Application of Selenium Can Alleviate the Stress of Cadmium on Rapeseed at Different Growth Stages in Soil

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimentation
2.3. Plant Sampling
2.4. Soil Sampling
2.5. DNA Extract and PCR Products Mixing and Purification
2.6. High-Throughput Sequencing
2.7. Statistical Analysis
3. Results
3.1. Effects of Different Amounts of Selenium Application on Agronomic Characteristics and Yield of Rapeseed during Maturity
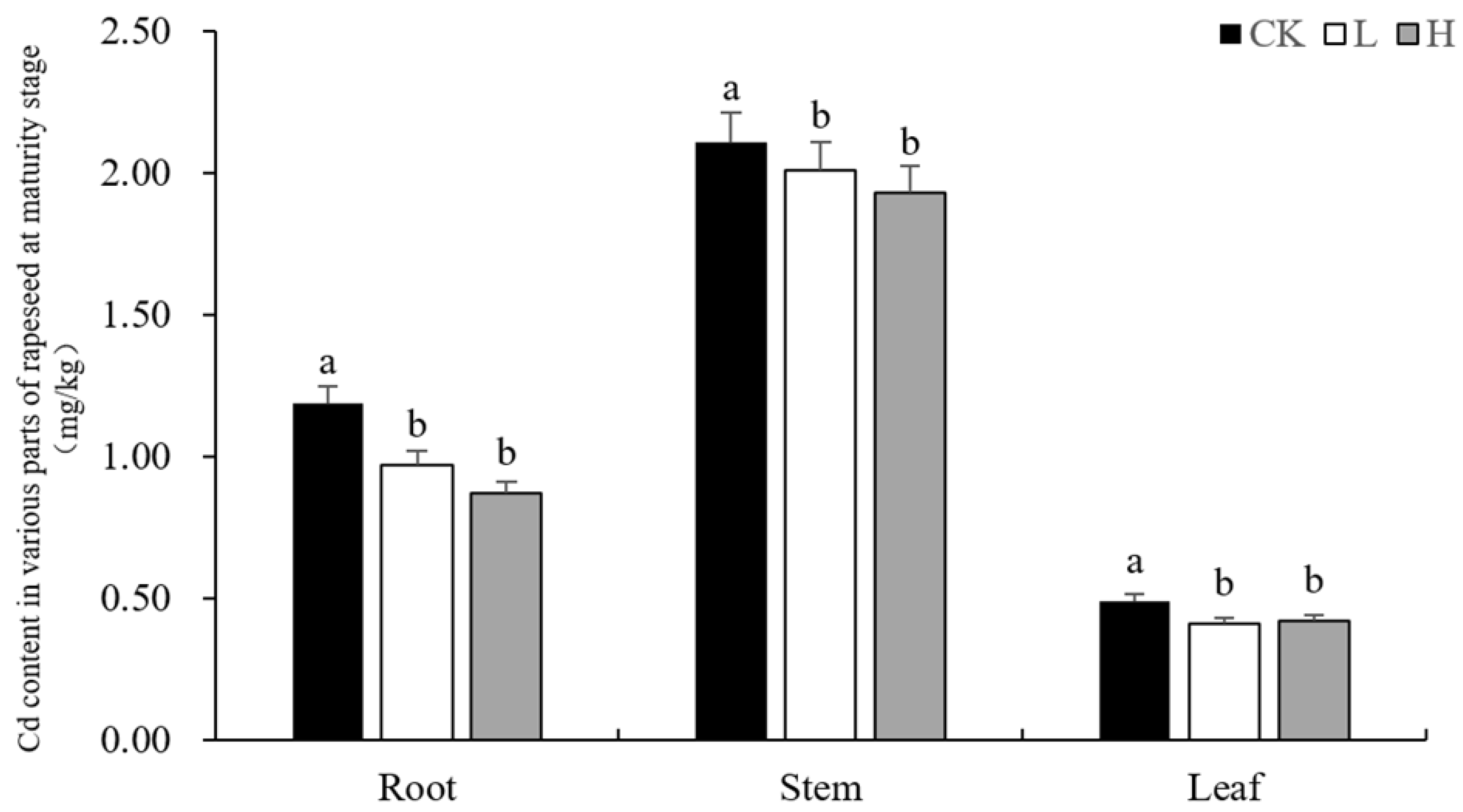
3.2. Effects of Different Amounts of Selenium on Cadmium Contents in Different Parts of Rapeseed
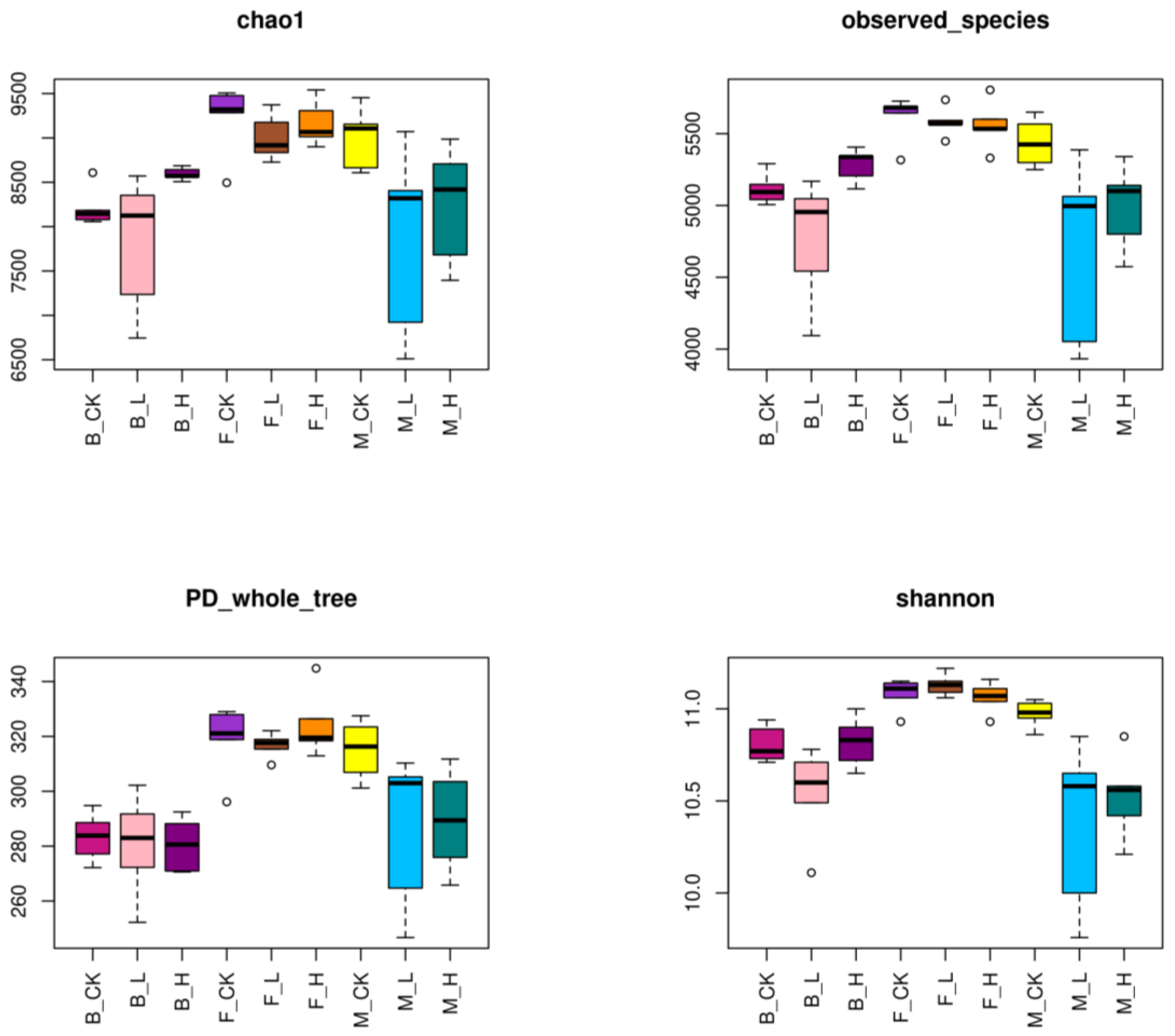
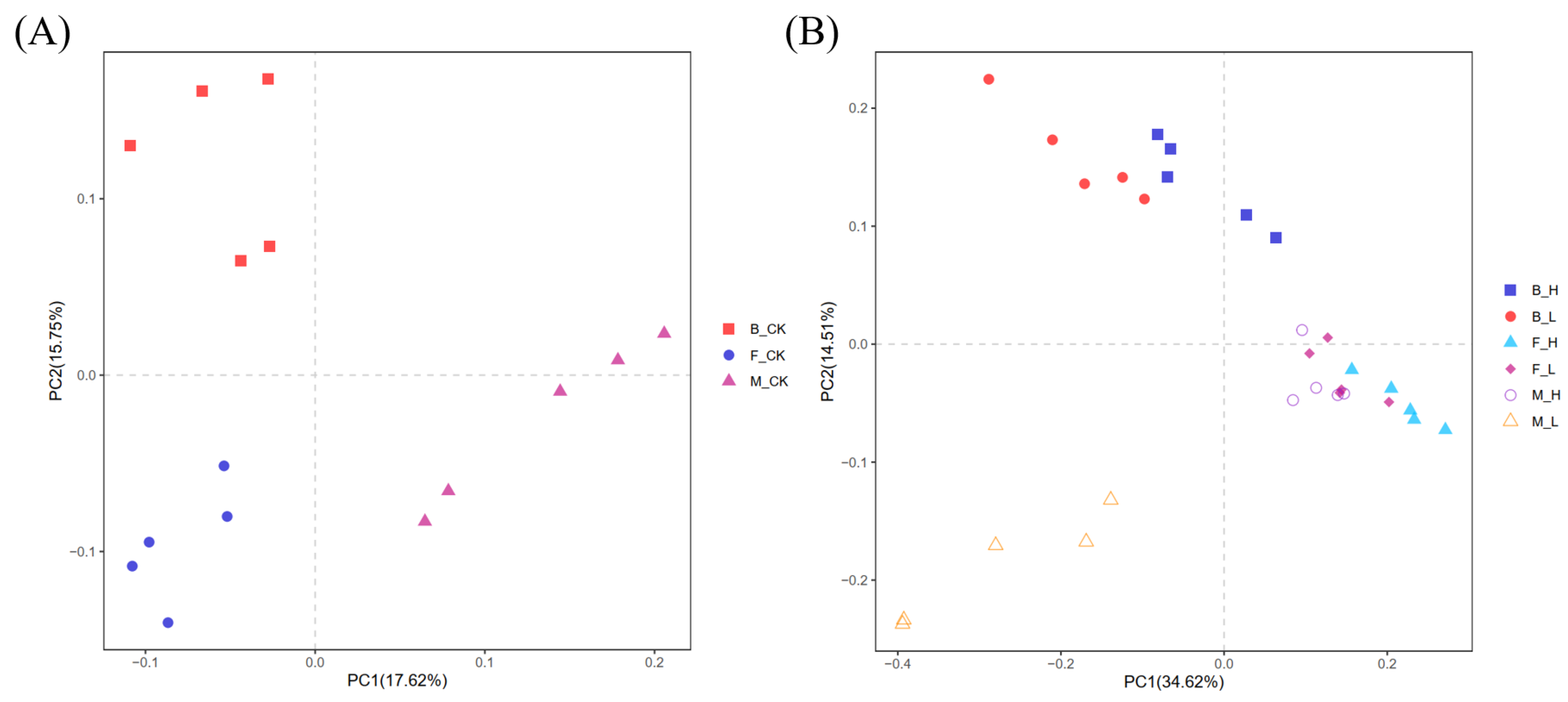
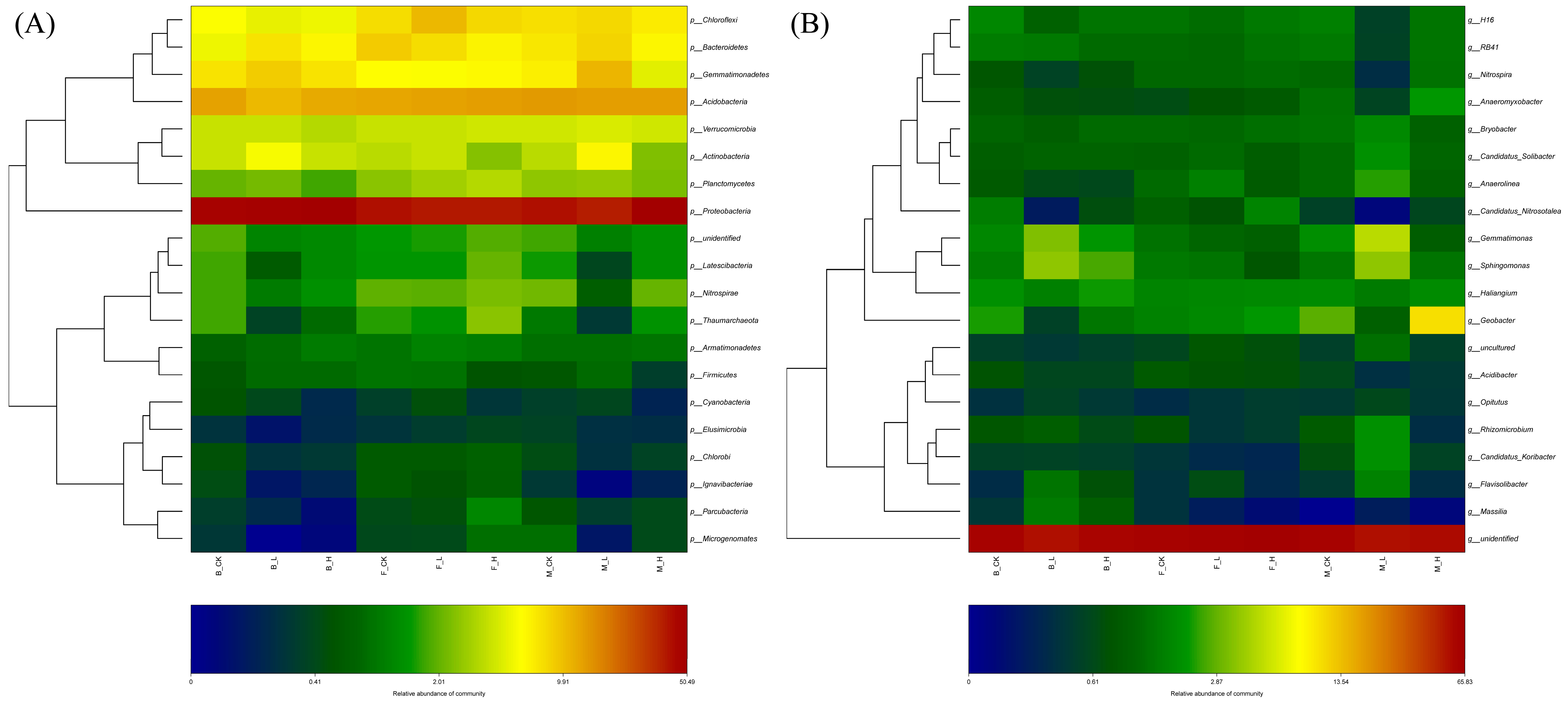
3.3. Diversity and Community Differences in Rhizosphere Microbial Communities
3.4. Effect of Root Selenium Treatment on Soil Bacterial Community Structure
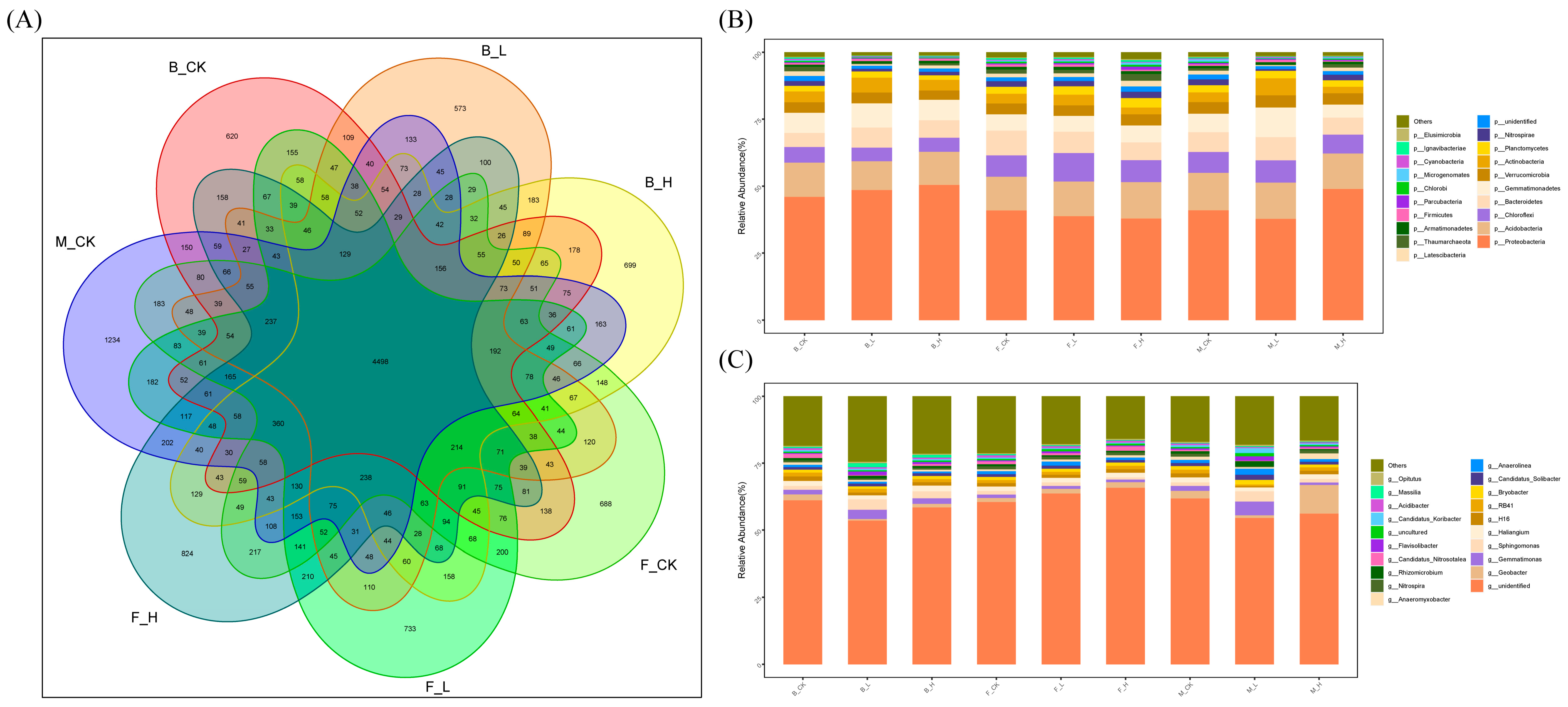
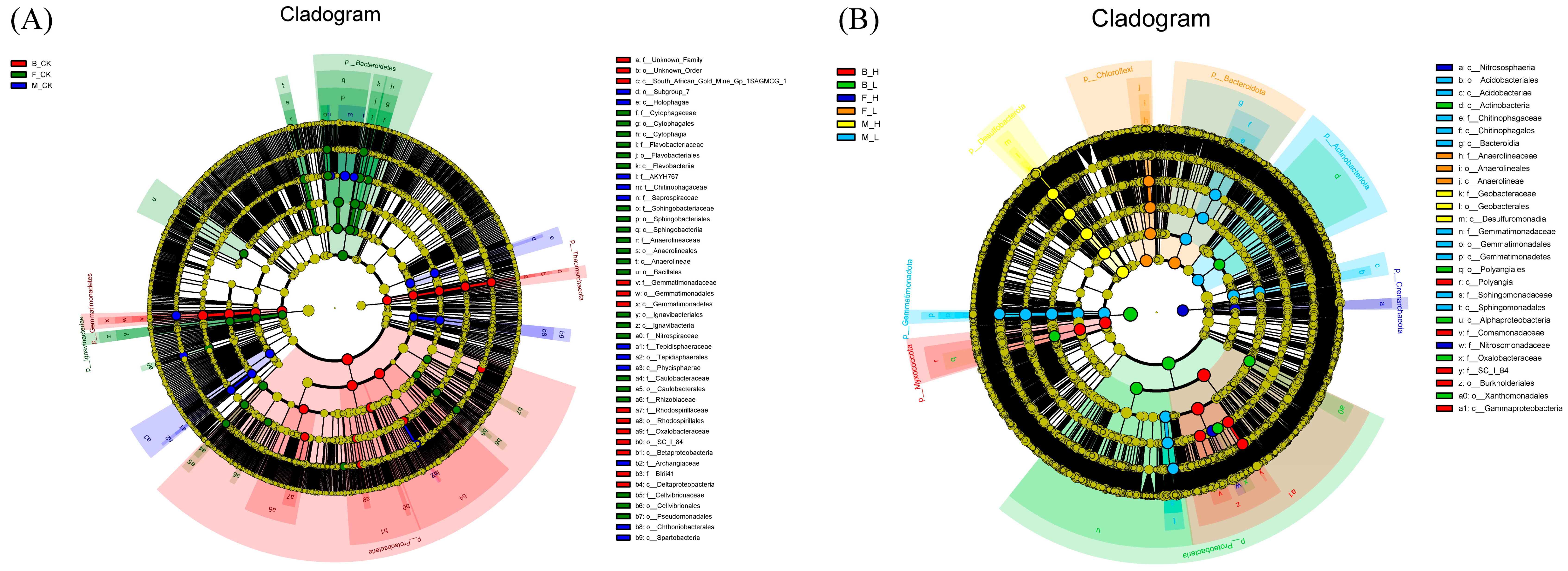
3.5. Effects of Non-Selenium Treatment on Different Soil Microbial Communities
3.6. Effects of Selenium Treatment on Different Soil Microbial Communities
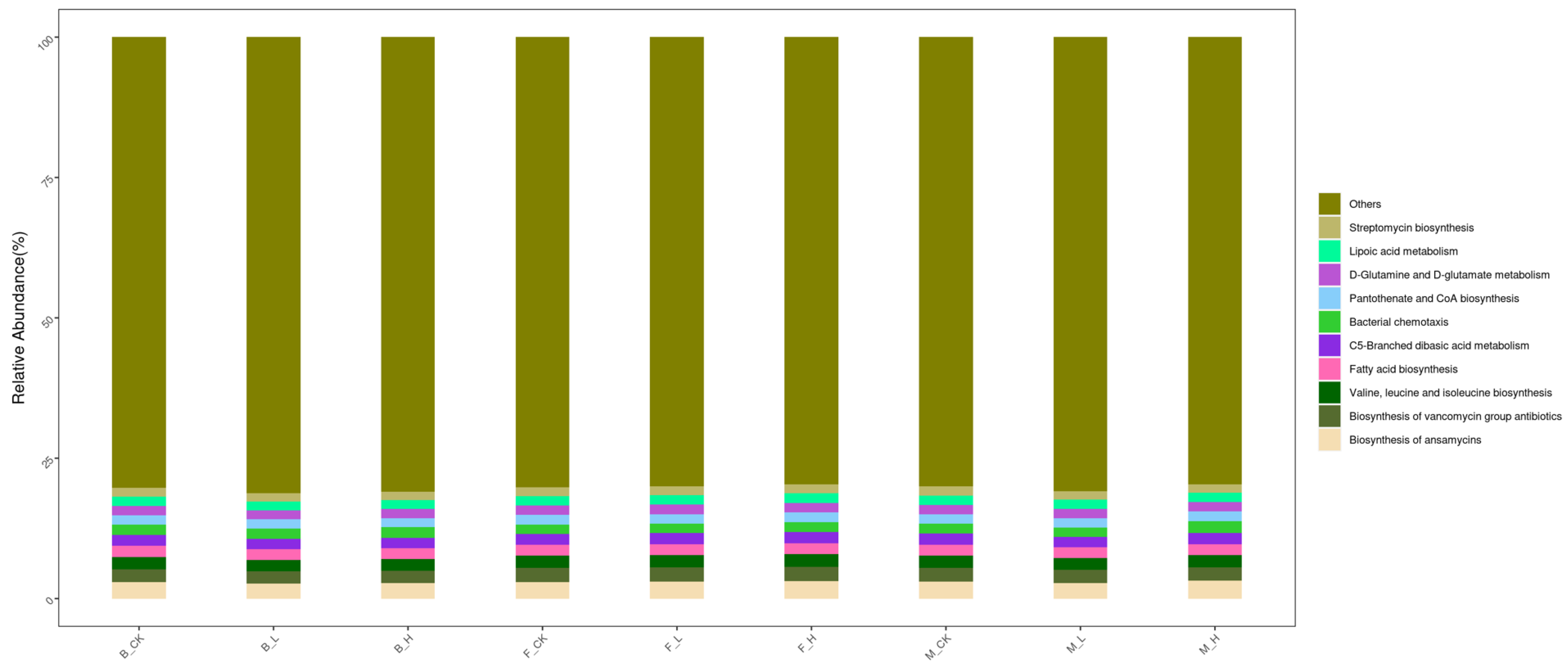
3.7. Effects of Selenium Treatment on Soil Metabolic Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Burger, J. Assessment and management of risk to wildlife from cadmium. Sci. Total Environ. 2008, 389, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Li, H.; Lu, J.; Bu, R.; Li, X.; Cong, R.; Lu, M. Crop rotation-dependent yield responses to fertilization in winter oilseed rape (Brassica napus L.). Crop J. 2015, 3, 396–404. [Google Scholar] [CrossRef]
- Weiser, C.; Fuß, R.; Kage, H.; Flessa, H. Do farmers in Germany exploit the potential yield and nitrogen benefifits from preceding oilseed rape in winter wheat cultivation? Arch. Agron. Soil Sci. 2018, 64, 25–37. [Google Scholar] [CrossRef]
- Leong, Y.K.; Chang, J.S. Bioremediation of heavy metals using microalgae: Recent advances and mechanisms. Bioresour. Technol. 2020, 303, 122886. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, N.; Raghuvanshi, R.; Upadhyay, M.K.; Srivastava, A.K.; Suprasanna, P.; Srivastava, S. Elemental (As, Zn, Fe and Cu) analysis and health risk assessment of rice grains and rice based food products collected from markets from different cities of Gangetic basin, India. J. Food Composit. Anal. 2020, 93, 103612. [Google Scholar] [CrossRef]
- Hu, Y.F.; Zhou, G.; Na, X.F.; Yang, L.; Nan, W.B.; Liu, X.; Zhang, Y.Q.; Li, J.L.; Bi, Y.R. Cadmium interferes with maintenance of auxin homeostasis in Arabidopsis seedlings. J. Plant Physiol. 2013, 170, 965–975. [Google Scholar] [CrossRef]
- Yang, G.L.; Zheng, M.M.; Tan, A.J.; Liu, Y.T.; Feng, D.; Lv, S.M. Research on the mechanisms of plant enrichment and detoxifification of cadmium. Biology 2021, 10, 544. [Google Scholar] [CrossRef]
- Yang, J.L.; Zhu, X.F.; Peng, Y.X.; You, X.; Zheng, C.; Li, G.X.; Liu, Y.; Shi, Y.Z.; Zheng, S.J. Cell wall hemicellulose contributes signifificantly to aluminum adsorption and root growth in Arabidopsis. Plant Physiol. 2011, 155, 1885–1892. [Google Scholar] [CrossRef]
- Zhao, H.D.; Du, L.Y.; Wu, Y.; Wu, X.N.; Han, W. Numerical assessment of the passivator efectiveness for Cd-contaminated soil remediation. Sci. Total Environ. 2021, 779, 146485. [Google Scholar] [CrossRef]
- Colin, Y.; Nicolitch, O.; Van NJ, D.; Zhou, J.Z.; Turpault, M.P.; Uroz, S. Taxonomic and functional shifts in the beech rhizosphere microbiome across a natural soil toposequence. Sci. Rep. 2017, 7, 9604. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, X.; Ji, B.; Struik, P.C.; Jin, K.; Tang, S. Coupling between the responses of plants, soil, and microorganisms following grazing exclusion in an overgrazed grassland. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Byers, A.K.; Condron, L.; O’Callaghan, M.; Waipara, N.; Black, A. Soil microbial community restructuring and functional changes in ancient kauri (Aga this australis) forests impacted by the invasive pathogen Phytophthora agathidicida. Soil Biol. Biochem. 2020, 150, 108016. [Google Scholar] [CrossRef]
- Fu, D.G.; Wu, X.N.; Duan, C.Q.; Zhao, L.Q.; Li, B. Different life-form plants exert different rhizosphere efects on phosphorus biogeochemistry in subtropical mountainous soils with low and high phosphorus content. Soil. Tillage Res. 2020, 199, 104516. [Google Scholar] [CrossRef]
- Liu, C.; Lin, H.; Li, B.; Dong, Y.; Yin, T. Responses of microbial communities and metabolic activities in the rhizosphere during phytoremediation of cd-contaminated soil. Ecotoxicol. Environ. Saf. 2020, 202, 110958. [Google Scholar] [CrossRef]
- Singh, J.P.; Vaidya, B.P.; Goodey, N.M.; Krumins, J.A. Soil microbial response to metal contamination in a vegetated and urban brownfeld. J. Environ. Manag. 2019, 244, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Xavier, J.C.; Costa, P.E.S.; Hissa, D.C.; Melo, V.M.M.; Falcão, R.M.; Balbino, V.Q.; Mendonca, L.A.R.; Lima, M.G.S.; Coutinho, H.D.M.; Verda, L.C. Evaluation of the microbial diversity and heavy metal resistance genes of a microbial community on contaminated environment. Appl. Geochem. 2019, 105, 1–6. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Pilon-Smits, E.A.; Zhao, F.J.; Williams, P.N.; Meharg, A.A. Selenium in higher plants: Understanding mechanisms for biofortifification and phytoremediation. Trends Plant Sci. 2009, 14, 436–442. [Google Scholar] [CrossRef]
- Natasha; Shahid, M.; Niazi, N.K.; Khalid SMurtaza, B.; Bibi, I.; Rashid, M.I. A critical review of selenium biogeochemical behavior in soil-plant system with an inference to human health. Environ. Pollut. 2018, 234, 915–934. [Google Scholar] [CrossRef]
- Schiavon, M.; Pilon-Smits, E.A. The fascinating facets of plant selenium accumulation—Biochemistry, physiology, evolution and ecology. New Phytol. 2017, 213, 1582–1596. [Google Scholar] [CrossRef]
- Li, C.; Hu, C.; Xie, J.; Shi, G.; Wang, X.; Yuan, X.; Li, K.; Chen, S.; Zhao, X.; Fan, G. Selenium Combined with Methyl Jasmonate to Control Tomato Gray Mold by Optimizing Microbial Community Structure in Plants. J. Fungi 2022, 8, 731. [Google Scholar] [CrossRef]
- Zhang, M.; Tang, S.; Huang, X.; Zhang, F.; Pang, Y.; Huang, Q.; Yi, Q. Selenium uptake, dynamic changes in selenium content and its inflfluence on photosynthesis and chlorophyll flfluorescence in rice (Oryza sativa L.). Environ. Exp. Bot. 2014, 107, 39–45. [Google Scholar] [CrossRef]
- Filek, M.; Keskinen, R.; Hartikainen, H.; Szarejko, I.; Janiak, A.; Miszalski, Z.; Golda, A. The protective role of selenium in rape seedlings subjected to cadmium stress. J. Plant Physiol. 2008, 165, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Griffifiths, B.S.; Philippot, L. Insights into the resistance and resilience of the soil microbial community. FEMS Microbiol. Rev. 2013, 37, 112–129. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.W.; Wei, C.Y.; Tu, S.X.; Ding, Y.Z.; Song, Z.G. A dual role of Se on cd toxicity: Evidences from the uptake of cd and some essential elements and the growth responses in paddy rice. Biol. Trace Elem. Res. 2013, 151, 113–121. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. Adv. Online Publ. 2013, 7, 1069–1079. [Google Scholar] [CrossRef]
- Wang, Y.; Sheng, H.-F.; He, Y.; Wu, J.-Y.; Jiang, Y.-X.; Tam, N.F.-Y.; Zhou, H.-W. Comparison of the Levels of Bacterial Diversity in Freshwater, Intertidal Wetland, and Marine Sed iments by Using Millions of Illumina Tags. Appl. Environ. Microbiol. 2012, 78, 8264. [Google Scholar] [CrossRef]
- Hussain, B.; Ashraf, M.N.; Shafeeq-Ur-Rahman Abbas, A.; Li, J.; Farooq, M. Cadmium stress in paddy fields: Effects of soil conditions and remediation strategies. Sci. Total Environ. 2021, 754, 142188. [Google Scholar] [CrossRef]
- Li, W.; Xu, B.; Song, Q.; Liu, X.; Xu, J.; Brookes, P.C. The identification of ‘hotspots’ of heavy metal pollution in soil-rice systems at a regional scale in eastern China. Sci. Total Environ. 2014, 472, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Cheng, H.; Tao, S. The Challenges and Solutions for Cadmium-contaminated Rice in China: A Critical Review. Environ. Int. 2016, 92–93, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Sehrish, A.K.; Aziz, R.; Hussain, M.M.; Rafiq, M.T.; Rizwan, M.; Muhammad, N.; Rafiq, M.K.; Sehar, A.; Din, J.U.; Al-Wabel, M.I.; et al. Effect of poultry litter biochar on chromium (Cr) bioavailability and accumulation in spinach (Spinacia oleracea) grown in Cr-polluted soil. Arab. J. Geosci. 2019, 12, 57. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets--iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, S.; Jiang, Z.; Wang, Y.; Zhang, Z. Selenium Biofortification Modulates Plant Growth, Microelement and Heavy Metal Concentrations, Selenium Uptake, and Accumulation in Black-Grained Wheat. Front. Plant Sci. 2021, 12, 748523. [Google Scholar] [CrossRef]
- De Feudis, M.; Massaccesi, L.; D’Amato, R.; Businelli, D.; Casucci, C.; Agnelli, A. Impact of Na-selenite fertilization on the microbial biomass and enzymes of a soil under corn (Zea mays L.) cultivation. Geoderma 2020, 373, 114425. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Ma, J.; Wu, F.; Wang, L.; Sun, L.; Zhang, P.; Wang, W.; Xu, J. Comparative physiological and soil microbial community structural analysis revealed that selenium alleviates cadmium stress in Perilla frutescens. Front. Plant Sci. 2022, 13, 1022935. [Google Scholar] [CrossRef]
- Rudi, K.; Berg, F.; Gaustad, E.; Tannes, T.; Vatn, M. Ratios between alpha-, beta- and gamma-proteobacteria in tap water determined by the ProteoQuant assay. Lett. Appl. Microbiol. 2010, 50, 1–6. [Google Scholar] [CrossRef]
- Liu, K.; Cai, M.; Hu, C.; Sun, X.; Cheng, Q.; Jia, W.; Yang, T.; Nie, M.; Zhao, X. Selenium (Se) reduces Sclerotinia stem rot disease incidence of oilseed rape by increasing plant Se concentration and shifting soil microbial community and functional profiles. Environ. Pollut. 2019, 254 Pt B, 113051. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Silicle Number per Plant | Seeds per Silicle | Seed Weight (mg) | Yield (kg/hm2) |
|---|---|---|---|---|
| CK | 610.4 ± 29.5 b | 23.26 ± 0.81 a | 2.92 ± 0.18 a | 2280.18 ± 82.51 b |
| L | 676.4 ± 22.1 a | 24.38 ± 0.59 a | 3.04 ± 0.23 a | 2757.23 ± 148.21 a |
| H | 611.8 ± 4.9 b | 23.53 ± 0.12 a | 2.94 ± 0.25 a | 2327.78 ± 40.43 ab |
| Sample | Raw_Tags | Clean_Tags | Coverage |
|---|---|---|---|
| B_CK | 105,498 | 101,630 | 96.33 |
| B_H | 361,809 | 343,433 | 94.92 |
| B_L | 91,700 | 89,008 | 97.06 |
| F_CK | 124,037 | 120,711 | 97.32 |
| F_H | 185,158 | 181,078 | 97.80 |
| F_L | 61,673 | 60,132 | 97.50 |
| M_CK | 512,856 | 496,359 | 96.78 |
| M_H | 216,585 | 210,290 | 97.09 |
| M_L | 350,507 | 336,036 | 95.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, L.; Zhou, B.; Hong, B.; Wang, X.; Chang, T.; Guan, C.; Guan, M. Application of Selenium Can Alleviate the Stress of Cadmium on Rapeseed at Different Growth Stages in Soil. Agronomy 2023, 13, 2228. https://doi.org/10.3390/agronomy13092228
Nie L, Zhou B, Hong B, Wang X, Chang T, Guan C, Guan M. Application of Selenium Can Alleviate the Stress of Cadmium on Rapeseed at Different Growth Stages in Soil. Agronomy. 2023; 13(9):2228. https://doi.org/10.3390/agronomy13092228
Chicago/Turabian StyleNie, Lingli, Bingqian Zhou, Bo Hong, Xiaodan Wang, Tao Chang, Chunyun Guan, and Mei Guan. 2023. "Application of Selenium Can Alleviate the Stress of Cadmium on Rapeseed at Different Growth Stages in Soil" Agronomy 13, no. 9: 2228. https://doi.org/10.3390/agronomy13092228