
Agro-Morphological and Molecular Variability among Algerian Faba Bean (Vicia faba L.) Accessions


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
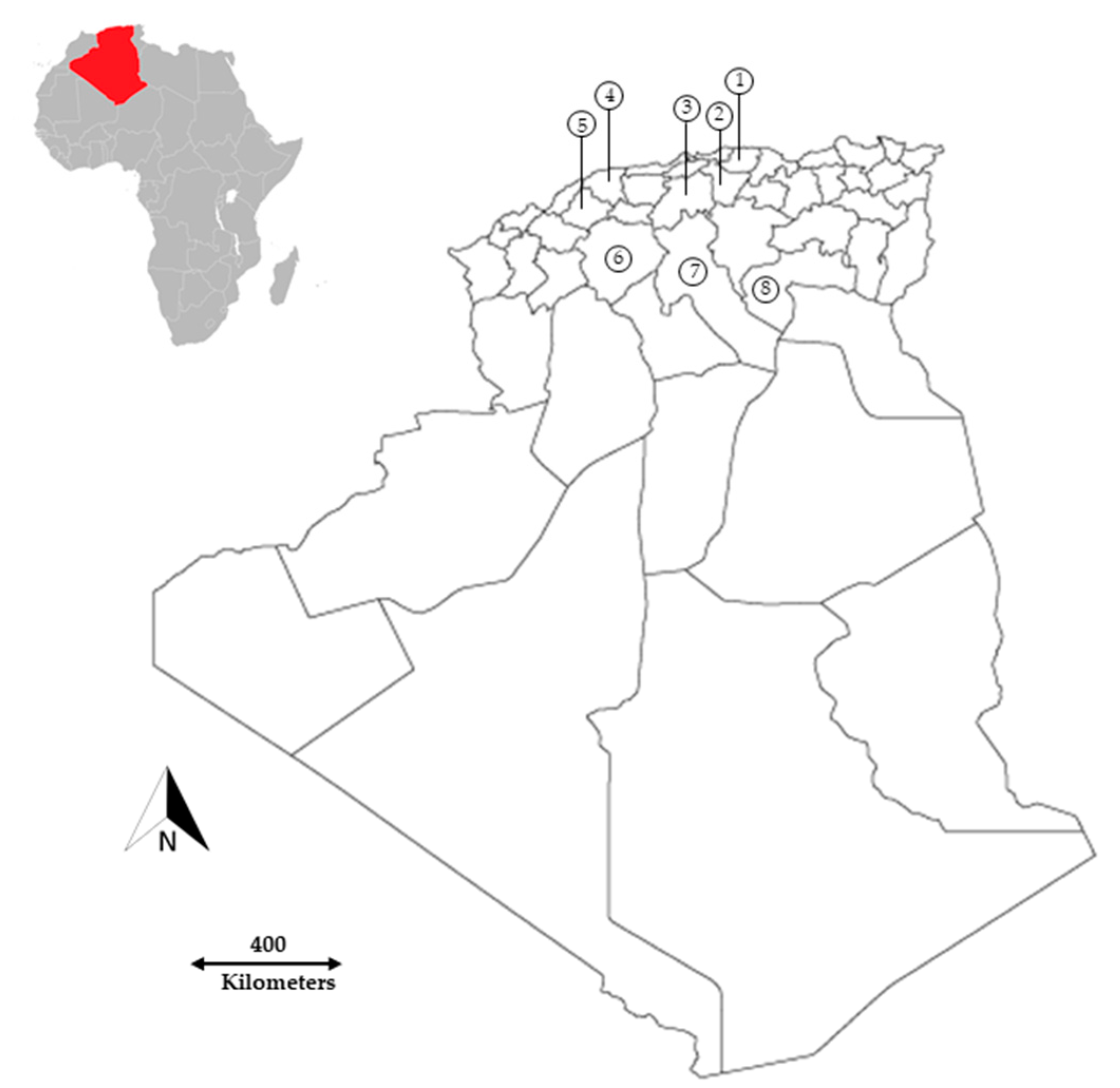
2.1. Plant Material
2.2. Agro-Morphological Analysis
Experimental Design and Agro-Morphological Traits
2.3. Molecular Analysis
2.3.1. DNA Extraction, Quantification, and Quality Analysis
2.3.2. PCR Amplification and SSR Analysis
2.4. Statistical Analysis
2.4.1. Agro-Morphological Data
2.4.2. Molecular Data
3. Results
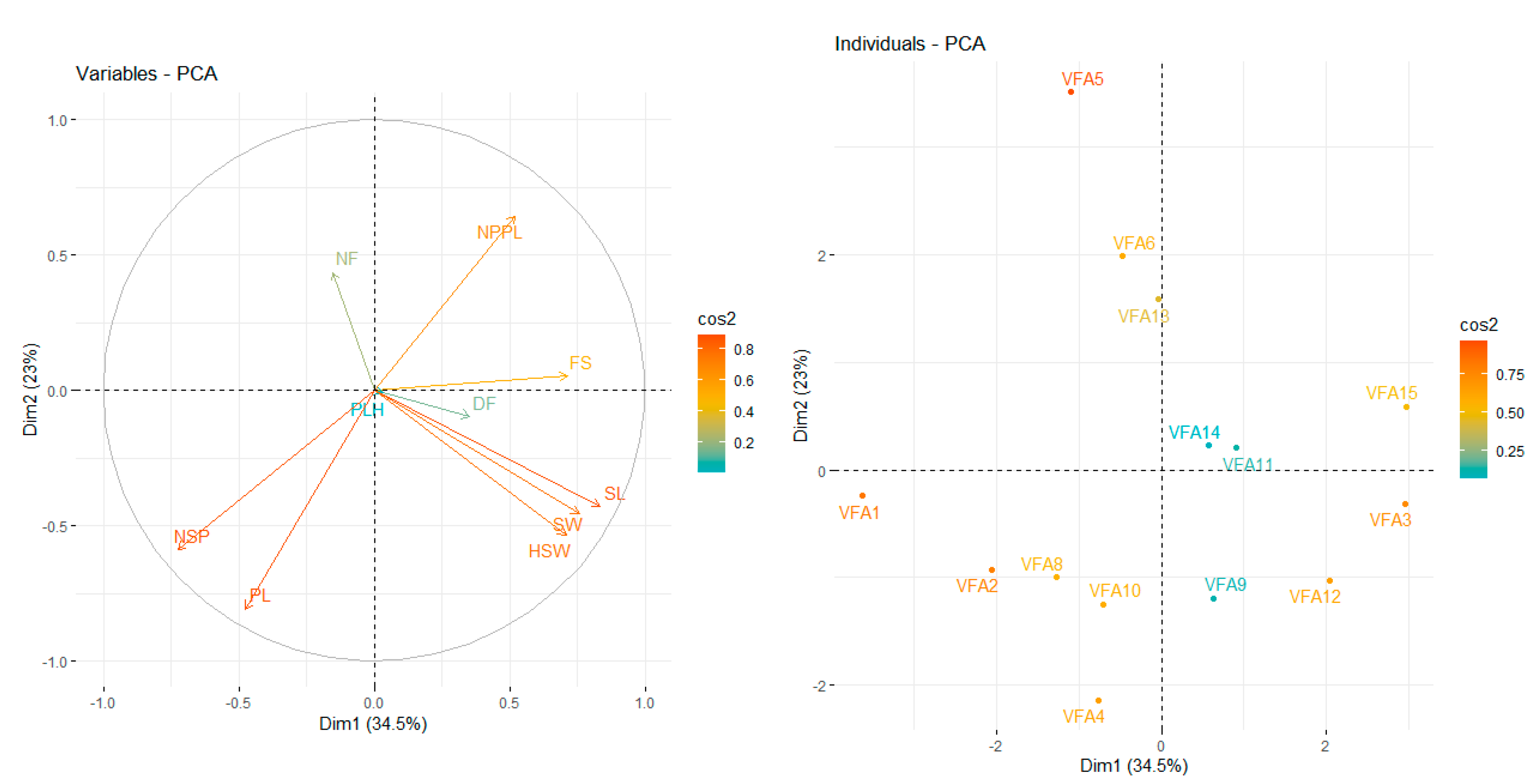
3.1. Agro-Morphological Traits
3.2. Molecular Characterization
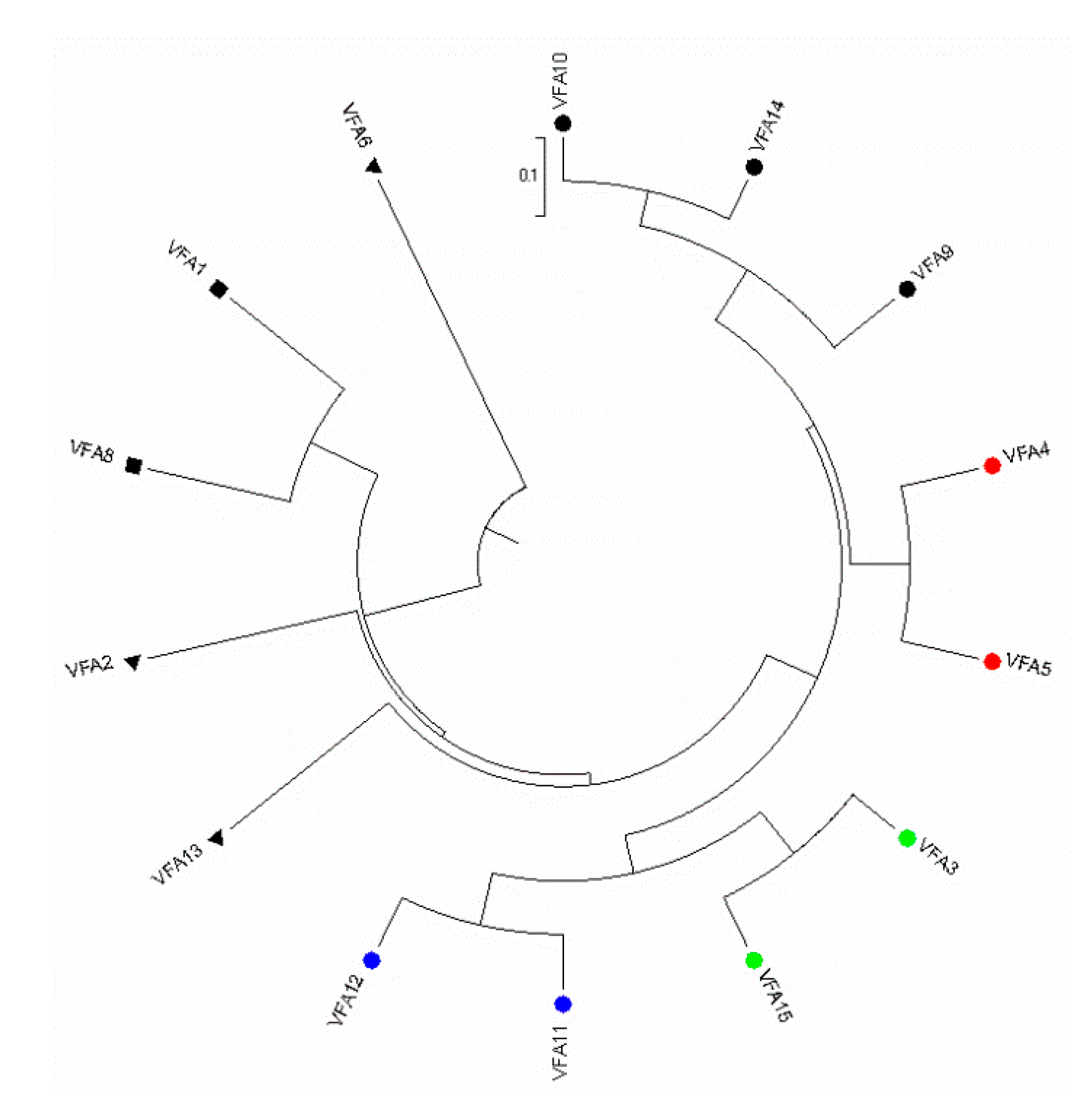
3.3. Genetic Structure
4. Discussion
4.1. Agromorphological Characterization
4.2. Molecular Characterization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tanno, K.; Willcox, G. The origins of Cicer arietinum L. and Vicia faba L.: Early finds from Tell el Kerkh. north-west Syria. late 10th millennium B.P. Veg. Hist. Archaeobot. 2006, 15, 197–204. [Google Scholar] [CrossRef]
- O’Sullivan, D.M.; Angra, D. Advances in faba bean genetics and genomics. Front. Genet. 2016, 7, 150. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization Corporate Statistical Database. Available online: http://www.fao.org/food-agriculture-statistics/en/ (accessed on 2 June 2021).
- FAOSTAT. Production Statistics of the Food Agriculture Organization of The United States. Available online: http://www.fao.org/faostat/en/#data/QA (accessed on 3 June 2021).
- Hamadache, A. Grandes cultures: Principaux itinéraires techniques des principales espèces de grandes cultures pluviales cultivées en Algérie et en Afrique du Nord (agriculture conventionnelle) Légumineuses alimentaires (Pois chiche-Féves-Lentille); Tome II; Ecole Nationale Supérieure Agronomique: Alger, Algerie, 2014; pp. 1–187. [Google Scholar]
- Duc, G.; Bao, S.; Baum, M.; Redden, B.; Sadiki, M.; Suso, M.J.; Vishniakova, M.; Zong, X. Diversity maintenance and use of Vicia faba L. genetic resources. Field Crop. Res. 2010, 115, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Terzopoulos, P.; Kaltsikes, P.; Bebeli, P. Collection, evaluation and classification of Greek populations of faba bean (Vicia faba L.). Genet. Resour. Crop. Evol. 2003, 50, 373–381. [Google Scholar] [CrossRef]
- Vörösváry, G.; Holly, L.; Strajeru, S.; Tamás, J.; Constantinovici, D.; Málnási Csizmadia, G.; Horváth, I. Studies on the varia-tion of agronomic traits in some faba bean (Vicia faba L.) landraces from Romania. Bulletin of University of Agricultural Sciences and Veterinary Medicine. Horticulture 2011, 68, 279–283. [Google Scholar]
- Ouji, A.; Rouaissi, M.; Abdellaoui, R.; El Gazzah, M. The use of reproductive vigor descriptors in studying genetic variability in nine Tunisian faba bean (Vicia faba L.) populations. Afr. J. Biotechnol. 2011, 10, 896–904. [Google Scholar]
- Yahia, Y.; Guetat, A.; Elfalleh, W.; Ferchichi, A.; Yahia, H.; Loumerm, M. Analysis of agromorphological diversity of southern Tunisia faba bean (Vicia faba L.) germplasm. Afr. J. Biotechnol. 2012, 11, 11913–11924. [Google Scholar]
- Ammar, M.H.; Anwar, F.; El-Harty, E.H.; Migdadi, H.M.; Abdel-Khalik, S.M.; A Alfaifi, S.; Farooq, M.; Alghamdi, S.S. Physiological and Yield Responses of Faba bean (Vicia faba L.) to Drought Stress in Managed and Open Field Environments. J. Agron. Crop. Sci. 2014, 201, 280–287. [Google Scholar] [CrossRef]
- Zong, X.; Liu, X.; Guan, J.; Wang, S.; Liu, Q.; Paull, J.G.; Redden, R. Molecular variation among Chinese and global winter faba bean germplasm. Theor. Appl. Genet. 2009, 118, 971–978. [Google Scholar] [CrossRef]
- Torres, A.M.; Weeden, N.F.; Martín, A. Linkage among isozyme, RFLP and RAPD markers in Vicia faba. Theor. Appl. Genet. 1993, 85, 937–945. [Google Scholar] [CrossRef]
- Link, W.; Dixkens, C.; Singh, M.; Schwall, M. Genetic diversity in European and Mediterranean faba bean germplasm revealed by RAPD markers. Theor. Appl. Genet. 1995, 90, 27–32. [Google Scholar] [CrossRef]
- Tahir, N.A. Identification of Genetic Variation in Some Faba Bean (Vicia faba L.) Genotypes Grown in Iraq Estimated with RAPD and SDS-PAGE of Seed Proteins. Indian J. Biotechnol. 2015, 14, 351–356. [Google Scholar]
- Kaur, S.; Kimber, R.B.; Cogan, N.O.; Materne, M.; Forster, J.W.; Paull, J.G. SNP discovery and high-density genetic mapping in faba bean (Vicia faba L.) permits identification of QTLs for ascochyta blight resistance. Plant Sci. 2014, 217-218, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Mulugeta, B.; Tesfaye, K.; Keneni, G.; Ahmed, S. Genetic diversity in spring faba bean (Vicia faba L.) genotypes as revealed by high-throughput KASP SNP markers. Genet. Resour. Crop. Evol. 2021, 68, 1971–1986. [Google Scholar] [CrossRef]
- Alghamdi, S.S.; Al-Faifi, S.A.; Migdadi, H.M.; Ammar, M.H.; Siddique, K. Inter-simple sequence repeat (ISSR)-based diversity assessment among faba bean genotypes. Crop. Pasture Sci. 2011, 62, 755–760. [Google Scholar] [CrossRef]
- Wang, H.-F.; Zong, X.-X.; Guan, J.-P.; Yang, T.; Sun, X.-L.; Ma, Y.; Redden, R. Genetic diversity and relationship of global faba bean (Vicia faba L.) germplasm revealed by ISSR markers. Theor. Appl. Genet. 2011, 124, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Bao, S.-Y.; Ford, R.; Jia, T.-J.; Guan, J.-P.; He, Y.-H.; Sun, X.-L.; Jiang, J.-Y.; Hao, J.-J.; Zhang, X.-Y.; et al. High-throughput novel microsatellite marker of faba bean via next generation sequencing. BMC Genom. 2012, 13, 602. [Google Scholar] [CrossRef] [Green Version]
- Yahia, Y.; Hannachi, H.; Monforte, A.J.; Cockram, J.; Loumerem, M.; Zarouri, B.; Ferchichi, A. Genetic diversity in Vicia faba L. populations cultivated in Tunisia revealed by simple sequence repeat analysis. Plant Genet. Resour. 2014, 12, 278–285. [Google Scholar] [CrossRef]
- El-Esawi, M.A. SSR analysis of genetic diversity and structure of the germplasm of faba bean (Vicia faba L.). Comptes Rendus Biol. 2017, 340, 474–480. [Google Scholar] [CrossRef]
- Babay, E.; Khamassi, K.H.; Sabetta, W.; Miazzi, M.M.; Montemurro, C.; Pignone, D.; Danzi, D.; Finetti-Sialer, M.M.; Mangini, G. Serendipitous in Situ Conservation of Faba Bean Landraces in Tunisia: A Case Study. Genes 2020, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Alghamdi, S.S.; Al-Faifi, S.A.; Migdadi, H.M.; Khan, M.A.; El-Harty, E.H.; Ammar, M.H. Molecular Diversity Assessment Using Sequence Related Amplified Polymorphism (SRAP) Markers in Vicia faba L. Int. J. Mol. Sci. 2012, 13, 16457–16471. [Google Scholar] [CrossRef]
- International Board for Plant Genetic Resources. Faba Bean Descriptors; IBPGR: Rome, Italy, 1985; p. 31. [Google Scholar]
- Union for The Protection of New Varieties of Plants. Broad Bean (Vicia Faba var Major Harz); UPOV: Bern, Switzerland, 2003; p. 29. [Google Scholar]
- Fulton, T.M.; Chunwongse, J.; Tanksley, S.D. Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Rep. 1995, 13, 207–209. [Google Scholar] [CrossRef]
- Zeid, M.; Mitchell, S.; Link, W.; Carter, M.; Nawar, A.; Fulton, T.; Kresovich, S. Simple sequence repeats (SSRs) in faba bean: New loci fromOrobanche-resistant cultivar ‘Giza 402’. Plant Breed. 2009, 128, 149–155. [Google Scholar] [CrossRef]
- Požárková, D.; Koblížková, A.; Román, B.; Torres, A.; Lucretti, S.; Lysák, M.; Dolezel, J.; Macas, J. Development and Characterization of Microsatellite Markers from Chromosome 1-Specific DNA Libraries of Vicia Faba. Biol. Plant. 2002, 45, 337–345. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 5 March 2021).
- Fernández, A.; Gómez, S. Solving Non-Uniqueness in Agglomerative Hierarchical Clustering Using Multidendrograms. J. Classif. 2008, 25, 43–65. [Google Scholar] [CrossRef] [Green Version]
- Aci, M.M.; Lupini, A.; Badagliacca, G.; Mauceri, A.; Lo Presti, E.; Preiti, G. Genetic diversity among Lathyrus spp. based on agronomic and molecular markers. Agronomy 2020, 10, 1182. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. genalex 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Excoffier, L.L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplo-types: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Battistuzzi, F.U. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Li., W. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy—The Principles and Practice of Numerical Classification; W.H. Freeman: San Francisco, CA, USA, 1973. [Google Scholar]
- Maalouf, F.; Hu, J.; O’Sullivan, D.M.; Zong, X.; Hamwieh, A.; Kumar, S.; Baum, M. Breeding and genomics status in faba bean (Vicia faba). Plant Breed. 2018, 138, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Qahtan, A.A.; Al-Atar, A.; Abdel-Salam, E.M.; El-Sheikh, M.A.; Gaafar, A.-R.Z.; Faisal, M. Genetic diversity and structure analysis of a worldwide collection of faba bean (Vicia faba) genotypes using ISSR markers. Int. J. Agric. Biol. 2021, 25, 683–691. [Google Scholar]
- Ouji, A.; El-Bok, S.; Rouaissi, M.; Syed, N.H.; Suso, M.J.; Flavell., A.J.; El Gazzah., M.; Ben Younes., M.; Kharrat, M. Genetic diversity of faba bean (Vicia faba L.) Populations estimated by isozymic and molecular markers: Relationship between the two methods. Eur. Sci. J. 2015, 11, 392–401. [Google Scholar]
- Sadiki, M.; Amri, A.; Birouk, A.; Bounejmate, M. Les ressources phytogénétiques des plantes cultivées au Maroc. In Etude de Faisabilité du Projet FEM/PNUD/FAO sur la Conservation et la Valorisation des Ressources Génétiques des Plantes au Maghreb; Ministère de l’Environnement: Rabat, Maroc, 1995. [Google Scholar]
- Terzopoulos, P.; Bebeli, P. Genetic diversity analysis of Mediterranean faba bean (Vicia faba L.) with ISSR markers. Field Crop. Res. 2008, 108, 39–44. [Google Scholar] [CrossRef]
- Perrino, P.; Robertson, L.D.; Silh, M.B. Maintenance. evaluation and use of faba bean germplasm collections: Problems and prospects. CIHEAM Options Méditerranéenne 1991, 110, 21–31. [Google Scholar]
- Polignano, G.B.; Alba, E.; Uggenti, P.; Scippa, G. Geographical Patterns of Variation in Bari Faba Bean Germplasm Collection. Genet. Resour. Crop. Evol. 1999, 46, 183–192. [Google Scholar] [CrossRef]
- Keneni, G.; Jarso, M.; Wolabu, T.; Dino, G. Extent and Pattern of Genetic Diversity for Morpho-agronomic Traits in Ethiopian Highland Pulse Landraces II. Faba Bean (Vicia faba L.). Genet. Resour. Crop. Evol. 2005, 52, 551–561. [Google Scholar] [CrossRef]
- Labuda, H. Flowering and pod setting of faba bean (Vicia faba L. var. major Harz) depending on nitrogen fertilization. Ann. UMCS Sect. EEE 2002, 10, 235–240. [Google Scholar]
- Singh, R.J.; Jauhar, P.P. Genetic Resources, Chromosome Engineering and Crop Improvement, 1st ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 204–228. [Google Scholar]
- Łabuda, H. Flowering and characteristics of useful traits of some faba bean (Vicia faba L. var. major Harz) cultivars and breeding lines. Acta Agrobot. 2012, 65, 139–148. [Google Scholar] [CrossRef]
- Marcellos, H.; Perryman, T. Pollination and fertilization in crops of Vicia faba. Aust. J. Agric. Res. 1988, 39, 579–587. [Google Scholar] [CrossRef]
- Karaköy, T.; Baloch, F.S.; Toklu, F.; Özkan, H. Variation for selected morphological and quality-related traits among 178 faba bean landraces collected from Turkey. Plant Genet. Resour. 2013, 12, 5–13. [Google Scholar] [CrossRef]
- Velcheva, N.; Petrova, S. Statistical analysis of genetic diversity using faba bean landraces database. Agric. Sci. Technol. 2020, 12, 211–215. [Google Scholar] [CrossRef]
- Rebaa, F.; Abid, G.; Aouida, M.; Abdelkarim, S.; Aroua, I.; Muhovski, Y.; Baudoin, J.-P.; M’Hamdi, M.; Sassi, K.; Jebara, M. Genetic variability in Tunisian populations of faba bean (Vicia faba L. var. major) assessed by morphological and SSR markers. Physiol. Mol. Biol. Plants 2017, 23, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louwaars, N.P. Plant breeding and diversity: A troubled relationship? Euphytica 2018, 214, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurmansyah; Alghamdi, S.S.; Migdadi, H.M.; Khan, M.A.; Afzal, M. AFLP-based analysis of variation and population structure in mutagenesis induced faba bean. Diversity 2020, 12, 303. [Google Scholar] [CrossRef]
- Liu, Y.; Hou, W. Genetic Diversity of Faba Bean Germplasms in Qinghai and Core Germplasm Identified based on AFLP Analysis. Legum. Genom. Genet. 2010, 1. [Google Scholar] [CrossRef]
- Ouji, A.; El Bok, S.; Syed, N.H.; Abdellaoui, R.; Rouaissi, M.; Flavell, A.J.; El Gazzah, M. Genetic diversity of faba bean (Vicia faba L.) populations revealed by sequence specific amplified polymorphism (SSAP) markers. Afr. J. Biotechnol. 2012, 11, 2162–2168. [Google Scholar]
- Abid, G.; Mingeot, D.; Udupa, S.M.; Muhovski, Y.; Watillon, B.; Sassi, K.; M’hamdi, M.; Souissi, F.; Mannai, K.; Barhoumi, F.; et al. Genetic relationship and diversity analysis of Faba Bean (Vicia Faba L. var. minor) genetic resources using morpho-logical and microsatellite molecular markers. Plant Mol. Biol. 2015, 33, 1755–1767. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, T.; Guan, J.; Wang, S.; Wang, H.; Sun, X.; Zong, X. Development and characterization of 21 EST-derived mi-crosatellite markers in Vicia faba (faba bean). Am. J. Bot. 2011, 98, 22–24. [Google Scholar] [CrossRef]
- Gong, Y.-M.; Xu, S.-C.; Mao, W.-H.; Li, Z.-Y.; Hu, Q.-Z.; Zhang, G.-W.; Ding, J. Genetic Diversity Analysis of Faba Bean (Vicia faba L.) Based on EST-SSR Markers. Agric. Sci. China 2011, 10, 838–844. [Google Scholar] [CrossRef]
- Tekalign, A.; Derera, J.; Sibiya, J.; Mumm, R.H. Molecular analysis for genetic diversity and population structure of Ethiopian faba bean (Vicia faba L) accessions. Plant Biol. Crop Res. 2019, 1, 1–10. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Gnanasambandam, A.; Paull, J.; Torres, A.; Kaur, S.; Leonforte, T.; Li, H.; Zong, X.; Yang, T.; Materne, M. Impact of Molecular Technologies on Faba Bean (Vicia faba L.) Breeding Strategies. Agronomy 2012, 2, 132–166. [Google Scholar] [CrossRef] [Green Version]
- Suresh, S.; Park, J.H.; Cho, G.T.; Lee, H.S.; Baek, H.J.; Lee, S.Y.; Chung, J.W. Development and molecular characterization of 55 novel polymorphic cDNA-SSR markers in faba bean (Vicia faba L.) using 454 pyrosequencing. Molecules 2013, 18, 1844–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alemayehu, N.; Keneni, G. Genetic consequences of failure to control outcrossing in breeding often cross-pollinated pulse and oil crops in Ethiopia. Ethiop. J. Crop Sci. 2020, 7, 1–29. [Google Scholar]
- Marzinziga, B.; Brünjesb, L.; Biagionic, S.; Behlingc, H.; Linkb, W.; Westphal, C. Bee pollinators of faba bean (Vicia faba L.) differ in their foraging behaviour and pollination efficiency. Agr. Ecosyst. Environ. 2018, 264, 24–33. [Google Scholar] [CrossRef]
- Corrado, G.; La Mura, M.; Ambrosino, O.; Pugliano, G.; Varricchio, P.; Rao, R. Relationships of Campanian olive cultivars: Comparative analysis of molecular and phenotypic data. Genome 2009, 52, 692–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accessions | DF | FS | NF | PLH | NPPL | PL | NSP | SL | SW | HSW | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| VF A1 | 79.67 | ±1.15 a | 98.67 | ±1.15 a | 4.33 | ±0.90 bcd | 72.85 | ±8.51 ab | 6.87 | ±1.94 a | 15.93 | ±2.07 de | 4.83 | ±0.99 g | 2.14 | ±0.12 ab | 1.46 | ±0.12 b | 108.33 | ±1.53 a |
| VF A2 | 82.33 | ±2.31 abc | 102.67 | ±2.31 ab | 4.60 | ±0.63 cde | 82.79 | ±8.06 cdef | 8.14 | ±2.95 ab | 16.14 | ±2.81 de | 5.00 | ±0.83 g | 2.21 | ±0.20 b | 1.59 | ±0.20 cd | 171.67 | ±7.64 d |
| VF A3 | 85.33 | ±1.15 c | 107.67 | ±1.15 c | 4.47 | ±0.83 bcd | 90.12 | ±7.20 g | 16.84 | ±4.55 de | 13.30 | ±1.83 c | 3.13 | ±0.43 bc | 2.86 | ±0.17 f | 1.95 | ±0.17 i | 236.33 | ±3.21 h |
| VF A4 | 80.33 | ±1.15 a | 99.33 | ±1.15 a | 4.20 | ±0.77 abc | 86.58 | ±13.76 efg | 6.50 | ±2.52 a | 16.75 | ±3.17 e | 4.23 | ±1.14 f | 2.63 | ±0.24 e | 1.94 | ±0.24 i | 185.67 | ±9.02 e |
| VF A5 | 81.67 | ±1.53 abc | 101.67 | ±1.53 ab | 5.07 | ±0.96 ef | 82.27 | ±11.43 cde | 17.79 | ±4.12 e | 10.69 | ±1.31 a | 2.93 | ±0.69 ab | 2.04 | ±0.16 a | 1.39 | ±0.16 a | 128.00 | ±4.00 b |
| VF A6 | 83.67 | ±2.52 abc | 102.00 | ±2.52 ab | 4.40 | ±0.83 bcd | 88.53 | ±14.18 fg | 15.69 | ±9.56 cde | 10.91 | ±0.84 a | 3.33 | ±0.84 bc | 2.14 | ±0.11 ab | 1.54 | ±0.11 c | 152.33 | ±8.74 c |
| VF A8 | 80.33 | ±1.15 a | 100.00 | ±1.15 a | 4.80 | ±0.68 def | 83.93 | ±9.14 def | 6.97 | ±2.34 a | 15.58 | ±2.03 d | 4.23 | ±0.97 f | 2.44 | ±0.20 cd | 1.74 | ±0.20 gh | 193.33 | ±6.66 ef |
| VF A9 | 91.00 | ±2.65 d | 107.33 | ±2.65 c | 3.73 | ±0.80 a | 80.60 | ±9.54 cde | 14.45 | ±13.77 bcde | 16.60 | ±3.51 de | 4.17 | ±0.83 ef | 2.43 | ±0.15 cd | 1.67 | ±0.15 efg | 186.67 | ±3.51 e |
| VF A10 | 81.33 | ±2.52 abc | 101.67 | ±2.52 ab | 4.20 | ±0.77 abc | 77.63 | ±11.89 bc | 6.63 | ±1.37 a | 15.75 | ±2.41 de | 3.77 | ±1.04 de | 2.50 | ±0.21 d | 1.65 | ±0.21 def | 202.67 | ±5.86 f |
| VF A11 | 79.67 | ±1.15 a | 101.33 | ±1.15 ab | 3.93 | ±0.80 ab | 85.35 | ±11.85 efg | 12.64 | ±6.61 abcd | 11.49 | ±1.66 ab | 3.00 | ±0.69 ab | 2.64 | ±0.29 e | 1.70 | ±0.29 fg | 215.00 | ±5.00 g |
| VF A12 | 82.67 | ±1.53 abc | 108.00 | ±1.53 c | 4.60 | ±0.74 cde | 78.65 | ±7.81 bcd | 7.77 | ±4.41 ab | 12.50 | ±1.14 bc | 3.53 | ±0.68 cd | 2.88 | ±0.27 f | 1.90 | ±0.27 i | 239.33 | ±14.47 h |
| VF A13 | 81.00 | ±2.00 ab | 107.67 | ± 2.00 c | 4.73 | ±0.70 cdef | 78.36 | ±8.93 bcd | 10.44 | ±5.58 abcd | 11.32 | ±1.87 a | 3.27 | ±0.83 bc | 2.38 | ±0.21 c | 1.61 | ±0.21 cde | 138.00 | ±3.00 b |
| VF A14 | 85.00 | ±6.00 bc | 102.33 | ±6.00 ab | 5.20 | ±1.32 f | 83.37 | ±9.01 cdef | 8.92 | ±3.28 abc | 12.90 | ±2.37 c | 3.27 | ±1.11 bc | 2.66 | ±0.26 e | 1.79 | ±0.26 h | 193.67 | ±3.51 ef |
| VF A15 | 81.33 | ±2.52 abc | 105.00 | ±2.52 bc | 4.20 | ±0.77 abc | 68.12 | ±12.28 a | 17.51 | ±7.61 e | 10.84 | ±0.56 a | 2.60 | ±0.50 a | 2.95 | ±0.21 f | 2.13 | ±0.21 i | 185.33 | ±1.53 e |
| Means | 82.52 | 103.24 | 4.46 | 81.36 | 11.23 | 13.62 | 3.66 | 2.49 | 1.72 | 181.17 | ||||||||||
| SD | ±2.10 | ±2.27 | ±0.82 | ±11.49 | ±5.04 | ±3.12 | ±1.09 | ±0.35 | ±0.25 | ±5.55 | ||||||||||
| CV% | 2.54% | 2.20% | 18.45% | 14.13% | 44.57% | 22.88% | 29.89% | 13.95% | 14.39% | 3.14% | ||||||||||
| Significance | *** | *** | *** | *** | ** | *** | *** | *** | *** | *** | ||||||||||
| Na | Ne | Ho | He | I | Fst | PIC | |
|---|---|---|---|---|---|---|---|
| JF1AAC2 | 2 | 1.57 | 0.57 | 0.42 | 0.39 | −0.35 | 0.32 |
| JF1AG3 | 3 | 1.42 | 0.81 | 0.60 | 0.44 | −0.36 | 0.48 |
| JF1AG2 | 2 | 1.07 | 0.66 | 0.47 | 0.29 | −0.42 | 0.34 |
| VFG34 | 3 | 1.92 | 0.92 | 0.59 | 0.64 | −0.56 | 0.49 |
| GA6 | 3 | 1.71 | 0.71 | 0.58 | 0.49 | −0.23 | 0.46 |
| GAII67 | 4 | 1.42 | 1.00 | 0.67 | 0.49 | −0.47 | 0.58 |
| GATA11 | 3 | 0.78 | 0.37 | 0.49 | 0.14 | 0.24 | 0.39 |
| Total | 20 | ||||||
| Mean | 2.85 | 1.41 | 0.72 | 0.54 | 0.41 | −0.30 | 0.442 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malek, N.; Aci, M.M.; Khamassi, K.; Lupini, A.; Rouissi, M.; Hanifi-Mekliche, L. Agro-Morphological and Molecular Variability among Algerian Faba Bean (Vicia faba L.) Accessions. Agronomy 2021, 11, 1456. https://doi.org/10.3390/agronomy11081456
Malek N, Aci MM, Khamassi K, Lupini A, Rouissi M, Hanifi-Mekliche L. Agro-Morphological and Molecular Variability among Algerian Faba Bean (Vicia faba L.) Accessions. Agronomy. 2021; 11(8):1456. https://doi.org/10.3390/agronomy11081456
Chicago/Turabian StyleMalek, Nawel, Meriem Miyassa Aci, Khalil Khamassi, Antonio Lupini, Mustapha Rouissi, and Leila Hanifi-Mekliche. 2021. "Agro-Morphological and Molecular Variability among Algerian Faba Bean (Vicia faba L.) Accessions" Agronomy 11, no. 8: 1456. https://doi.org/10.3390/agronomy11081456