
Optimization of Conditions for In Vitro Culture of Selected Arbutus unedo L. Genotypes



1
Department of Horticulture, Agriculture Faculty, Kocaeli University, Kocaeli 41100, Turkey
2
Kocaeli Directorate, Ministry of Agriculture and Forestry, Kocaeli 41100, Turkey
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(3), 623; https://doi.org/10.3390/agronomy12030623
Submission received: 9 December 2021
/
Revised: 22 February 2022
/
Accepted: 25 February 2022
/
Published: 3 March 2022
Abstract
:The strawberry tree is an evergreen shrub tree that grows on sea-facing slopes under a Mediterranean climate. It has a key role in the lives of the rural inhabitants of forested areas and is one of the most promising species for developing new approaches in cultivation for improved fruit. As a result of the growing interest of farmers, selection studies have focused on the selection of highly productive varieties of Arbutus unedo, whereby there is a requirement for the selected varieties to be propagated at a large scale. In this study, the in vitro propagation potential of the selected A. unedo genotypes, YV2 and YV4, was investigated. The best results in terms of decreasing browning with a high level of sterilization were achieved with 50% ethanol (1 min) and 20% NaCIO (12 min) for both shoot tip and nodium explants. In terms of growth performance, both genotypes displayed the highest proliferation rate in WPM medium-supplemented with BAP (2.0 mg/L) and with added IBA and GA3 (0.1 mg/L each), in which the shoot number and elongation were also the highest. YV2 microcuttings showed higher rooting rates when cultured in WPM medium, with an added 2.0 mg/L IBA, whereas the addition of 4.0 mg/L NAA was optimal for YV4. The resulting root quality was sufficient for surviving the acclimatization stage. The genotype effect was clear in all stages during the in vitro propagation for all growth parameters.
1. Introduction
The strawberry tree (Arbutus unedo L., Ericaceae family) is an evergreen shrub tree belonging to the Mediterranean maquis. Plants grow spontaneously on the sea-facing slopes that are characterized by hot summers and mild and rainy winters. The strawberry tree is a sclerophyllous evergreen shrub that prefers siliceous soils and grows in altitudes of up to 800 m. It has a wide continental distribution area that includes the western coast of North America and Central America, Eastern Europe, the Mediterranean region, North Africa, and parts of the Middle East. In Turkey, it is distributed on all seashores in the maquis forest areas, such as along the Aegean Sea, the Sea of Marmara, the Black Sea, and in Mediterranean maquis. It is a hardy plant, and this plays a key role in the reforestation programs of areas where forest fires are common during the dry season [1]. It produces red spherical fruits, which have a diameter of about 2–3 cm [2]. These berries are desirable for fresh consumption or processed as jams, candies, or liqueurs [3], and the wood of the plant can be used as firewood and to prepare charcoal [4]. This species has also been used for ornamental purposes, with its flowers and fruit structures being considered an attractive addition in landscaped areas. Knowledge of the healthy properties of Arbutus unedo fruits has an ancient history, so they have been used in folk medicine for many years [5]. The fruit has received increasing interest because of its common traditional, industrial, and pharmaceutical uses [6,7]. The fruits of the plant have been traditionally used as antiseptics, diuretics, and laxatives in folk medicine. Different extracts obtained from Arbutus unedo L. fruits and leaves have high pharmacological potential and can be used for the development of new drugs to treat diseases such as diabetes and hypertension, among others [5]. Arbutus unedo fruits have high amounts of phenolic contents and display antibacterial effects. When compared with other berries, the fruits show higher amounts of vitamin C, mainly in the ascorbic acid form. Therefore, Arbutus unedo fruits should be considered one of the new important sources of safe antioxidants [8,9,10]. The use of A. unedo extracts as bio-preservatives in the functional food industry has been suggested [11]. In recent years, minor fruits, especially berries, have been of increasing interest.
As mentioned, Arbutus unedo is widely distributed all over the world, with its wide distribution in areas with a Mediterranean climate. The results of a number of studies focusing on the genetic diversity of natural populations of Arbutus unedo have shown that there is a wide biodiversity within the populations [12,13,14]. Arbutus unedo trees have lignotubers at the stem base, and this structure is viewed as providing the plant with wide ecological potential as a native fruit of the Mediterranean ecosystem. It helps to stabilize soils and prevent erosion; it also has a strong regeneration capacity following fire, and survives in drought conditions and poor soil types [15,16]. Lignotubers are also important storage organs for carbohydrates and, thus, play an important role in resprouting after forest fires or in the recovery process after disturbances [17].
The Food and Agriculture Organization (FAO) is currently focused on the ways in which to increase and support the culturing of valuable wild species [18], with the aim of preserving plant biodiversity. Among these species, Arbutus unedo springs to attention due to its high distribution and evaluation possibilities. The cultivation of wild species, which can generally be collected from nature and evaluated, is the most important step for the protection of biological destruction. In this case, the need for quality propagation material is essential for establishing healthy gardens. In subsequent breeding work, researchers should pay attention to the capacity to ease and standardize the propagation capacity of plants; and this will not only facilitate the planting of modern orchards, but may also reduce the cost of afforestation studies. The rooting rate of Arbutus unedo L. cuttings is moderately low and changeable with regard to genotype and maturity [19,20]. The propagation of selected strawberry genotypes is very important. Seed propagation results in genetic differences cannot be used to obtain uniform seedling materials. The frequency of rooting when using cuttings varies with genetic structure and can be very low in some cases [20,21]. In vitro culture is a good alternative method to obtain bulk material when conventional methods are insufficient, and when rejuvenation problems persist. The determination of in vitro propagation protocols for the rapid multiplication of the material and to obtain disease-free plants for specific genotypes is very important. Thus, this paper examines the in vitro propagation capacity of the strawberry tree and conditions for its optimization.
2. Materials and Methods
In the research, YV2 and YV4 strawberry tree genotypes, which were included in the selection study conducted by [12], were used as plant material. Woody shoots from naturally grown plants in the forest area of Kocaeli Samanlı Mountain in Turkey were collected before the flowering stage in the autumn of 2016, during September and October (Figure 1). Shoots with lengths of 40–50 cm and diameters of 3–5 cm were washed under running tap water and then placed into a big plastic bucket filled with tap water. Buckets were placed in the laboratory at 20–25 °C, under a 16 h photoperiod provided by cool-white fluorescent light with a light intensity of 3000 lux to induce the sprouting of epicormic shoots.
Newly grown apical shoots and nodal explants with axillary buds were collected when they reached a length of 5–10 cm, for use in culture establishment (Figure 2). All experimental studies were conducted in the research laboratory of the Agriculture Faculty at Kocaeli University between 2016 and 2018.
2.1. Determination of the Sterilization Protocol
In this stage, shoot tips and nodal segments of a strawberry tree of about 1.0–1.5 cm in length were collected from sprouted epicormic shoots. These explants were washed in running tap water for 30 min to decrease the microorganism density. Then, the sterilization procedures given in Table 1 were applied. The explants were immersed in 0%, 50%, or 70% ethanol for 1 or 2 min under aseptic conditions.
After the alcohol was removed via dipping the explants in sterile distilled water for 5 min, the explants were shaken in the 20% sodium hypochlorite (NaCIO) solution containing two added drops of Tween 20 (C58H114O26) for 10, 12, or 15 min, and then rinsed three times with sterile distilled water to remove all traces of disinfectant. In this stage, sterilized shoot tips and internodes were cultured on a hormone-free MS medium. The culturing was conducted in 10 mL test tubes containing 10 mL of the nutrient medium. After ten days, cultures were evaluated for contamination and the enzymatic browning of tissues.
2.2. Culture Establishment
Epicormic shoots were collected from woody shoots. Shoot tips and nodal segments were also used as starter materials due to having limited material. Sterilization was conducted in two stages. First, materials were dipped in a 50% ethanol solution for 1 min and rinsed with sterile distilled water for 5 min to remove the alcohol. Then, both types of material were rinsed in a 20% sodium hypochlorite solution with two added drops of Tween 20 for 12 min. Explants were cultured on the hormone-free MS medium, and 7 g/L agar and 30 g/L sucrose were added. The pH values of the media were adjusted to 5.7 by the addition of 0.1 N NaOH or 0.1 N HCL. The culturing was conducted in 10 mL culture tubes containing 10 mL of nutrient medium. The culture medium was changed every 3 days to decrease the enzymatic browning effect of phenolics.
2.3. Proliferation Stage
Healthy shoot tips and internodes that were obtained in the establishment stage were cultured for shoot proliferation. Two different nutrient media were used in the proliferation stage experiments: the full-strength MS (Murashige and Skoog) [22] and full-strength WPM (woody plant medium) [23]. MS or WPM media were supplemented with different concentrations of BAP (0.0, 0.5, 1.0, 2.0, or 4.0 mg/L), IBA (0.1 or 0.5 mg/L), and GA3 (0.1 mg/L) for inducing shoot proliferation, and they were enriched with 30 g/L sucrose and gelled with agar (7 g/L). The pH was adjusted to 5.7 ± 0.1 before autoclaving. The cultures were incubated at 25 ± 1 °C under a 16 h photoperiod with a photosynthetic photon at 4000 lux light illumination, provided by white fluorescent tubes. The appearance of shoots was taken into consideration in the frequency of the culture period (4–6 week intervals). Three Erlenmeyer flasks (250 mL volume, containing 50 mL of the nutrient medium) containing eight explants formed one treatment, with each treatment carried out in triplicates. Shoot tips (≥0.5 cm) obtained from cultures were transferred to new culture media for the induction of multiple shoots. The proliferation rate and the average number of shoots induced per explant, as well as the length of shoots, were recorded before each transfer. The percentage of shoot multiplication (%), the average number of shoots per explant, and the average shoot length (cm) were recorded in this stage.
2.4. Rooting Stage
Healthy shoots (≥1.5 cm) obtained in the proliferation stage were transferred to the WPM medium for the rooting stage. In the rooting experiments, the WPM full-strength medium was supplemented with IBA or NAA (1.0, 2.0, and 4.0 mg/L doses for each auxin) to induce the rooting initiation and root formation of microshoots.
Rooting media were solidified by 0.7% agar and 3% sucrose added prior to autoclaving at 121 °C and 1.2 kg/cm2 for 20 min. The pH values of the media were adjusted to 5.7 by the addition of 0.1 N HCL or 0.1 N NaOH. Cultures were evaluated at the end of 40 days. The percentage of rooting (%), the average root number, and the average root length were recorded in rooting stage.
2.5. Acclimatization Stage
Microshoots with well-developed roots were taken out of the test tubes and thoroughly washed with sterile distilled water to remove any remaining medium. Plantlets were transferred to plastic pots (5 cm diameter) containing a combination of autoclaved peatmoss and perlite at 1:1 (v/v); each of the potted plantlets were covered with transparent polythene bags to supply a high-humidity atmosphere. They were irrigated with sterile distilled water when needed and grown under 16 h light/8 h dark at 24 ± 1 °C. Initially, the cups were incubated in the culture room under a 4000 lux light regime at a temperature of 25–28 °C for about 20 days. In order to acclimate to the environmental conditions, the plastic bags covered on plantlets were cut gradually, and the humidity was decreased step by step by puncturing the plastic cover day by day. Data on acclimatization were recorded after 20 days in greenhouse conditions and the percentage of surviving microplants was recorded. The success of the hardening protocol was determined by calculating the survival percentage.
2.6. Statistical Analysis
Each experiment was set up using a completely randomized design with three replicates. In the sterilization stage, 15 explants were taken for each replicate. Establishment experiments were obtained with eight shoot tips or nodiums in each replicate. Healthy shoots were taken for the proliferation stage with eight explants in each replicate. The proliferation stage was repeated two times, and mean values of the results were used for statistical analysis. The qualitive data obtained as percentages were first submitted to arcsine transformation and used in statistical analysis. Rooted shoots were transferred for acclimatization, and the results were given as a percentage because the raw numbers were not equivalent for comparison in statistical analysis. Statistical analysis was performed by an analysis of variance (ANOVA) using Minitab version 13.0 (S0064 Minitab Release 13, Lisance No: wcp 1331.00197, Trakya University, Edirne-Turkey). MinitabMeans were separated according to a Tukey test at a probability of p < 0.05.
3. Results and Discussion
3.1. Sterilization Stage
The establishment of aseptic tissues involves a sterilizing protocol. This is a very critical point for the next stage in the healthy development of tissue cultures [24]. In this study, different sterilization treatments were conducted to investigate the effect of sterilizing agents, the type of starting material on the percentage of live explants, and the percentage of contaminated explants. The sterilization of both types of material (shoot tips and nodal segments) increased in strength with the addition of higher concentrations of sodium hypochlorite in the ethanol solutions. The effect of sterilization treatments is given in Table 1. NaCIO was not effective at a desirable level when used alone, i.e., undiluted. The timing of EtOH exposure is another important factor determining the survival of explants. A pre-sterilization of 2 min resulted in higher explant death rates. When all parameters were evaluated together, the best results were for 50% EtOH (1 min) + 20% NaCIO (12 min) treatment, which gave satisfactory results in terms of achieving nearly complete sterilization of the materials. The infection and browning formation of both the shoot tips and nodiums were at a minimum level in this treatment (Table 1).
In the literature, several chemicals are used for the sterilization of culture starter materials at different concentrations and times of immersion. The use of NaClO for the surface sterilization of explants has been reported in many previous studies because of its simplicity, effectiveness, and low cost. NaClO is also a highly effective sterilant against all kinds of bacteria, fungi, and viruses. Increasing the NaClO concentration and exposure time resulted in low survival percentages, and, on the other hand, contamination increased with lower concentrations of NaCIO [25,26,27]. NaClO is usually applied with other chemicals in sterilization. Damiano et al. [28] reported that using a NaClO and Na merthiolate combination for 20 min could successfully sterilize axillary bud segments of pomegranate and mulberry.
A two-step sterilization process with ethanol was found to be effective in a previous study [25,27,29], which is in harmony with our findings. In our study, EtOH with NaClO also provided the best results for all observed parameters. On the other hand, EtOH is known to be a powerful, yet phytotoxic sterilizing agent, so we saw the same effect with a two-minute application. However, the treatment with 70% EtOH (2 min) and 20% NaCIO (15 min) for the surface sterilization of nodal segments of Arbutus pavarii was found to give satisfactory results in that study, which is in contrast with our results, in which the 2 min immersion time in EtOH had a negative effect on the survival of Arbutus unedo genotype explants examined in this study.
As a satisfactory result of the sterilization process, both the shoot tips and nodal parts of epicormic shoots were used for culture establishment. Sterilization was made with a pretreatment with 50% EtOH for 1 min and rinsed with sterile distilled water to remove detergent residues. Then, in the second step of sterilization, explants were shaken with 20% NaCIO for 12 min, followed by rinsing in sterile distilled water three times (each for 5 min), after which they were ready to culture.
3.2. Establishment Stage
The structure of starter explants is one of the keys determining the success in establishing cultures. The establishment cultures were performed using shoot tips and nodiums of YV2 and YV4 genotypes, which were cultured in a hormone-free MS medium. Shoot tip survival rates were higher for shoot tip explants of both genotypes; however, this difference was not significant. The highest survival rate was observed with YV2 genotype shoot tips (93.33%), whereas browning formation occurred at a minimum level. The YV4 genotype shoot and nodium explants showed similar results in terms of survival and recorded higher rates of browning. With this genotype, the death of explants increased (Table 2). Even though the establishment stage cultures were grown in a hormone-free MS medium, axillary buds sprouted. We thought that this may be due to the nodium being richer than shoot tips in terms of nutrition content (Figure 3). Therefore, axillary buds sprouted readily after they were planted in a culture medium. No proliferation occurred for shoot tip explants of either genotype, and only elongation was observed. The number of new shoots was higher when using the nodium parts of YV2, and the shoot length was slightly longer. Both types of explant sources have been used in previous studies. In some studies, the data indicate that shoot apices are more effective for shoot productivity in the establishment stage than nodal segments [30]. On the other hand, nodium parts are more common in woody plants and were successfully used for culture initiation in other previous studies [29,31,32,33] and subjected to further proliferation. In previous studies of Arbutus unedo, the axillary shoots and nodiums were used as starter material, for which satisfactory results for culture establishment were obtained [34,35]. Gomes and Canhato [36] performed an in vitro culture of Arbutus unedo from shoot tips and nodiums. As with our results, nodium explants showed precocious proliferation compared to Musa spp. shoot tips [37].
In the study, higher amounts of browning in the nodal parts may be due to a high content of phenolic substances and browning activities. The discharge of phenolic compounds into in vitro culture media is a frequently encountered problem. Phenolic compounds are the main reason for lethal browning reactions in the in vitro cultures and are responsible for high mortality rates [32,38,39]. To prevent or decrease the effect of internal plant phenolics, frequent changing the culture medium was suggested. In our study, transferring the explants to a new nutrient medium every three days reduced blackening to reasonable limits for Arbutus unedo. Plant materials were collected from outdoor plants in autumn just before the flowering season, and epicormic shoots were stimulated. We think that our materials perhaps had higher levels of phenolic substances due to the seasonal variation. Cao et al. [40] reported a significant seasonal variation in the levels of phenolic compounds and antioxidant activity in species, i.e., Cyclocarya paliurus.
In culture establishment, the percentage of proliferation and new shoot numbers was very low. However, the shoot quality was sufficient for subcultures, which were successfully transferred for proliferation after 30 days.
3.3. Proliferation Stage
Using the appropriate type and concentration of plant growth regulators is important in controlling the growth and morphogenesis of microshoots.
The highest proliferation rate and longest shoots of YV2 were obtained with the WPM nutrient medium fortified with 2.0 mg/L of BAP + 0.1 mg/L of IBA + 0.1 mg/L of GA3 (100% and 2.03 cm, respectively). Although the WPM medium with 1.0 mg/L of BAP produced the highest shoot number (2.32 per explant), 2.0 mg/L was the optimal concentration overall in both nutrient media among the tested concentrations of BAP when considering other growth parameters (Table 3). Higher concentrations of growth regulators resulted in decreased proliferation rate and shoot length. Shoot proliferation was negatively affected in the MS medium supplemented with GA3 and IBA, and resulted in the formation of more shoots with higher levels of browning. The addition of higher concentrations of IBA and GA3 to the MS medium similarly resulted in a further increase in browning (Figure 4).
In cultures of YV4 during the proliferation stage on the WPM medium, supplemented with various concentrations of BAP, 2.0 mg/L of BAP produced the highest shoot proliferation in comparison with the MS medium, although this difference was not statistically significant from the rates for other WPM medium formulations. The WPM medium was supplemented with 2.0 mg/L of BAP and 0.1 mg/L of IBA, which gave the highest proliferation rate (86.67%) for YV4, as was the case for genotype YV2 (Table 4). Shoot number and shoot length were also the highest in this medium. The inhibitory effect of higher BAP concentrations was observed in genotype YV4, and the MS medium with 6.0 mg/L BAP added gave the shortest shoots and highest browning percentage (Table 4). IBA induced the browning of shoots; browning formation was significantly increased in the MS medium with the addition of 0.5 mg/L of IBA, when compared with the MS medium that contained 0.1 mg/L of GA3 with one of the BAP concentrations (Figure 5).
Genotype, nutrient medium, and medium composition are very important factors for the induction and growth of new shoots from both of the studied genotypes. The microelement concentrations of the MS medium were higher, therefore, being a richer medium than the WPM medium. In particular, the contents of the nitrate form of nitrogen and calcium were higher. Previously, researchers have suggested that the type and amount of nitrogen may affect the growing habits of shoots [41]. In previous studies, Arbutus unedo genotypes exhibited various growing habits in different nutrient media. The half-strength MS medium was not satisfactory for Arbutus unedo, whereas the WPM medium provided good results. The effects of the media changed according to the genotypes. The effects of the MS medium were altered in the proliferation stage by varying IBA and GA3 concentrations. A total of 0.1 mg/L of IBA and 0.1 mg/L of GA3 provided more satisfactory results for both of the two genotypes. Increasing the concentration of IBA in culture media induced browning formation, especially when used with 0.25 mg/L of GA3. Gomes and Canhato [36] reported significant effects of BA on shoot length. In proliferating cultures, the shoot number and length were not at satisfactory levels for either genotype. The lower concentrations of BAP were insufficient for shoot multiplication, whereas higher concentrations prevent shoot elongation. In our study, a significant decrease in shoot proliferation rates was noticed in the MS medium supplemented with 1.0 mg/L of BA, 0.5 mg/L of IBA, and 0.25 mg/L of GA3. When the medium was supplemented with 0.5 mg/L of IBA, browning formation was stimulated, and shoots died because of browning in both genotypes. Similar effects were reported in other species with higher doses of IBA [32]. In in vitro hazelnut cultures, supplementation of the MS medium with 1 or 2 mg/L of IBA increased browning formation on shoot apices [42]. The MS medium free from IBA gave more satisfactory results. The shoot proliferation in cultures indicate the function of cytokinins. The manipulation of cytokinin and auxin concentrations combined with gibberellin directly affected the proliferation. In our study, explant proliferation was induced with increasing BAP concentrations (up to 4.0 mg/L doses of BAP), whereas higher concentrations of BAP had negative effects on culture success and shoot quality. This result was in accordance with the results for Arbutus andrachne in [43]. On the contrary, another study conducted with Arbutus unedo showed that increasing doses of BAP clearly increased the number of shoots [35], as in our studies. They showed that the multiplication rate increases with the amount of BAP, whereas the rate of elongation was found to decrease in another study [27]. However, some other researchers found that increasing doses of BAP caused decreasing proliferation rates with production of only a few small microshoots [32].
We observed that the YV2 exhibited proliferation performance superior to YV4. The measured data show that shoot formation and growing characteristics were influenced by the genotypes, as suggested in previous studies [12,44]. In Arbutus pavarii cultures, kinetin was found to be more effective than BA, whereas Gomes and Canhato [36] suggested BAP was the optimal cytokinin source for Arbutus unedo cultures.
In previous studies, genotype effects were found to be important in the in vitro culture of Arbutus unedo [36]. This conclusion is in harmony with our results. Some Arbutus clones did not produce sufficiently long shoots for the rooting stage. Here, both genotypes produced shoots tall enough to progress to the rooting stage. Similarly, A. andrachne and A. unedo shoots successfully proliferated, and micropropagation was suggested as a good tool for these species [31,34,45]. In particular, the role of genotypes on proliferation capacity was declared in previous results as well [20,21,35,46].
3.4. In Vitro Rooting
The survival rates of YV2 microshoots were very high, whereas browning formation was not observed on rooting cultures. The effects of auxins on rooting formation were statistically significant, and the highest rooting percentage (77.78%) was observed on the WPM medium supplemented with 2.0 mg/L of IBA; however, it was statistically different from the 1.0 mg/L of IBA (Table 5).
The highest number of roots was 6.25 per shoot. The longest root length was obtained with 2.0 mg/L of IBA, in which a mean of 1.38 cm was reached, which is also suitable for commercial propagation. The callus formation rate on the deep side of the shoots increased with the increasing concentrations of IBA and NAA (Figure 6).
The inclusion of 4.0 mg/L of IBA had the highest microshoot survival rate in the rooting stage, with a 75.00% rooting rate for YV4. The rooting percentage was the highest with either 4.0 mg/L of NAA or 4.0 mg/L of IBA (66.67% and 55.55%, respectively). Although the root number was satisfactory, the roots were very fragile and surrounded by a smooth, vitrified callus formation. This negated the success of the plantlets’ acclimatization. The roots for this genotype were smaller than 0.5 cm and did not reach commercial size (Table 6). Callus formation with a yellowish-brown color was observed in media with 2 mg/L of IBA and 2 and 4 mg/L of NAA, and callus size increased with higher auxin concentrations. Microshoots continued to elongate in the rooting medium. The maximum shoot elongation was observed in rooting media including 2.0 mg/L of IBA (Figure 6).
The effects of the genotypes were observed in the rooting stage, and when the effective auxin types and concentrations were varied. YV2 microshoots survived when 1.0 and 2.0 mg/L of IBA was included in the medium, whereas these concentrations were not enough to produce roots in the case of the YV4 genotype. Rooting character also differed greatly between the genotypes, with YV2 having superior rooting quality to YV4.
In previous studies, breakable roots with surrounding callus could not be observed on the roots of Arbutus unedo. In our study, YV2 and YV4 had different root characteristics in all culture stages. The rooting rate and response to auxin types changed according to genotype. Gomes et al. [35] found that genotypes could not directly affect rooting percentage, whereas root length was affected by genetic structure.
Auxins alone or in combination with a very low concentration of cytokinin play an important role in root induction. When added in appropriate concentrations, they may regulate cell elongation and the formation of adventitious roots [47]. Exposure time to auxin is another critical point of the rooting stage. Similar results were obtained with other species; Pistacia vera shoots rooted at a high level with external auxin treatment [33]. In this study, we placed the microshoots in rooting media for 45 days. In previous studies, Arbutus unedo shoots were dipped in an IBA solution before ex vitro rooting, or cultured in media containing IBA for a few days to stimulate root formation [36]. On the other hand, Mereti et al. [34] could not obtain an ex vitro rooting rate higher than 60% from Arbutus unedo shoots, whereas the in vitro rooting rate was increased in the WPM medium supplemented with IBA and NAA. They suggested that good-quality roots are the key to the best acclimatization phase. Ex vitro rooting was also successful in some other species [48].
Whereas shoot length was enough, both NAA and IBA increased the shoot elongation in the rooting stage, especially for unrooted microshoots. In future studies, microshoots may be transferred to media supplemented with suitable auxin–cytokinin concentrations. In that case, well-developed shoots in elongation cultures could be transferred to rooting media. In that case, hormone-free media may be sufficient for good rooting if there is sufficient stimulation before the rooting stage.
3.5. Acclimatization
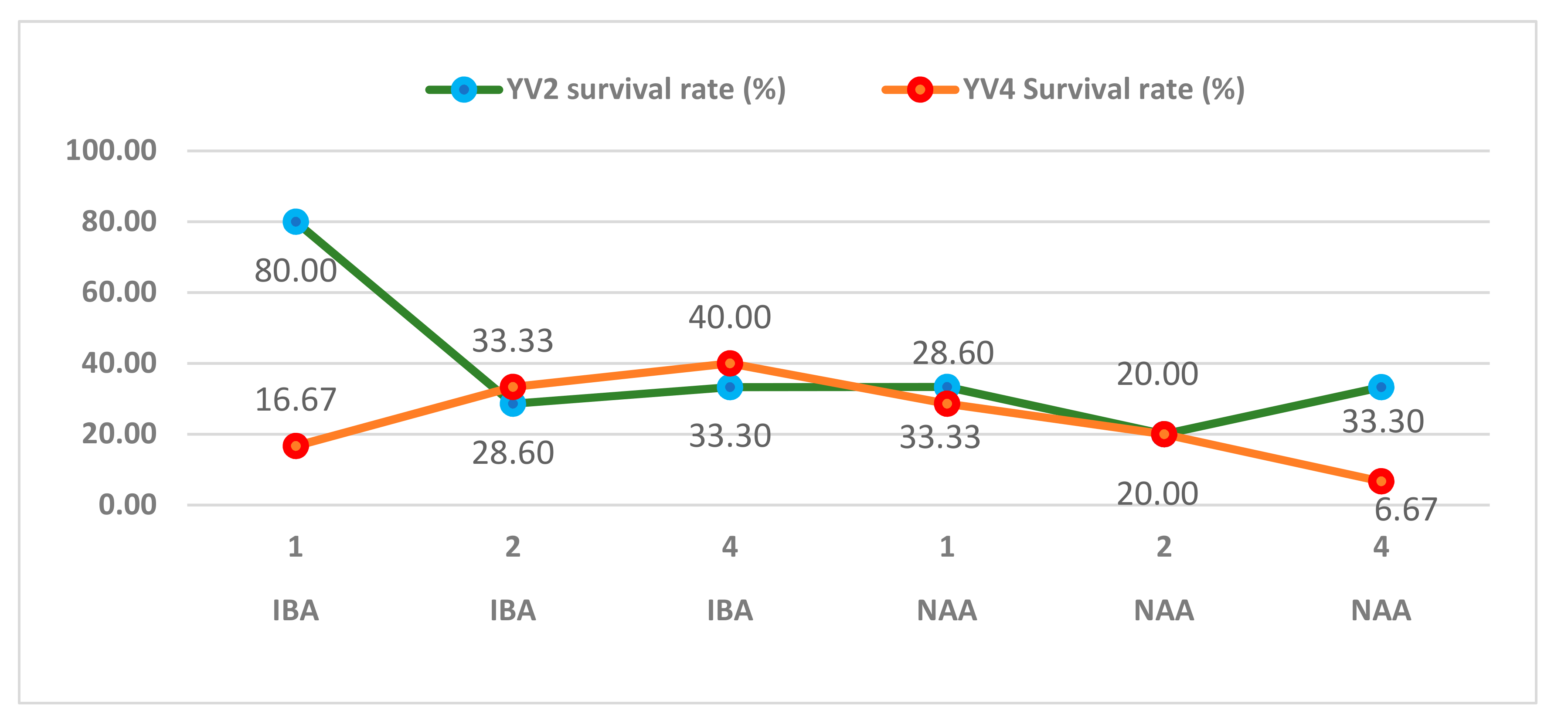
The in vitro plantlets were acclimatized to improve their prospect of survival when transferred to the soil. Approximately 80% of the YV2 plantlets rooted in the medium with 1.0 mg/L of IBA were sufficiently acclimatized after being transferred to ex vitro conditions in the growth room. The other IBA concentrations and all NAA concentrations did not reach the same survival rates (Figure 7). For the YV4 genotype, the highest survival rate was obtained with plantlets rooted in the WPM medium containing 4.0 mg/L of IBA. Plantlets of this genotype rooted in 4.0 mg/L of NAA had vitrified roots with a highly fragile structure; thus, they could not survive the acclimatization stage.
Plantlets were rooted in small, closed tubes at a high air humidity and transferred to the greenhouse in very different environmental conditions. Survival rates are directly dependent on species and acclimatization conditions [45,49]. Results of studies conducted on Arbutus unedo showed that survival of the acclimatization stage is easily achieved for this species if plantlets have a good root quality [34,36]. The failure to rapidly optimize their anatomy and physiology is responsible for their poor performance during acclimatization. In our study, YV4 failed in the step of adapting to external conditions as a result of low root quality, whereasYV2 microshoots successfully acclimatized.
4. Conclusions
Plant tissue culturing is an effective method of commercially producing plantlets. The first step for success is performing optimal explant sterilization in terms of the protocol and selection of appropriate starter materials. We suggest that ethanol with sodium hypochlorite is a satisfactory and economical combination for the sterilization of explants. The observations obtained from the establishment of shoot tip and nodium cultures indicated the suitability of the starting materials for subsequent studies. Despite the differences in the proliferation rates of genotypes in MS and WPM media, we suggest that a suitable medium must be determined for each genotype. Different cultural conditions and mediums should be improved. Again, hormonal content needs to be studied in-depth. In future studies, different cytokine concentrations, together with auxins, may be studied. Although the root quality of the shoots just reached the desired levels in the study, the success in surviving the acclimatization stage reveals that the obstacles concerning the propagation of Arbutus unedo can be overcome via tissue culture. Firstly, it is necessary to improve root quality and supply the strongest roots for in vivo adaptation. İn vivo rooting treatments could be beneficial to increase survival. The degree of success observed in the production of genotypes in previous studies has been quite different, and shows the necessity of continuing studies considering genotypes individually, in order to find a suitable candidate and conditions for commercial applications.
Author Contributions
Conceptualization: M.S.D.; methodology: M.S.D. and S.M.; software: M.S.D.; formal analysis: M.S.D. and S.M.; writing—original draft preparation: M.S.D.; writing—review and editing: M.S.D.; visualization: M.S.D.; supervision and project administration: M.S.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was not funded.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This study was partly produced from the master’s degree thesis of Serdar Memiş at Kocaeli University of Applied Science, Institute of Horticulture, Breeding Department.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schröter, D.; Cramer, W.; Leemans, R.; Prentice, I.C.; Araújo, M.B.; Arnell, N.W.; Bondehau, A.; Bugmann, H.; Carter, T.R.; Gracia, C.A.; et al. Ecosystem service supply and vulnerability to Gobal Change in Europe. Science 2005, 310, 1333–1337. [Google Scholar] [CrossRef] [Green Version]
- Miguel, M.G.; Faleiro, M.L.; Guerreiro, A.C.; Antunes, M.D. Arbutus unedo L.: Chemical and Biological Properties. Molecules 2014, 19, 15799–15823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alarcão-E-Silva, M.; Leitao, A.; Gil Azinheira, H.; Leitão, M. The Arbutus Berry: Studies on its Color and Chemical Characteristics at Two Mature Stages. J. Food Compos. Anal. 2001, 14, 27–35. [Google Scholar] [CrossRef]
- Kalis, A.J.; Schoch, W.H. Anthracological studies on the early Holocene sediments of the Grotta di Corbeddu (Nuoro, Sardinia). Archäologische Ber. 2019, 30, 193–210. [Google Scholar]
- Morgado, S.; Morgado, M.; Plácido, A.I.; Roque, F.; Duarte, A.P. Arbutus unedo L.: From traditional medicine to potential uses in modern pharmacotherapy. J. Ethnopharmacol. 2018, 225, 90–102. [Google Scholar] [CrossRef]
- Torres, J.A.; Valle, F.; Pinto, C.; García-Fuentes, A.; Salazar, C.; Cano, E. Arbutus unedo L. communities in southern Iberian Peninsula mountains. Plant Ecol. 2002, 160, 207–223. [Google Scholar] [CrossRef]
- Takrouni, M.M.; Ali, I.B.E.H.; Messaoued, C.; Boussaid, M. Genetic variability of Tunisian wild strawberry tree (Arbutus unedo L.) populations interfered from isozyme markers. Sci. Hortic. 2012, 146, 92–98. [Google Scholar] [CrossRef]
- Alexandre, A.M.R.C.; Matias, A.A.; Bronze, M.R.; Cocero, M.J.; Mato, R. Phenolic Compounds Extraction of Arbutus unedo L.: Process Intensification by Microwave Pretreatment. Processes 2020, 8, 298. [Google Scholar] [CrossRef] [Green Version]
- Zenginbal, H.; Gündoğdu, M. Physicochemical characterization of strawberry tree (Arbutus unedo L.) genotypes naturally growing in Düzce and Zonguldak province. Anadolu J. Agric. Sci. 2016, 31, 332–336. [Google Scholar] [CrossRef]
- Gündoğdu, M.; Ercisli, S.; Canan, I.; Orman, E.; Sameeullah, M.; Naeem, M.; ben Ayed, R. Diversity in phenolic compounds, biochemical and pomological characteristics of Arbutus unedo fruits. Folia Hortic. 2018, 30, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Bouyahya, A.; Moussaoui, N.; Abrini, I.; Dakka, N. Determination of Phenolic Contents, Antioxidant and Antibacterial Ac-tivities of Strawberry Tree (Arbutus unedo L.) Leaf Extracts. Br. Biotechnol. J. 2016, 14, 1–10. [Google Scholar] [CrossRef]
- Sulusoglu, M.; Cavusoglu, A.; Erkal, S. Arbutus unedo L. (Strawberry tree) selection in Turkey Samanli Mountain Locations. J. Med. Plants Res. 2011, 5, 3545–3551. [Google Scholar]
- Santiso, X.; Lopez, L.; Retuerto, R.; Barreiro, R. Population Structure of a Widespread Species under Balancing Selection: The Case of Arbutus unedo L. Front. Plant Sci. 2016, 6, 1264. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Rodriguez, B.M.; Sachez-Moreno, C.; De Ancos, B.; de Cortes Sánchez-Mata, M.; Fernández-Ruiz, V.; Cámara, M.; Tardío, I. Wild Arbutus unedo L. and Rubus ulmifolius Schott fruits are underutilized sources of valuable bioactive compounds with antioxidant capacity. Fruits 2014, 169, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Montserrat, V.; Einer, J.; Terradas, J. Effect of local competition on resuprouting of Arbutus unedo after Clipping. J. Veg. Sci. 1994, 5, 145–152. [Google Scholar]
- Ogaya, R.; Penuelas, J. Phenological patterns of Quercus ilex, Phillyrea latifolia, and Arbutus unedo growing under a field ex-perimental drought. Ecoscience 2004, 11, 263–270. [Google Scholar] [CrossRef]
- Canadell, J.; López-Soria, L. Lignotuber reserves support regrowth following clipping of two Mediterranean shrubs. Funct. Ecol. 1998, 12, 31–38. [Google Scholar] [CrossRef]
- FAO. The Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture; Food and Agriculture Organization: Rome, Italy, 2010. [Google Scholar]
- Pignatti, G.; Crobeddu, S. Effects of rejuvenation on cutting propagation of Mediterranean shrub species. Forest 2005, 2, 290–295. [Google Scholar] [CrossRef]
- Sulusoglu, M. Development of a Rooted Cutting Propagation Method for Selected Arbutus unedo L. Types and Seasonal Variation in Rooting Capacity. J. Agric. Sci. 2012, 4, 216. [Google Scholar] [CrossRef]
- Metaxas, D.J.; Syros, T.D.; Yupsanis, T.; Economou, A.S. Peroxidases during adventitious rooting in cuttings of Arbutus unedo and Taxus baccata as affected by plant genotype and growth regulator treatment. Plant Growth Regul. 2004, 44, 257–266. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and biossays with tobacco cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lloyd, G.B.; McCown, B.H. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shhot-tip culture. Proc. Int. Plant Prop. Soc. 1981, 30, 421–427. [Google Scholar]
- Vijayan, K.; Tikader, A.; da Silva, J.A.T. Application of Tissue Culture Techniques for Propagation and Crop. Improvement in Mulberry (Morus spp.). Tree For. Sci. Biotechnol. 2011, 5, 1–13. [Google Scholar]
- Wolella, E.K. Surface sterilization and in vitro propagation of Prunus domestica L. cv. Stanley using axillary buds as explants. J. Biotech. Res. 2017, 8, 18–26. [Google Scholar]
- Örgeç, M.; Karakaş, F.P.; Sahin, G.; Ağıl, F.; Zencirci, N. Einkorn (Triticum monococcum ssp. monococcum) in vitro propagation sterilization protocol. Int. J. Second. Metab. 2018, 5, 67–74. [Google Scholar] [CrossRef]
- Taha, H.; Ghazy, U.M.; Magdy Gabr, A.M.; EL-Kazzaz, A.A.A.; Ahmed, E.A.M.M.; Haggag, K.M. Optimization of in vitro culture conditions affecting propagation of mulberry plant. Bull. Natl. Res. Cent. 2020, 44, 60. [Google Scholar] [CrossRef] [Green Version]
- Damiano, C.; Padro, M.D.A.; Frattarelli, A. Propagation and establishment In vitro of myrtle (Myrtus communis L.), pomegranate (Punica granatum L.) and mulberry (Morus Alba L.). Propag. Ornam. Plants 2008, 8, 3–8. [Google Scholar]
- Elmaghabi, A.M.; Hammud, S.; Abugnia, E. In Vitro Plant Regeneration of Libyan Wild Plants: Edible Species (Arbutus pavar-ii) and Endanger Species [Haplophyllum tuberculatum (Forsk.) Juss]. ANADOLU J. Aegean Agric. Res. Inst. 2017, 27, 127–132. [Google Scholar]
- Banu, N.A.; İslam, S.; İslam, M.A.; Alam, M.K. In vitro Propagation from Shoot Tip and Nodal Segment in Summer Tomato (Lycopersicon esculentum Mill.). Plant Environ. Dev. 2017, 6, 31–38. [Google Scholar]
- Bertsouklis, K.; Papafotiou, M. In vitro propagation of Arbutus andrachne L. Acta Hortic. 2009, 813, 477–480. [Google Scholar] [CrossRef]
- Boudabous, M.; Mars, M.; Marzouguni, N.; Ferchichi, A. Micropropagation of apple (Malus communis L. Cultivar Douce de Djerba) through in vitro culture of axillary buds. Acta Bot. Gall. 2010, 157, 513–524. [Google Scholar] [CrossRef]
- Benmahioul, B.; Kaïd-Harche, M.; Daguin, F. In vitro regeneration of Pistacia vera L. from nodal explants. J. For. Sci. 2016, 62, 198–203. [Google Scholar] [CrossRef]
- Mereti, M.; Grigoriadou, K.; Nanos, G.D. Micropropagation of the strawberry tree, Arbutus unedo L. Sci. Hortic. 2002, 93, 143–148. [Google Scholar] [CrossRef]
- Gomes, F.; Simões, M.; Lopes, M.L.; Canhoto, J.M. Effect of plant growth regulators and genotype on the micropropagation of adult trees of Arbutus unedo L. (strawberry tree). New Biotechnol. 2010, 27, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.; Canhoto, J.M. Micropropagation of strawberry tree (Arbutus unedo L.) from adult plants. Vitr. Cell. Dev. Biol. Plant 2009, 45, 72–82. [Google Scholar] [CrossRef]
- Youmbi, E.; Nyambeu, M.N.T.; Ngaha, D.; Fonbah, C. In vitro proliferation ability of axillary buds in Musa spp. Agron. Afr. 2014, 26, 1–7. [Google Scholar]
- Roussos, P.; Pontikis, C.A. Phenolic compounds in olive explants and their contribution to browning during the establishment stage in vitro. Gartenbauwissenschaft 2001, 66, 298–303. [Google Scholar]
- Ko, H.; Su, C.C.; Chen, C.L.; Chao, C.P. Control of lethal browning of tissue culture plantlets of Cavendishbanana cv. For-mosana with ascorbic acid. Plant Cell Tiss Organ Cult. 2009, 96, 137–141. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Fang, S.; Fu, X.; Shang, X.; Yang, W. Seasonal Variation in Phenolic Compounds and Antioxidant Activity in Leaves of Cyclocarya paliurus (Batal.) Iljinskaja. Forests 2019, 10, 624. [Google Scholar] [CrossRef] [Green Version]
- Dal Vesco, L.; Guerra, M.P. The effectiveness of nitrogen sources in Feijoa somatic embryogenesis Lirio. Plant Cell. Tissue Organ Cult. 2001, 64, 19–25. [Google Scholar] [CrossRef]
- Kaplan, N.; İslam, A.; Bilir Ekbiç, H. Çakıldak Fındık çeşidinin in vitro sürgün ucu kültür ile çoğaltılması. Akad. Ziraat Derg. 2020, 9, 193–200. [Google Scholar]
- Karam, N.S.; Al-Salem, M.M. Breaking dormancy in Arbutus andrachna L. seeds by stratification and gibberellic acid. Seed Sci. Technol. 2001, 29, 51–56. [Google Scholar]
- Gajdosova, A.A.; Libiakova, G.; Fejer, J. Improvement of selected Amaranthus cultivars by means of mutation induction and biotechnological approaches. In Breeding of Neglected and Under-Utilized Crops, Spices and Herbs; Science Publishers Inc.: Hauppauge, NY, USA, 2007; pp. 151–169. [Google Scholar]
- Mostafa, S.E.; Karam, N.S.; Shibli, R.A.; Alali, F.Q. Micropropagation and production of arbutin in oriental strawberry tree (Arbutus andrachne L.). Plant Cell Tissue Organ Cult. (PCTOC) 2010, 103, 111–121. [Google Scholar] [CrossRef]
- Sulusoglu, M.; Memis, S. In vitro Culture Establishment and Plant Regeneration of Arbutus unedo L. In Proceedings of the XXX International Horticultural Congress IHC2018, İstanbul, Turkey, 12–16 August 2018; Acta Horticulturae: Leuven, Belgium, 2020; Volume 1285, pp. 109–112. [Google Scholar]
- Su, Y.H.; Liu, Y.B.; Zhang, X.S. Auxin–Cytokinin Interaction Regulates Meristem Development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yeh, D. Elimination of In vitro Contamination, Shoot Multiplication, and Ex vitro Rooting of Aglaonema. HortScience 2007, 42, 629–632. [Google Scholar] [CrossRef] [Green Version]
- Shibli, R.A.; Jaradat, A.; Ajlouni, M.; Aljanabi, S.; Shatnawi, M.A. Micropropagation in wild pear (Pyrus syrica). Sci. Hortic. 1997, 68, 237–242. [Google Scholar] [CrossRef]
Figure 1.
Naturally growing plants of Arbutus unedo L. genotypes (YV2 and YV4) in the forest land.

Figure 2.
Epicormic shoot formation from the woody shoots of Arbutus unedo.

Figure 3.
Axillary shoot formation from nodium and shoot tip sprouting.

Figure 4.
Proliferation stage of YV2 genotype in WPM and MS media supplemented with different growth regulators as indicated.
Figure 4.
Proliferation stage of YV2 genotype in WPM and MS media supplemented with different growth regulators as indicated.

Figure 5.
Proliferation stage of YV4 genotype in the WPM and MS media supplemented with different growth regulators.
Figure 5.
Proliferation stage of YV4 genotype in the WPM and MS media supplemented with different growth regulators.

Figure 6.
Root formation in YV2 and YV4 microshoots.

Figure 7.
Survival rate of in vitro propagated YV2 and YV4 genotype plantlets at 4 weeks after transplanting.
Figure 7.
Survival rate of in vitro propagated YV2 and YV4 genotype plantlets at 4 weeks after transplanting.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of sterilization treatments with ethanol and sodium hypochlorite on the survival, infection, or browning of explants.
Table 1.
Effect of sterilization treatments with ethanol and sodium hypochlorite on the survival, infection, or browning of explants.
| Treatment | Survival (%) | Infection (%) | Browning (%) | ||||
|---|---|---|---|---|---|---|---|
| Shoot Tip | Nodium | Shoot Tip | Nodium | Shoot Tip | Nodium | ||
| 1. | 70% EtOH 2 min + 20% NaCIO 15 min | 44.81 de * | 30.56 ef | 8.52 d | 31.02 ab | 46.67 a | 38.43 ab |
| 2. | 70% EtOH 1 min + 20% NaCIO 12 min | 64.65 bc | 77.38 ab | 23.74 bc | 8.9 d | 11.62 de | 13.69 de |
| 3. | 50% EtOH 1 min + 20% NaCIO 10 min | 55.28 cd | 65.51 bc | 13.37 cd | 12.24 cd | 31.35 bc | 22.25 cd |
| 4. | 50% EtOH 1 min + 20% NaCIO 12 min | 85.06 a | 84.41 a | 7.14 d | 7.52 d | 7.79 e | 8.07 e |
| 5. | 20% NaCIO 15 min | 58.12 cd | 26.50 f | 29.96 ab | 39.6 a | 11.92 de | 33.91 abc |
* Shoot tips and nodiums were evaluated separately for statistical analysis; * Parameter values in the same column followed by different letters indicate the values are significantly different according to the Tukey test (p < 0.05).
Table 2.
Survival, proliferation, and shoot development in the establishment stage of Arbutus genotypes.
Table 2.
Survival, proliferation, and shoot development in the establishment stage of Arbutus genotypes.
| Survival (%) | Browning (%) | Proliferation (%) | Shoot Number | Shoot Length (cm) | ||
|---|---|---|---|---|---|---|
| YV2 * | Shoot tip | 93.33 a * | 6.67 b | 0.00 b | 0.00 b | 0.00 b |
| Nodium | 86.67 a | 13.33 a | 35.00 a | 1.67 a | 0.53 a | |
| YV4 * | Shoot tip | 73.33 a | 26.67 a | 0.00 b | 0.00 b | 0.00 b |
| Nodium | 73.33 a | 26.67 a | 27.78 a | 1.00 a | 0.63 a | |
* Each genotype was evaluated individually; any two mean values in the same column followed by a different letter are significantly different according to the Tukey test, (p < 0.05).
Table 3.
Effect of growth hormones on explant response and survival, as determined through the percentages of proliferation and browning, as well as the shoot number and shoot length of YV2.
Table 3.
Effect of growth hormones on explant response and survival, as determined through the percentages of proliferation and browning, as well as the shoot number and shoot length of YV2.
| Nutrient Medium and Growth Regulators | Proliferation (%) | Shoot Number | Shoot Length (cm) | Browning (%) | ||||
|---|---|---|---|---|---|---|---|---|
| BAP (mg/L) | GA3 (mg/L) | IBA (mg/L) | Medium | |||||
| 1 | 0.5 | 0.1 | - | MS | 72.00 abcd * | 1.60 abcd | 1.47 abcd | 5.00 c |
| 2 | 1.0 | 0.1 | - | MS | 80.00 abc | 1.66 abc | 1.64 abc | 5.00 c |
| 3 | 2.0 | 0.1 | - | MS | 96.00 ab | 1.82 ab | 1.76 ab | 0.00 c |
| 4 | 4.0 | 0.1 | - | MS | 80.00 abc | 1.74 abc | 1.18 bcde | 5.00 c |
| 5 | 1.0 | 0.25 | 0.5 | MS | 12.94 e | 0.84 d | 0.60 e | 45.34 a |
| 6 | 2.0 | 0.25 | 0.5 | MS | 57.94 cd | 1.36 bcd | 1.09 bcde | 64.07 a |
| 7 | 4.0 | 0.25 | 0.5 | MS | 42.33 cde | 1.18 bcd | 0.73 de | 46.61 a |
| 8 | 6.0 | 0.25 | 0.5 | MS | 31.46 de | 0.85 d | 0.72 de | 40.00 ab |
| 9 | 1.0 | 0.1 | 0.1 | WPM | 73.20 abcd | 2.32 a | 1.58 abc | 8.32 bc |
| 10 | 2.0 | 0.1 | 0.1 | WPM | 100 a | 1.72 abc | 2.03 a | 28.00 abc |
| 11 | 4.0 | 0.1 | 0.1 | WPM | 66.66 cd | 1.12 bcd | 1.27 abcde | 30.07 abc |
* Parameter values in the same column, followed by different letters, are significantly different according to the Tukey test (p < 0.05).
Table 4.
Effect of growth hormones on the percentages of explants responding, surviving, browning, and proliferating, as well as their effect on the shoot number and shoot length of YV4.
Table 4.
Effect of growth hormones on the percentages of explants responding, surviving, browning, and proliferating, as well as their effect on the shoot number and shoot length of YV4.
| Nutrient Medium and Growth Regulators | Proliferation (%) | Shoot Number | Shoot Length (cm) | Browning (%) | ||||
|---|---|---|---|---|---|---|---|---|
| BAP (mg/L) | GA3 (mg/L) | IBA (mg/L) | Medium | |||||
| 1 | 0.5 | 0.1 | - | MS | 42.67 bcd * | 1.12 cd | 0.89 bc | 4.00 c |
| 2 | 1.0 | 0.1 | - | MS | 32.00 d | 1.06 cd | 0.75 c | 0.00 c |
| 3 | 2.0 | 0.1 | - | MS | 52.00 abcd | 1.66 bcd | 1.15 bc | 0.00 c |
| 4 | 4.0 | 0.1 | - | MS | 40.00 bcd | 2.20 abcd | 1.42 bc | 0.00 c |
| 5 | 1.0 | 0.25 | 0.5 | MS | 18.67 d | 1.20 cd | 0.57 c | 50.64 ab |
| 6 | 2.0 | 0.25 | 0.5 | MS | 59.33 abcd | 2.24 abc | 1.08 bc | 20.67 abc |
| 7 | 4.0 | 0.25 | 0.5 | MS | 50.11 abcd | 1.66 bcd | 0.77 c | 50.67 ab |
| 8 | 6.0 | 0.25 | 0.5 | MS | 29.33 d | 1.18 cd | 0.53 c | 56.00 a |
| 9 | 1.0 | 0.1 | 0.1 | WPM | 79.33 ab | 2.82 ab | 1.76 ab | 12.31 bc |
| 10 | 2.0 | 0.1 | 0.1 | WPM | 86.67 a | 3.22 a | 2.63 a | 20.00 abc |
| 11 | 4.0 | 0.1 | 0.1 | WPM | 76.00 abc | 2.10 abcd | 1.45 bc | 21.90 abc |
* Parameter values in the same column, followed by different letters, are significantly different according to the Tukey test (p < 0.05).
Table 5.
The effects of auxins (IBA and NAA) on the rooting of in vitro-raised microshoots of YV2 on the WPM medium.
Table 5.
The effects of auxins (IBA and NAA) on the rooting of in vitro-raised microshoots of YV2 on the WPM medium.
| Concentration (mg/L) | Survival * (%) | Rooting (%) | Root Number | Root Length (cm) | Callus (%) | Callus Size (cm) | Shoot Length (cm) |
|---|---|---|---|---|---|---|---|
| 1 IBA | 100 | 22.22 b * | 3.00 b | 0.86 b | 13.33 b | 0.51 | 2.12 bc |
| 2 IBA | 100 | 77.78 a | 6.25 a | 1.38 a | 33.37 ab | 0.50 | 2.55 a |
| 4 IBA | 100 | 55.55 ab | 6.00 a | 0.89 b | 55.55 a | 0.50 | 1.93 d |
| 1 NAA | 88.88 | 66.66 ab | 3.75 b | 0.58 c | 11.11 b | 0.60 | 2.05 c |
| 2 NAA | 88.88 | 55.55 ab | 3.00 b | 0.40 d | 37.33 ab | 0.57 | 2.18 b |
| 4 NAA | 100 | 44.44 ab | 5.50 a | 0.38 d | 55.5 a | 0.50 | 1.82 e |
* Parameter values in the same column followed by different letters are significantly different according to the Tukey test (p < 0.05).
Table 6.
The effects of auxins (IBA and NAA) on the rooting of in vitro-cultured microshoots of YV4 on WPM medium.
Table 6.
The effects of auxins (IBA and NAA) on the rooting of in vitro-cultured microshoots of YV4 on WPM medium.
| Concentration (mg/L) | Survival * (%) | Rooting * (%) | Root Number | Root Length (cm) | Callus (%) | Callus Size (cm) | Shoot Length (cm) |
|---|---|---|---|---|---|---|---|
| 1 IBA | 75.00 | 0.00 c * | 0.00 b | 0.00 b | 0.00 b | 0.00 b | 2.22 bc |
| 2 IBA | 100 | 0.00 c | 0.00 b | 0.00 b | 0.00 b | 0.00 b | 2.64 a |
| 4 IBA | 75.00 | 55.55 a | 4.00 a | 0.32 a | 27.33 a | 0.90 a | 2.46 ab |
| 1 NAA | 33.33 | 33.33 b | 6.00 a | 0.45 a | 0.00 b | 0.00 b | 2.37 ab |
| 2 NAA | 66.67 | 0.00 c | 0.00 b | 0.00 b | 13.33 ab | 0.57 ab | 1.95 c |
| 4 NAA | 66.67 | 66.66 a | 4.00 a | 0.40 a | 17.33 ab | 0.81 a | 1.60 d |
* Parameter values in the same column followed by different letters are significantly different according to the Tukey test (p < 0.05).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sulusoglu Durul, M.; Memis, S. Optimization of Conditions for In Vitro Culture of Selected Arbutus unedo L. Genotypes. Agronomy 2022, 12, 623. https://doi.org/10.3390/agronomy12030623
AMA Style
Sulusoglu Durul M, Memis S. Optimization of Conditions for In Vitro Culture of Selected Arbutus unedo L. Genotypes. Agronomy. 2022; 12(3):623. https://doi.org/10.3390/agronomy12030623
Chicago/Turabian StyleSulusoglu Durul, Melekber, and Serdar Memis. 2022. "Optimization of Conditions for In Vitro Culture of Selected Arbutus unedo L. Genotypes" Agronomy 12, no. 3: 623. https://doi.org/10.3390/agronomy12030623
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.