat stress alters redox balance in porcine skeletal muscle. Physiol. Rep. 2017, 5, e13267. [Google Scholar] [CrossRef] [PubMed]
- Charismiadou, M.A.; Goliomytis, M.; Charalambous, A.; Ntetska, I.; Giamouri, E.; Deligeorgis, S.G. Antioxidant status, meat oxidative stability and quality characteristics of lambs fed with hesperidin, naringin or α-tocopheryl acetate supplemented diets: Antioxidant status, meat oxidative stability and quality characteristics of lambs fed with hesperid. J. Sci. Food Agric. 2018, 99, 343–349. [Google Scholar]
- Sandercock, D.A.; Hunter, R.R.; Nute, G.R.; Mitchell, M.A.; Hocking, P.M. Acute Heat Stress-Induced Alterations in Blood Acid-Base Status and Skeletal Muscle Membrane Integrity in Broiler Chickens at Two Ages: Implications for Meat Quality. Poult. Sci. 2001, 80, 418–425. [Google Scholar] [CrossRef]
- Lu, Q.; Wen, J.; Zhang, H. Effect of Chronic Heat Exposure on Fat Deposition and Meat Quality in Two Genetic Types of Chicken. Poult. Sci. 2007, 86, 1059–1064. [Google Scholar] [CrossRef]
- Wang, R.R.; Pan, X.J.; Peng, Z.Q. Effects of heat exposure on muscle oxidation and protein functionalities of pectoralis majors in broilers. Poult. Sci. 2009, 88, 1078–1084. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Jia, G.Q.; Zuo, J.J.; Zhang, Y.; Lei, J.; Ren, L.; Feng, D.Y. Effects of constant and cyclic heat stress on muscle metabolism and meat quality of broiler breast fillet and thigh meat. Poult. Sci. 2012, 91, 2931–2937. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.C.; Mani, V.; Weber, T.E.; Rhoads, R.P.; Patience, J.F.; Baumgard, L.H.; Gabler, N.K. Heat stress and reduced plane of nutrition decreases intestinal integrity and function in pigs. J. Anim. Sci. 2013, 91, 5183–5193. [Google Scholar] [CrossRef]
- Huynh, T.T.T.; Aarnink, A.J.A.; Verstegen, M.W.A.; Gerrits, W.J.J.; Heetkamp, M.J.W.; Kemp, B.; Canh, T.T. Effects of increasing temperatures on physiological changes in pigs at different relative humidities. J. Anim. Sci. 2005, 83, 1385–1396. [Google Scholar] [CrossRef]
- Pearce, S.C.; Gabler, N.K.; Ross, J.W.; Escobar, J.; Patience, J.F.; Rhoads, R.P.; Baumgard, L.H. The effects of heat stress and plane of nutrition on metabolism in growing pigs. J. Anim. Sci. 2013, 91, 2108–2118. [Google Scholar] [CrossRef] [Green Version]
- Seibert, J.T.; Abuajamieh, M.; Fernandez, M.V.S.; Johnson, J.S.; Kvidera, S.K.; Horst, E.A.; Mayorga, E.J.; Lei, S.; Patience, J.F.; Ross, J.W.; et al. Effects of heat stress and insulin sensitizers on pig adipose tissue. J. Anim. Sci. 2018, 96, 510–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahjoubi, E.; Amanlou, H.; Mirzaei-Alamouti, H.R.; Aghaziarati, N.; Yazdi, M.H.; Noori, G.R.; Yuan, K.; Baumgard, L.H. The effect of cyclical and mild heat stress on productivity and metabolism in Afshari lambs. J. Anim. Sci. 2014, 92, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Bujak, R.; Struck-Lewicka, W.; Markuszewski, M.J.; Kaliszan, R. Metabolomics for laboratory diagnostics. J. Pharm. Biomed. Anal. 2015, 113, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Innovation: Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Serum metabolomics study of nutrient metabolic variations in chronic heat-stressed broilers. Br. J. Nutr. 2018, 119, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Zhao, W.; Wang, L.; Guo, F.; Song, D.; Zhang, Q.; Zhang, D.; Fan, Y.; Wang, J. Integration of metabolomic and transcriptomic profiles to identify biomarkers in serum of lung cancer. J. Cell. Biochem. 2019, 120, 11981–11989. [Google Scholar] [CrossRef]
- Yang, P.; Hao, Y.; Feng, J.; Lin, H.; Feng, Y.; Wu, X.; Yang, X.; Gu, X. The Expression of Carnosine and Its Effect on the Antioxidant Capacity of Longissimus dorsi Muscle in Finishing Pigs Exposed to Constant Heat Stress. Asian Australas. J. Anim. Sci. 2014, 27, 1763–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milfort, M.C.; Rekaya, R.; Aggrey, S.E. Cellular antioxidant enzyme activity and biomarkers for oxidative stress are affected by heat stress. Int. J. Biometeorol. 2019, 63, 1569–1584. [Google Scholar]
- Hoenicke, K.; Borchert, O.; Grüning, K.; Simat, T.J. “Untypical aging off-flavor” in wine: Synthesis of potential degradation compounds of indole-3-acetic acid and kynurenine and their evaluation as precursors of 2-aminoacetophenone. J. Agric. Food Chem. 2002, 50, 4303–4309. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Wang, S.; Xu, Z.; Wang, X.; Feng, J.; Shan, T.; Wang, Y. Betaine promotes lipid accumulation in adipogenic-differentiated skeletal muscle cells through ERK/PPARγ signalling pathway. Mol. Cell. Biochem. 2018, 447, 137–149. [Google Scholar] [CrossRef]
- Yong, Z.; Ma, Q.G.; Bai, X.M.; Zhao, L.H.; Wang, Q.; Ji, C.; Liu, L.T.; Yin, H.C. Effects of Dietary Acetyl-L-Carnitine on Meat Quality and Lipid Metabolism in Arbor Acres Broilers. Asian Australas. J. Anim. Sci. 2010, 23, 1639–1644. [Google Scholar]
- Di Cesare Mannelli, L.; Ghelardini, C.; Toscano, A.; Pacini, A.; Bartolini, A. The neuropathy-protective agent acetyl-l-carnitine activates protein kinase C-γ and MAPKs in a rat model of neuropathic pain. Neuroscience 2010, 165, 1345–1352. [Google Scholar] [CrossRef]
- Cui, Y.; Hao, Y.; Li, J.; Gao, Y.; Gu, X. Proteomic changes of the porcine skeletal muscle in response to chronic heat stress. J. Sci. Food Agric. 2018, 98, 3315–3323. [Google Scholar] [CrossRef]
- Kurihara, K. Umami the Fifth Basic Taste: History of Studies on Receptor Mechanisms and Role as a Food Flavor. BioMed Res. Int. 2015, 2015, 189402. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.H.; Chen, J.L.; Zhao, G.P.; Zheng, M.Q.; Wen, J. Correlation between polymorphisms in ADSL and GARS-AIRS-GART genes with inosine 5′-monophosphate (IMP) contents in Beijing-you chickens. Br. Poult. Sci. 2010, 51, 609–613. [Google Scholar] [CrossRef]
- Ye, J.; Li, J.; Yu, Y.; Wei, Q.; Deng, W.; Yu, L. l-carnitine attenuates oxidant injury in HK-2 cells via ROS-mitochondria pathway. Regul. Pept. 2010, 161, 58–66. [Google Scholar] [CrossRef]
- Ramayo-Caldas, Y.; Mach, N.; Esteve-Codina, A.; Corominas, J.; Castelló, A.; Ballester, M.; Estellé, J.; Ibáñez-Escriche, N.; Fernández, A.I.; Pérez-Enciso, M.; et al. Liver transcriptome profile in pigs with extreme phenotypes of intramuscular fatty acid composition. BMC Genom. 2012, 13, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannier, L.; Mullen, A.; Hamill, R.; Stapleton, P.; Sweeney, T. Association analysis of single nucleotide polymorphisms in DGAT1, TG and FABP4 genes and intramuscular fat in crossbred Bos taurus cattle. Meat Sci. 2010, 85, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Dashdorj, D.; Amna, T.; Hwang, I. Influence of specific taste-active components on meat flavor as affected by intrinsic and extrinsic factors: An overview. Eur. Food Res. Technol. 2015, 241, 157–171. [Google Scholar] [CrossRef]
- Katakura, Y.; Totsuka, M.; Imabayashi, E.; Matsuda, H.; Hisatsune, T. Anserine/Carnosine Supplementation Suppresses the Expression of the Inflammatory Chemokine CCL24 in Peripheral Blood Mononuclear Cells from Elderly People. Nutrients 2017, 9, 1199. [Google Scholar] [CrossRef] [Green Version]
- Stvolinsky, S.; Oropova, K.; Gordeeva, M.; Kazey, V.; Sato, T.; Meguro, K.; Boldyrev, A. Carnosine and its (S)-Trolox™ derivative protect animals against oxidative stress. Amino Acids 2012, 43, 165–170. [Google Scholar] [CrossRef]
- Kulebyakin, K.; Karpova, L.; Lakonsteva, E.; Krasavin, M.; Boldyrev, A. Carnosine protects neurons against oxidative stress and modulates the time profile of MAPK cascade signaling. Amino Acids 2012, 43, 91–96. [Google Scholar] [CrossRef]
- Rocha, C.C.; Kawai, G.K.V.; de Agostini Losano, J.D.; de Souza Ramos Angrimani, D.; Rui, B.R.; de Cássia Bicudo, L.; da Silva, B.D.C.S.; Alonso, M.A.; Mendes, C.M.; Assumpção, M.E.O.D.; et al. Carnosine as malondialdehyde scavenger in stallion seminal plasma and its role in sperm function and oxidative status. Theriogenology 2018, 119, 10–17. [Google Scholar] [CrossRef]
- Lee, J.; Park, J.R.; Lee, H.; Jang, S.; Ryu, S.M.; Kim, H.; Kim, D.; Jang, A.; Yang, S.R. l-carnosine induces apoptosis/cell cycle arrest via suppression of NF-kappa B/STAT1 pathway in HCT116 colorectal cancer cells. In Vitro Cell. Dev. Biol. Anim. 2018, 54, 505–512. [Google Scholar] [CrossRef]
- Manhiani, P.S.; Northcutt, J.K.; Han, I.; Bridges, W.C.; Scott, T.R.; Dawson, P.L. Effect of stress on carnosine levels in brain, breast, and thigh of broilers. Poult. Sci. 2011, 90, 2348–2354. [Google Scholar] [CrossRef]
- Annadurai, T.; Vigneshwari, S.; Thirukumaran, R.; Thomas, P.A.; Geraldine, P. Acetyl-l-carnitine prevents carbon tetrachloride-induced oxidative stress in various tissues of Wistar rats. J. Physiol. Biochem. 2011, 67, 519–530. [Google Scholar] [CrossRef]
- Assaf, N.; Shalby, A.B.; Khalil, W.K.; Ahmed, H.H. Biochemical and genetic alterations of oxidant/antioxidant status of the brain in rats treated with dexamethasone: Protective roles of melatonin and acetyl-l-carnitine. J. Physiol. Biochem. 2012, 68, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Zidan, A.; Hedya, S.E.; Elfeky, D.M.; Abdin, A.A. The possible anti-apoptotic and antioxidant effects of acetyl l-carnitine as an add-on therapy on a relapsing-remitting model of experimental autoimmune encephalomyelitis in rats. Biomed. Pharmacother. 2018, 103, 1302–1311. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Xie, S.J.; Zhao, Y.P.; Dong, Z.J.; Zou, Z.M. Effects of heat stress with organophosphorus pesticide intoxication on blood AChE activity and tissue anti-oxidation ability in mice. Mil. Med. Sci. 2012, 36, 430–433. [Google Scholar]
- Kang, S.Z.; Zhang, H.C.; Wu, Y.H.; Gu, K.R. Advance in preparation, purification and detection of l-alpha-glycerylphosphorylcholine. China Oils Fats 2017, 42, 103–107. [Google Scholar]
- Zhang, J.X.Z. The Effect of the Exercise Performance with the Lack of Vitamin B1. J. Ankang Teach. Coll. 2003, 15, 76–78. [Google Scholar]
- Collie, J.T.; Greaves, R.F.; Jones, O.A.; Lam, Q.; Eastwood, G.M.; Bellomo, R. Vitamin B1 in critically ill patients: Needs and challenges. Clin. Chem. Lab. Med. 2017, 55, 1652–1668. [Google Scholar] [CrossRef]
- Hao, Y.; Feng, Y.; Yang, P.; Feng, J.; Lin, H.; Gu, X. Nutritional and physiological responses of finishing pigs exposed to a permanent heat exposure during three weeks. Arch. Anim. Nutr. 2014, 68, 296–308. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human Metabolome Database ID | Symbol ID | Identity | Formula | tR (min) | HS vs CR | PF vs CR | HS vs PF | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| VIP Values | p-Values | Fold Change | VIP Values | p-Values | Fold Change | VIP Values | p-Values | Fold Change | |||||
| HMDB00033 | 1 | L-carnosine | C9H14N4O3 | 0.94 | 11.69 | 0.002 | 14.3↓ | - | - | - | 3.02 | 0.008 | 1.45↓ |
| HMDB00895 | 2 | acetylcholine | C7H16NO2 | 0.89 | - | - | - | 3.66 | 0.026 | 1.47↑ | 3.25 | 0.020 | 1.32↓ |
| HMDB00175 | 3 | inosinic acid | C10H13N4O8P | 1.52 | 4.39 | 0.006 | 1.58↑ | 5.70 | 0.002 | 1.80↑ | 2.04 | 6.94E-5 | 1.14↓ |
| HMDB00062 | 4 | L-carnitine | C7H15NO3 | 0.83 | 2.78 | 0.031 | 1.13↓ | 2.93 | 0.080 | 1.13↓ | - | - | - |
| HMDB00194 | 5 | L-anserine | C10H16N4O3 | 0.87 | 3.10 | 0.121 | 1.34↑ | - | - | - | 5.68 | 0.073 | 1.55↓ |
| HMDB00086 | 6 | L-α-glycerylphosphorylcholine | C8H20NO6P | 0.79 | 4.46 | 0.054 | 1.20↑ | 5.05 | 0.163 | 1.23↓ | 9.96 | 0.014 | 1.43↑ |
| HMDB00201 | 7 | acetylcarnitine | C9H17NO4 | 1.32 | 7.66 | 0.050 | 1.20↑ | 6.89 | 0.023 | 1.22↑ | - | - | - |
| HMDB01512 | 8 | thiamin triphosphate | C12H20N4O10P3S | 0.86 | 2.40 | 0.010 | 4.34↓ | 2.80 | 0.010 | 13.4↓ | - | - | - |
| HMDB13647 | 9 | adenosine thiamine diphosphate | C22H30N9O10P2S | 1.51 | 2.62 | 0.004 | 2.12↑ | 3.44 | 0.001 | 2.61↑ | - | - | - |
| Metabolites | pH 45 min | pH 24 h | pH 48 h | Drip Loss 24 h | Drip Loss 48 h | L* 45 min | a* 45 min | b* 45 min | L* 24 h | a* 24 h | b* 24 h | Ec 45 min | Ec 48 h |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L-anserine | −0.20 | −0.26 | 0.12 | 0.20 | 0.03 | −0.18 | −0.07 | −0.26 | −0.26 | 0.00 | −0.12 | −0.22 | 0.35 |
| thiamin triphosphate | 0.22 | 0.27 | 0.11 | −0.30 | −0.50 ** | −0.24 | 0.41 * | 0.53 ** | −0.03 | 0.08 | 0.50 ** | 0.07 | −0.23 |
| inosinic acid | −0.21 | −0.13 | 0.22 | 0.16 | 0.24 | 0.04 | −0.12 | −0.12 | −0.09 | 0.18 | −0.00 | −0.33 | 0.17 |
| L-α-glycerylphosphorylcholine | −0.11 | 0.11 | 0.10 | 0.28 | 0.28 | 0.40 | 0.19 | 0.29 | 0.59 ** | −0.10 | 0.27 | −0.26 | 0.43 * |
| L-carnitine | −0.01 | 0.02 | 0.22 | 0.36 | −0.07 | −0.11 | 0.22 | 0.05 | 0.29 | −0.22 | 0.16 | −0.24 | 0.32 |
| Acetylcarnitine | 0.08 | −0.15 | −0.12 | 0.06 | −0.01 | 0.08 | −0.26 | −0.15 | 0.10 | −0.05 | 0.06 | −0.18 | −0.24 |
| L-carnosine | 0.10 | 0.35 | 0.13 | −0.10 | −0.11 | −0.07 | 0.05 | 0.34 | −0.01 | 0.20 | 0.08 | −0.16 | −0.04 |
| Acetylcholine | 0.04 | −0.18 | −0.06 | −0.17 | −0.13 | −0.39 * | −0.33 | −0.35 | −0.56 ** | 0.12 | −0.20 | −0.09 | −0.25 |
| adenosine thiamine diphosphatedenosine | −0.15 | −0.13 | 0.25 | 0.15 | 0.23 | −0.02 | −0.15 | −0.19 | −0.11 | 0.15 | −0.05 | −0.34 | 0.16 |
| Items | L-anserine | Thiamin Triphosphate | Inosinic Acid | L-α-glycerylphosphorylcholine | L-carnitine | Acetylcarnitine | L-carnosine | Acetylcholine | Adenosine Thiamine Diphosphatedenosine |
|---|---|---|---|---|---|---|---|---|---|
| MDA | 0.00 | −0.33 | 0.27 | 0.31 | 0.23 | 0.48 ** | −0.35 | −0.02 | 0.28 |
| SOD | −0.39 | 0.67 ** | −0.51 ** | 0.03 | −0.06 | −0.48 ** | 0.32 | −0.39 | −0.56 ** |
| LDH | 0.22 | −0.32 | 0.14 | 0.01 | 0.20 | 0.21 | −0.15 | 0.21 | 0.20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Yang, P.; Cui, Y.; Meng, Q.; Feng, Y.; Hao, Y.; Liu, J.; Piao, X.; Gu, X. Identification of Metabonomics Changes in Longissimus Dorsi Muscle of Finishing Pigs Following Heat Stress through LC-MS/MS-Based Metabonomics Method. Animals 2020, 10, 129. https://doi.org/10.3390/ani10010129
Gao J, Yang P, Cui Y, Meng Q, Feng Y, Hao Y, Liu J, Piao X, Gu X. Identification of Metabonomics Changes in Longissimus Dorsi Muscle of Finishing Pigs Following Heat Stress through LC-MS/MS-Based Metabonomics Method. Animals. 2020; 10(1):129. https://doi.org/10.3390/ani10010129
Chicago/Turabian StyleGao, Jie, Peige Yang, Yanjun Cui, Qingshi Meng, Yuejin Feng, Yue Hao, Jiru Liu, Xiangshu Piao, and Xianhong Gu. 2020. "Identification of Metabonomics Changes in Longissimus Dorsi Muscle of Finishing Pigs Following Heat Stress through LC-MS/MS-Based Metabonomics Method" Animals 10, no. 1: 129. https://doi.org/10.3390/ani10010129