Exogenous Potassium Treatments Elevate Salt Tolerance and Performances of Glycine max L. by Boosting Antioxidant Defense System under Actual Saline Field Conditions


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Treatments, Design, and Plant Material of the Experiments
2.2. Measurements
2.2.1. Growth Traits and Photosynthesis Pigment Analyses
2.2.2. Extraction and Determination of Inorganic Solutes
2.2.3. Determination of Osmoprotectants and Non-Enzymatic Antioxidant Contents
2.2.4. Determination of Malondialdehyde and Hydrogen Peroxide Contents
2.2.5. Antioxidant Enzymes
2.2.6. Determination of Yield Attributes and Seed Quality
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Essa, T.; Al-Ani, D. Effect of salt stress on the performance of six soybean genotypes. Pak. J. Biol. Sci. 2001, 4, 175–177. [Google Scholar]
- Agarwal, A.K. Biofuels (alcohols and biodiesel) applications as fuels for internal combustion engines. Prog. Energy Combust. Sci. 2007, 33, 233–271. [Google Scholar] [CrossRef]
- Shi, G.; Cai, Q. Zinc tolerance and accumulation in eight oil crops. J. Plant Nutr. 2010, 33, 982–997. [Google Scholar] [CrossRef]
- Krishnan, H.B. Biochemistry and molecular biology of soybean seed storage proteins. J. New Seeds 2001, 2, 1–25. [Google Scholar] [CrossRef]
- Anonymous. Agricultural Statistics of Pakistan; Ministry of Food, Agriculture and Live Stock, (Economic Wing): Islamabad, Pakistan, 2009; pp. 65–66. [Google Scholar]
- FAOSTAT. Food and Agriculture Organization of the United Nations Statistics Division. 2020. Available online: http://faostat.fao.org/site/567/DesktopDefault.aspx (accessed on 20 September 2020).
- Seleiman, M.F.; Kheir, A.S. Saline soil properties, quality and productivity of wheat grown with bagasse ash and thiourea in different climatic zones. Chemosphere 2018, 193, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Kheir, A.S.; Ali, O.A.; Hafez, E.; Elshamey, E.A.; Zhou, Z.; Wang, B.; Lin, X.; Ge, Y.; Fahmy, A.E.; et al. A vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manag. 2020, 277, 111388. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Wang, X.; Ma, C. Grain legumes and fear of salt stress: Focus on mechanisms and management strategies. Int. J. Mol. Sci. 2019, 20, 799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rady, M.M.; Taha, R.S.; Mahdi, A.H.A. Proline enhances growth, productivity and anatomy of two varieties of Lupinus termis L. grown under salt stress. S. Afr. J. Bot. 2016, 102, 221–227. [Google Scholar] [CrossRef]
- Jha, U.C.; Bohra, A.; Jha, R.; Parida, S.K. Salinity stress response and ‘omics’ approaches for improving salinity stress tolerance in major grain legumes. Plant Cell Rep. 2019, 38, 255–277. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination; Plenum Press: New York, NY, USA, 1994. [Google Scholar]
- Mcdonald, M.B. Seed priming. In Seed Technology and Its Biological Basis; Black, M., Bewley, J.D., Eds.; Sheffield Academic Press: Sheffield, UK, 2000; pp. 287–325. [Google Scholar]
- Soeda, Y.; Konings, M.C.; Vorst, O.; van Houwelingen, A.M.; Stoopen, G.M.; Maliepaard, C.A.; Kodde, J.; Bino, R.J.; Groot, S.P.; van der Geest, A.H. Gene expression programs during Brassica oleracea seed maturation, osmopriming, and germination are indicators of progression of the germination process and the stress tolerance level. Plant Physiol. 2005, 137, 354–368. [Google Scholar] [CrossRef] [Green Version]
- El-Lethy, S.R.; Abdelhamid, M.T.; Reda, F. Effect of potassium application on wheat (Triticum aestivum L.) cultivars grown under salinity stress. World Appl. Sci. J. 2013, 26, 840–850. [Google Scholar]
- Seleiman, M.F. Use of plant nutrients in improving abiotic stress tolerance in wheat. In Wheat Production in Changing Environments: Management, Adaptation and Tolerance; Hasanuzzaman, M., Nahar, K., Hossain, A., Eds.; Springer Nature: Singapore, 2019; pp. 481–495. ISBN 978-981-13-6883-7. [Google Scholar] [CrossRef]
- Xu, X.; Du, X.; Wang, F.; Sha, J.; Chen, Q.; Tian, G.; Zhu, Z.; Ge, S.; Jiang, Y. Effects of Potassium Levels on Plant Growth, Accumulation and Distribution of Carbon, and Nitrate Metabolism in Apple Dwarf Rootstock Seedlings. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Waraich, E.A.; Ahmad, R.; Halim, A.; Aziz, T. Alleviation of temperature stress by nutrient management in crop plants: A review. J. Soil Sci. Plant Nutr. 2012, 12, 221–244. [Google Scholar] [CrossRef] [Green Version]
- Anaraki, Z.; Shariati, M.; Hosseini Tafreshi, S. Transient silencing of phytoene desaturase reveals critical roles on plant response to salinity stress. Acta Physiol. Plant. 2017, 39, 161. [Google Scholar] [CrossRef]
- Clark, A.J.; Landolt, W.; Bucher, J.B.; Strasser, R.J. Beech (Fagus sylvatica) response to ozone exposure assessed with a chlorophyll a fluorescence performance index. Environ. Pollut. 2000, 109, 501–507. [Google Scholar] [CrossRef]
- Malavolta, E.; Vitti, G.C.; Oliveira, S.A. Avaliação do Estudonutricional das Plantas, Princípios e Aplicações; POTAFOS: Piracicaba, Brazil, 1989; 97p. [Google Scholar]
- Gaines, T.P.; Parker, M.B.; Gascho, G.J. Automated determination of chlorides in soil and plant tissue by sodium nitrate Extraction 1. Agron. J. 1984, 76, 371–374. [Google Scholar] [CrossRef]
- Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing Adverse Effects of Pb on Maize Plants by Combined Treatment with Jasmonic, Salicylic Acids and Proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Emerich, D.W.; Sanchez-Diaz, M. Water stress induced changes in the concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 8, 455–460. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2 vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Konings, E.J.M.; Roomans, H.H.S.; Beljaars, P.R. Liquid chromatographic determination of tocopherols and tocotrienols in margarine, infant foods, and vegetables. J. AOAC Int. 1996, 79, 902–906. [Google Scholar] [CrossRef] [Green Version]
- Ching, L.S.; Mohamed, S. Alpha-tocopherol content in 62 edible tropical plants. J. Agric. Food Chem. 2001, 49, 3101–3105. [Google Scholar] [CrossRef]
- MadhavaRao, K.V.; Sresty, T.V.S. Antioxidative parameters in the seedlings of pigeonpea (Cajanuscajan L. Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Havir, E.A.; McHale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–868. [Google Scholar]
- Kar, M.; Mishra, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis of Association of Official Agricultural Chemists, 18th ed.; The Association of Official Analytical Chemists: Washington, DC, USA, 2010. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: Singapore, 1984; p. 680. [Google Scholar]
- Semida, W.M.; Taha, R.S.; Abdelhamid, M.T.; Rady, M.M. Foliar-applied ɑ-tocopherol enhances salt-tolerance in Vicia faba L. plants grown under saline conditions. S. Afr. J. Bot. 2014, 95, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Mahdi, A.H.A. Improvement of salt tolerance in Vicia faba (L.) Plants by exogenous application of polyamines. Egypt. J. Agron. 2016, 38, 1–21. [Google Scholar]
- Taha, R.S. Improving Salt Tolerance of Helianthus annuus (L.) Plants by Moringa oleifera Leaf extract. Egypt. J. Agron. 2016, 38, 117–140. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Gil, S.S.; Fujita, M. Drought stress responses in plants, oxidative stress and antioxidant defense. In Climate Change and Plant Abiotic Stress Tolerance; Gill, S.S., Tuteja, N., Eds.; Wiley: Weinheim, Germany, 2014; pp. 209–249. [Google Scholar]
- Hameed, A.; Goher, M.; Iqbal, N. Drought induced programmed cell death and associated changes in antioxidants, proteases, and lipid peroxidation in wheat leaves. Biologia Plant. 2013, 57, 370–374. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Semida, W.M.; Taha, R.S.; Rady, M.M. Effect of summer-fall deficit irrigation on morpho-physiological, anatomical responses, fruit yield and water use efficiency of cucumber under salt affected soil. Sci. Hortic. 2018, 237, 148–155. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxygen processing in photosynthesis: Regulation and signalling. New Phytol. 2000, 146, 359–388. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, J.; Wu, L. Methylglyoxal-induced mitochondrial dysfunction in vascular smooth muscle cells. Biochem. Pharmacol. 2009, 77, 1709–1716. [Google Scholar] [CrossRef]
- Desai, K.M.; Chang, T.; Wang, H.; Banigesh, A.; Dhar, A.; Liu, J.; Untereiner, A.; Wu, L. Oxidative stress and aging: Is methylglyoxal the hidden enemy? Can. J. Physiol. Pharmacol. 2010, 88, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Taha, R.S.; Alharby, H.F.; Bamagoos, A.A.; Medani, R.A.; Rady, M.M. Elevating tolerance of drought stress in Ocimum basilicum using pollen grains extract; a natural biostimulant by regulation of plant performance and antioxidant defense system. S. Afr. J. Bot. 2020, 128, 42–53. [Google Scholar] [CrossRef]
- Hernández, J.A.; Jiménez, A.; Mullineaux, P.; Sevilia, F. Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defences. Plant Cell Environ. 2000, 23, 853–862. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Ling, F.; Silberbush, M. Response of maize to foliar vs. soil application of nitrogen–phosphorus–potassium fertilizers. J. Plant Nutr. 2002, 25, 2333–2342. [Google Scholar] [CrossRef]
- Pettigrew, W.T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol. Plant. 2008, 133, 670–681. [Google Scholar] [CrossRef]
- Zörb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture-Status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; pp. 178–189. [Google Scholar]
- Shingles, R.; McCarty, R.E. Direct measurement of ATP-dependent proton concentration changes and characterization of a K-stimulated ATPase in pea chloroplast inner envelope vesicles. Plant Physiol. 1994, 106, 731–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Luo, W.; Lin, W.; Ma, L.; Kabir, M.H. Model of cation transportation mediated by high-affinity potassium transporters (HKTs) in higher plants. Biol. Proced. Online 2015, 17, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Shabala, S.N.; Lew, R.R. Turgor regulation in osmotically stressed Arabidopsis epidermal root cells. Direct support for the role of inorganic ion uptake as revealed by concurrent flux and cell turgor measurements. Plant Physiol. 2002, 129, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Munns, R.; Colmer, T.D. Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot. 2015, 115, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Shabala, L.; Zhang, J.; Pottosin, I.; Bose, J.; Zhu, M.; Fuglsang, A.T.; Velarde-Buendia, A.; Massart, A.; Hill, C.B.; Roessner, U.; et al. Cell-type specific H+-ATPase activity in root tissues enables K+ retention and mediates acclimation of barley (Hordeum vulgare L.) to salinity stress. Plant Physiol. 2016, 172, 2445–2458. [Google Scholar] [CrossRef] [Green Version]
- Fayez, K.A.; Bazaid, S.A. Improving drought and salinity tolerance in barley by application of salicylic acid and potassium nitrate. J. Saudi Soc. Agric. Sci. 2014, 13, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, K.; Bhaduri, D.; Meena, H.N.; Kalariya, K. External potassium (K+) application improves salinity tolerance by promoting Na+-exclusion, K+-accumulation and osmotic adjustment in contrasting peanut cultivars. Plant Physiol. Biochem. 2016, 103, 143–153. [Google Scholar] [CrossRef]
- Saneoka, H.; Nagasaka, C.; Hahn, D.T.; Yang, W.J.; Premachandra, G.S.; Joly, R.J.; Rhodes, D. Salt tolerance of glycinebetaine-defcient and -containing maize lines. Plant Physiol. 1995, 107, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Shomer, I.; Frenkel, H.; Polinger, C. The existence of a diffuse electric double layer at cellulose fbril surfaces and its role in the swelling mechanism of parenchyma plant cell walls. Carbohydr. Polym. 1991, 16, 199–210. [Google Scholar] [CrossRef]
- Rady, M.M.; Taha, R.S.; Semida, W.M.; Alharby, H.F. Modulation of salt stress effects on Vicia faba L. plants grown on a reclaimed-saline soil by salicylic acid application. Rom. Agric. Res. 2017, 34, 175–185. [Google Scholar]
- Anjum, N.A.; Aref, I.M.; Duarte, A.C.; Pereira, E.; Ahmad, I.; Iqbal, M. Glutathione and proline can coordinately make plants withstand the joint attack of metal (loid) and salinity stresses. Front. Plant Sci. 2014, 5, 662. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Ahmad, A.; Ansari, M.K.A.; Qureshi, M.I.; Aref, I.M.; Khan, P.R.; Hegazy, S.S.; El-Atta, H.; Husen, A.; Hakeem, K.R. Improving the phytoextraction capacity of plants to scavenge metal (loid)-contaminated sites. Environ. Rev. 2015, 23, 44–65. [Google Scholar] [CrossRef]
- Khan, M.N.; Siddiqui, M.H.; Mohammad, F.; Khan, M.M.A.; Naeem, M. Salinity induced changes in growth, enzyme activities, photosynthesis, proline accumulation and yield in linseed genotypes. World J. Agric. Sci. 2007, 3, 685–695. [Google Scholar]
- Qureshi, M.I.; Abdin, M.Z.; Ahmad, J.; Iqbal, M. Effect of long-term salinity on cellular antioxidants, compatible solute and fatty acid profile of Sweet annie (Artemisia annua L.). Phytochemistry 2013, 95, 215–223. [Google Scholar] [CrossRef]
- Anjum, N.A.; Ahmad, I.; Mohmood, I.; Pacheco, M.; Duarte, A.C.; Pereira, E.; Umar, S.; Ahmad, A.; Khan, N.A.; Iqbal, M.; et al. Modulation of glutathione and its related enzymes in plants’ responses to toxic metals and metalloids—a review. Environ. Exp. Bot. 2011, 75, 307–324. [Google Scholar] [CrossRef]
- Anjum, N.A.; Sofo, A.; Scopa, A.; Roychoudhury, A.; Gill, S.S.; Iqbal, M.; Lukatkin, A.S.; Pereira, E.; Duarte, A.C.; Ahmad, I. Lipids and proteins-major targets of oxidative modifications in abiotic stressed plants. Environ. Sci. Pollut. Res. 2015, 22, 4099–4121. [Google Scholar] [CrossRef]
- Liang, T.B.; Wang, Z.L.; Wang, R.J.; Liu, L.L.; Shi, C.Y. Effects of potassium humate on ginger root growth and its active oxygen metabolism. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2007, 18, 813–817. [Google Scholar]
- Zheng, Y.; Jia, A.; Ning, T.; Xu, J.; Li, Z.; Jiang, G. Potassium nitrate application alleviates sodium chloride stress in winter wheat cultivars differing in salt tolerance. J. Plant Physiol. 2008, 165, 1455–1465. [Google Scholar] [CrossRef]
- Jan, A.U.; Hadi, F.; Nawaz, M.A.; Rahman, K. Potassium and zinc increase tolerance to salt stress in wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2017, 116, 139–149. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Particle Size Distribution | FC | pH | ECe dS m−1 | CaCO3 % | OM % | Available Macro-Nutrients (ppm) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sand % | Silt % | Clay % | Texture class | N | P | K | |||||
| Normal soil (Location 1) | |||||||||||
| 25.9 | 27.0 | 47.1 | Clay | 29.2 | 7.45 | 2.68 | 3.98 | 1.02 | 242 | 4.26 | 266 |
| Saline soil (Location 2) | |||||||||||
| 28.7 | 29.3 | 42.0 | Clay | 27.2 | 7.62 | 7.46 | 3.45 | 0.86 | 198 | 1.92 | 162 |
| Parameters | Normal (N) (Control) | Treatments | ||||
|---|---|---|---|---|---|---|
| N+ K(SS) | N+ K (FS) | Salinity (S) | S+ K(SS) | S+ K (FS) | ||
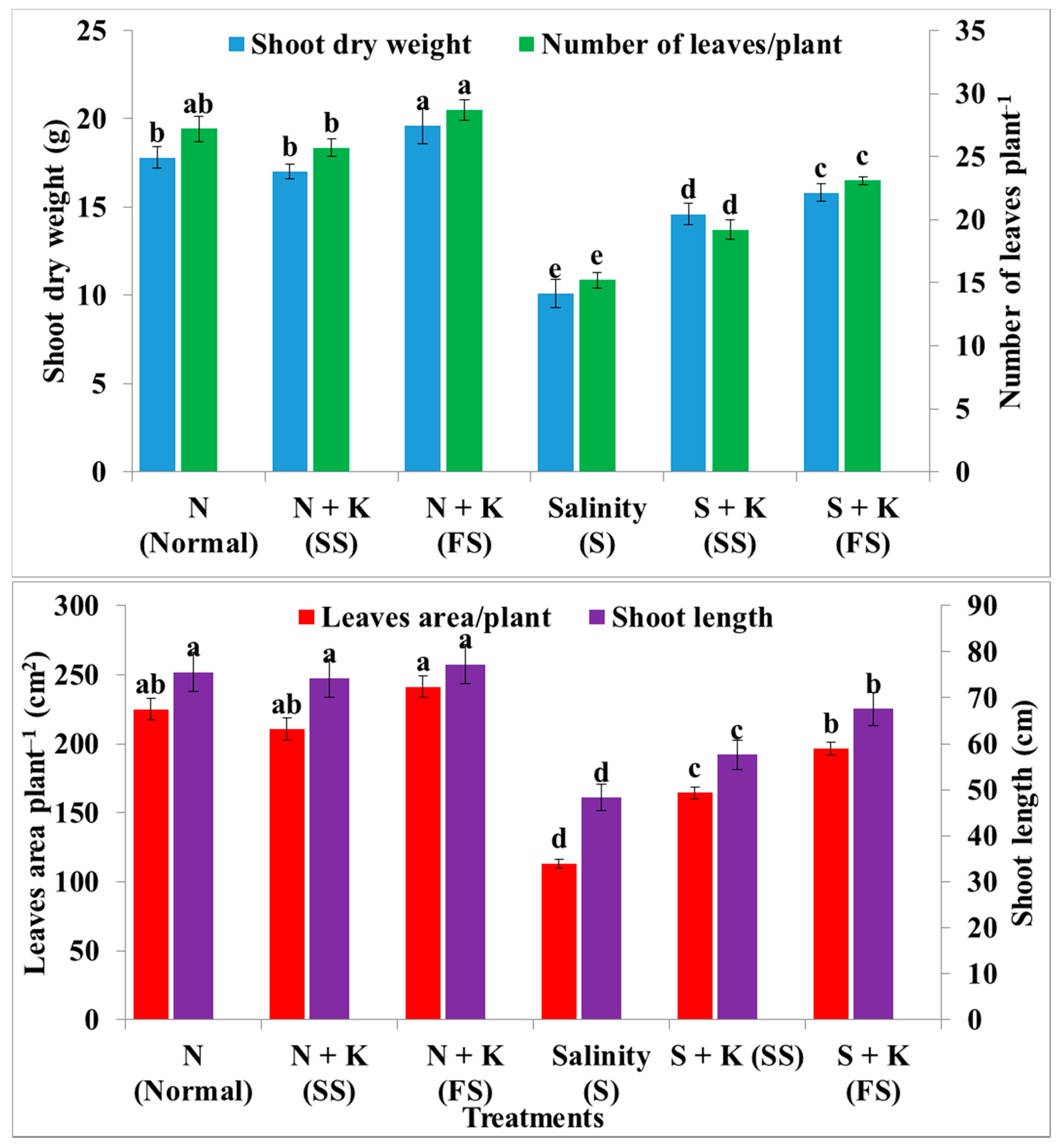
| Shoot length | a | −1.9a | +2.2a | −36.1d | −23.8c | −10.6b |
| Number of leaves plant‒1 | ab | −5.5b | +5.5a | −44.1d | −29.4c | −15.1b |
| Leaves area plant‒1 | ab | −19.8ab | +7.2a | −49.7d | −27.0c | −12.7b |
| Shoot dry weight | b | −4.5b | +10.1a | −43.3e | −18.0d | −11.2c |
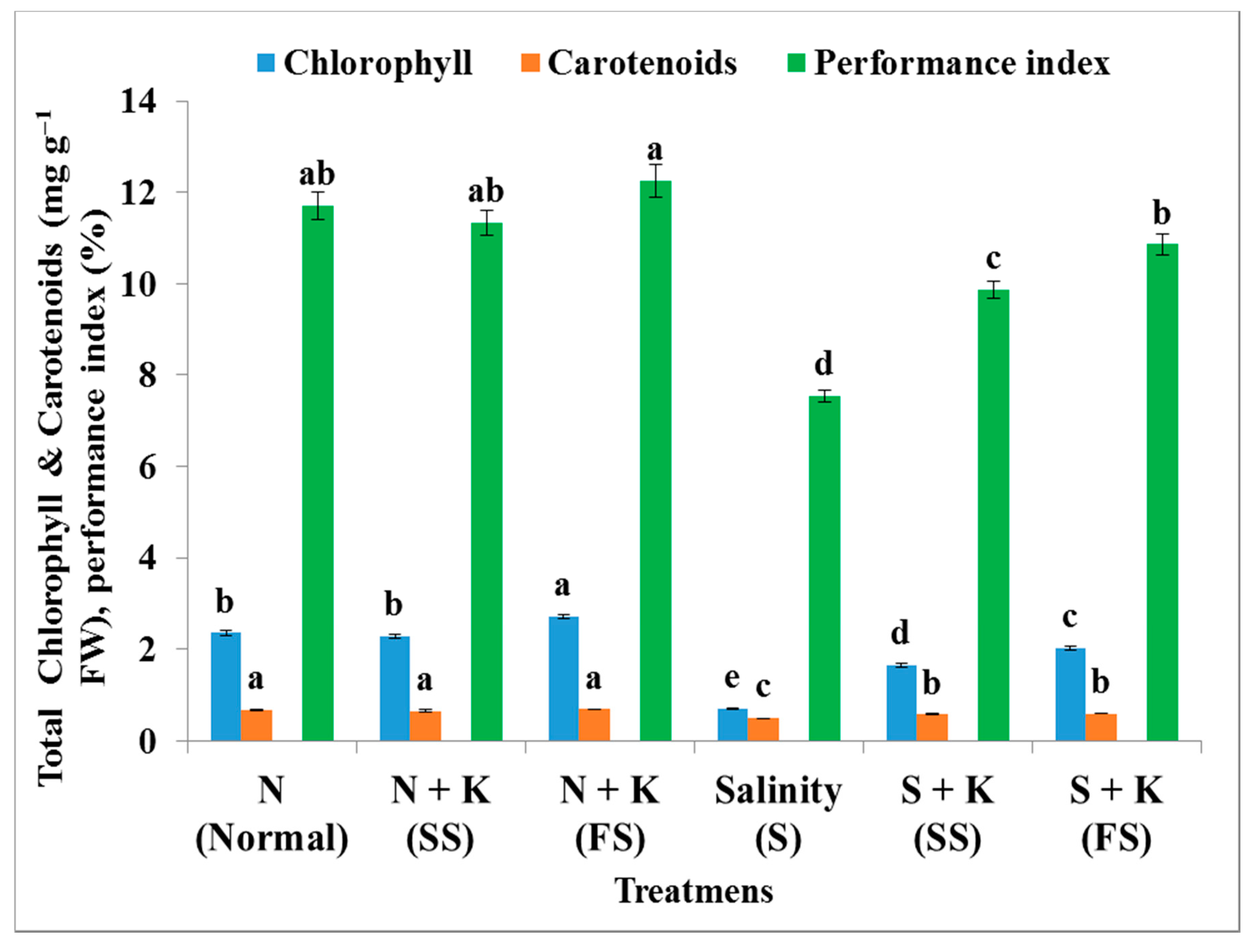
| Total chlorophylls content | b | −3.4b | +15.3a | −69.9e | −29.7d | −14.4c |
| Total carotenoids content | a | −1.5a | +3.0a | −25.4c | −13.4b | −10.4b |
| Performance index | ab | −3.2ab | +4.8a | −35.5d | −15.7c | −7.2b |
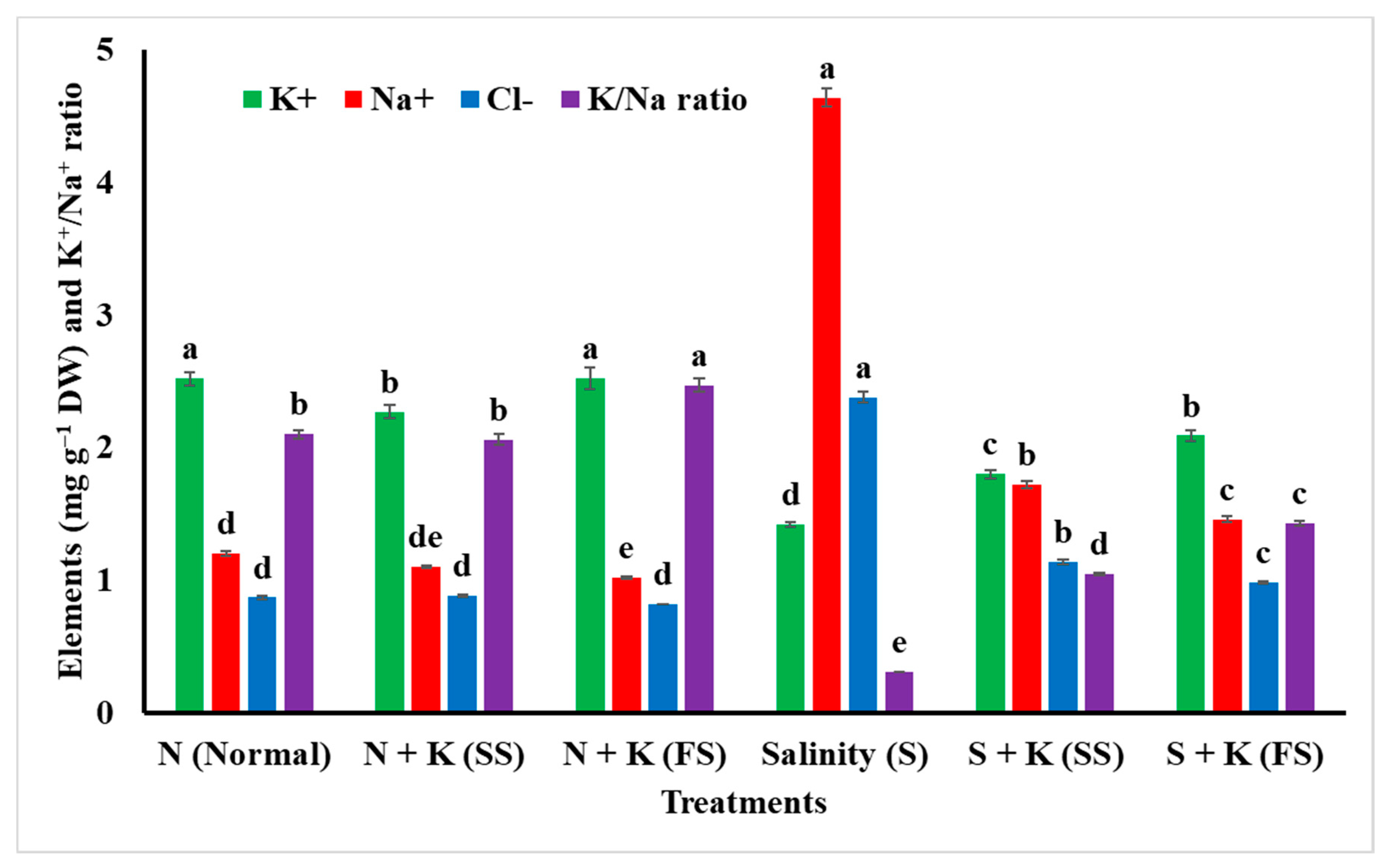
| Potassium (K+) content | a | −9.9b | 0.0a | −43.7d | −28.6c | −17.1b |
| Sodium (Na+) content | d | −8.3de | −15.0e | +286.7a | +43.3b | +21.7c |
| Chlorine (Cl‒) content | d | +1.1d | −5.7d | +173.6a | +31.0b | +12.6c |
| K+/Na+ ratio | b | −1.9b | +17.6a | −85.2e | −50.0d | −31.9c |
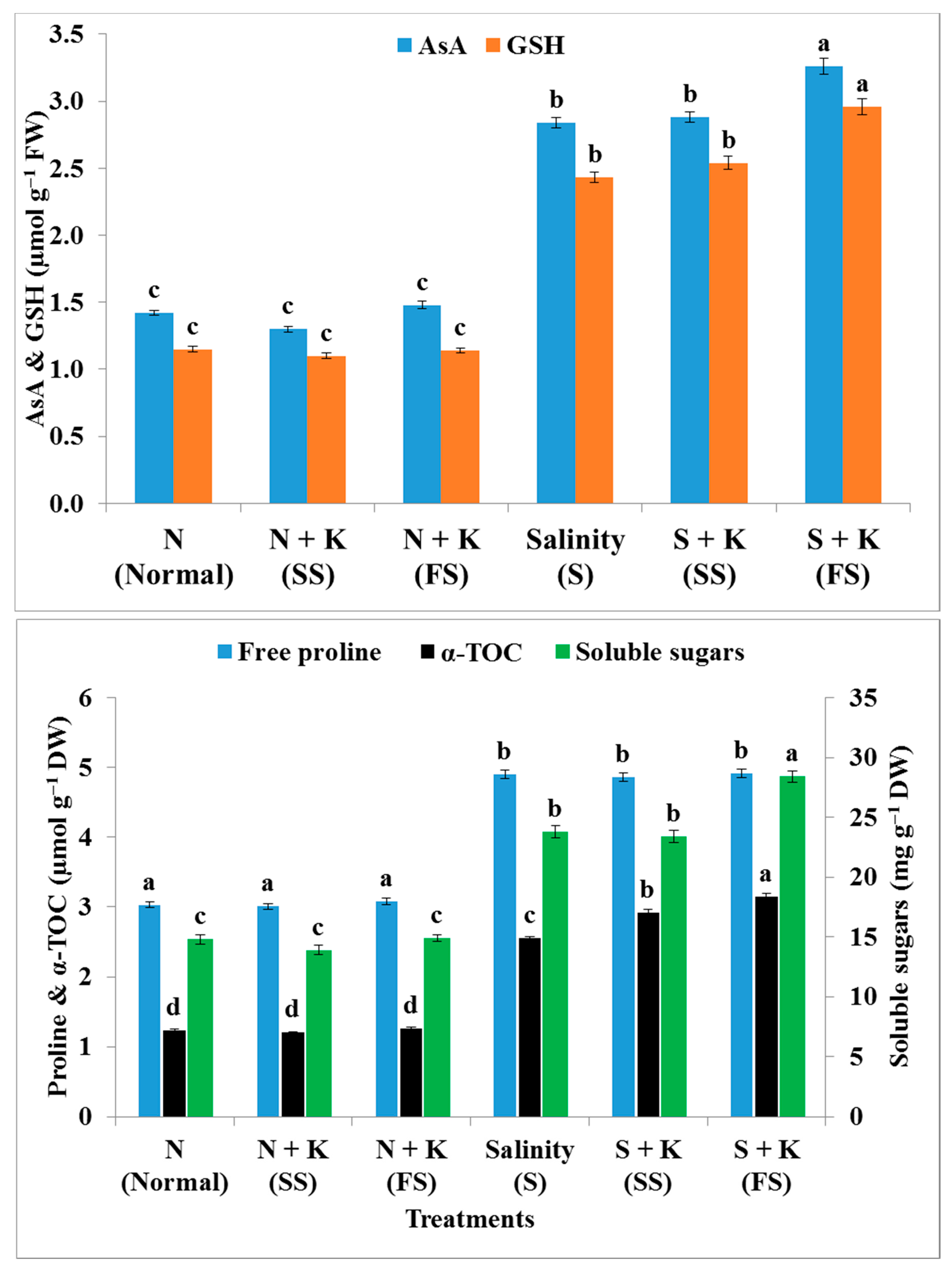
| Free proline content | b | −0.7b | +1.7b | +61.7a | +60.4a | +62.4a |
| Soluble sugars content | c | −6.1c | +0.7c | +60.8b | +58.1b | +91.9a |
| Ascorbate (AsA) content | c | −8.5c | +4.2c | +100.0b | +102.8b | +129.6a |
| Glutathione (GSH) content | c | −4.3c | −0.9c | +111.3b | +120.9b | +157.4a |
| α-Tocopherol content | d | −2.4d | +2.4d | +107.3c | +137.4b | +156.1a |
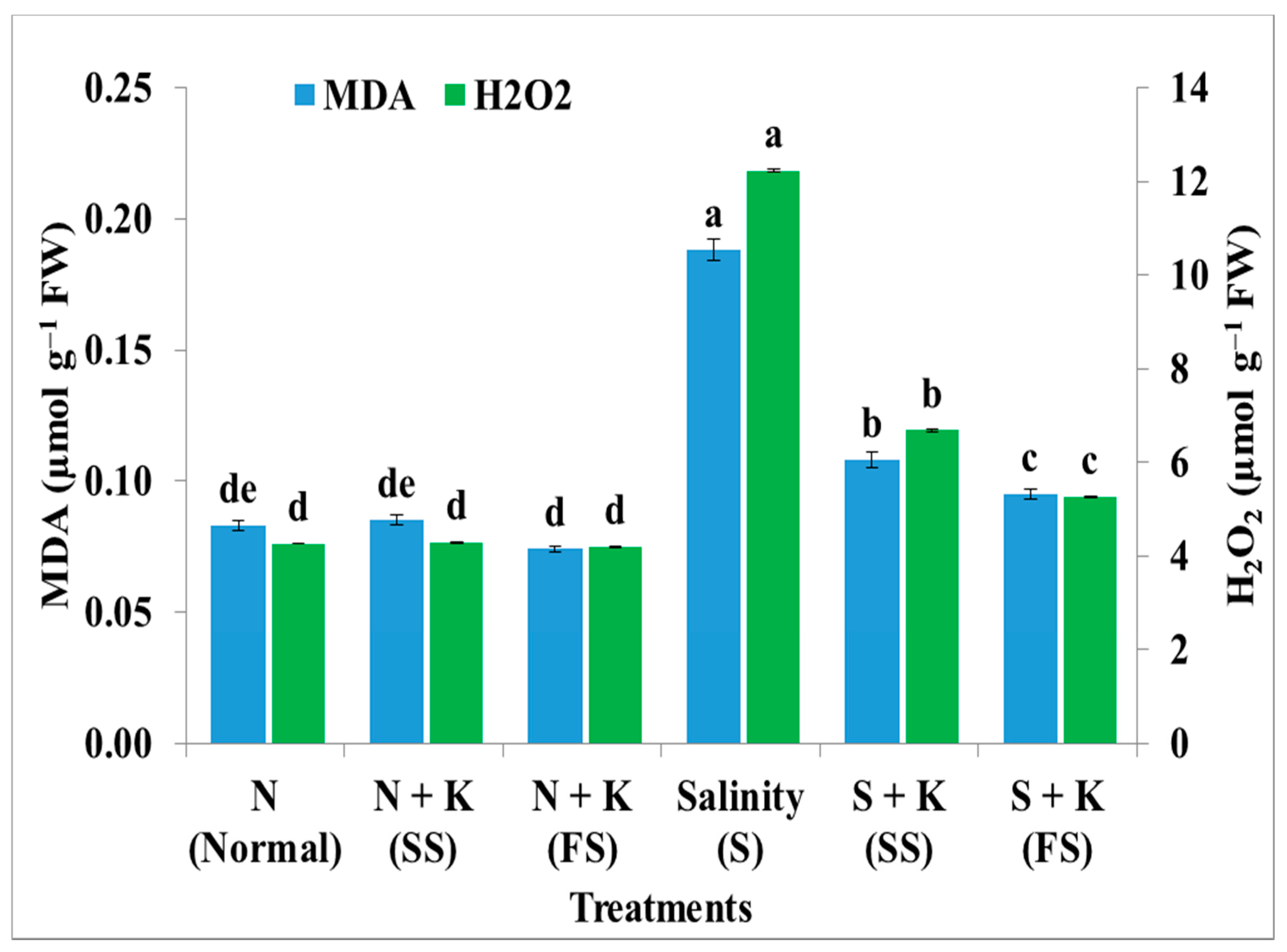
| Malondialdehyde level | de | +2.4de | −10.8e | +126.5a | +30.1b | +14.5c |
| Hydrogen peroxide level | d | +0.7d | −1.6d | +187.1a | +56.8b | +23.5c |
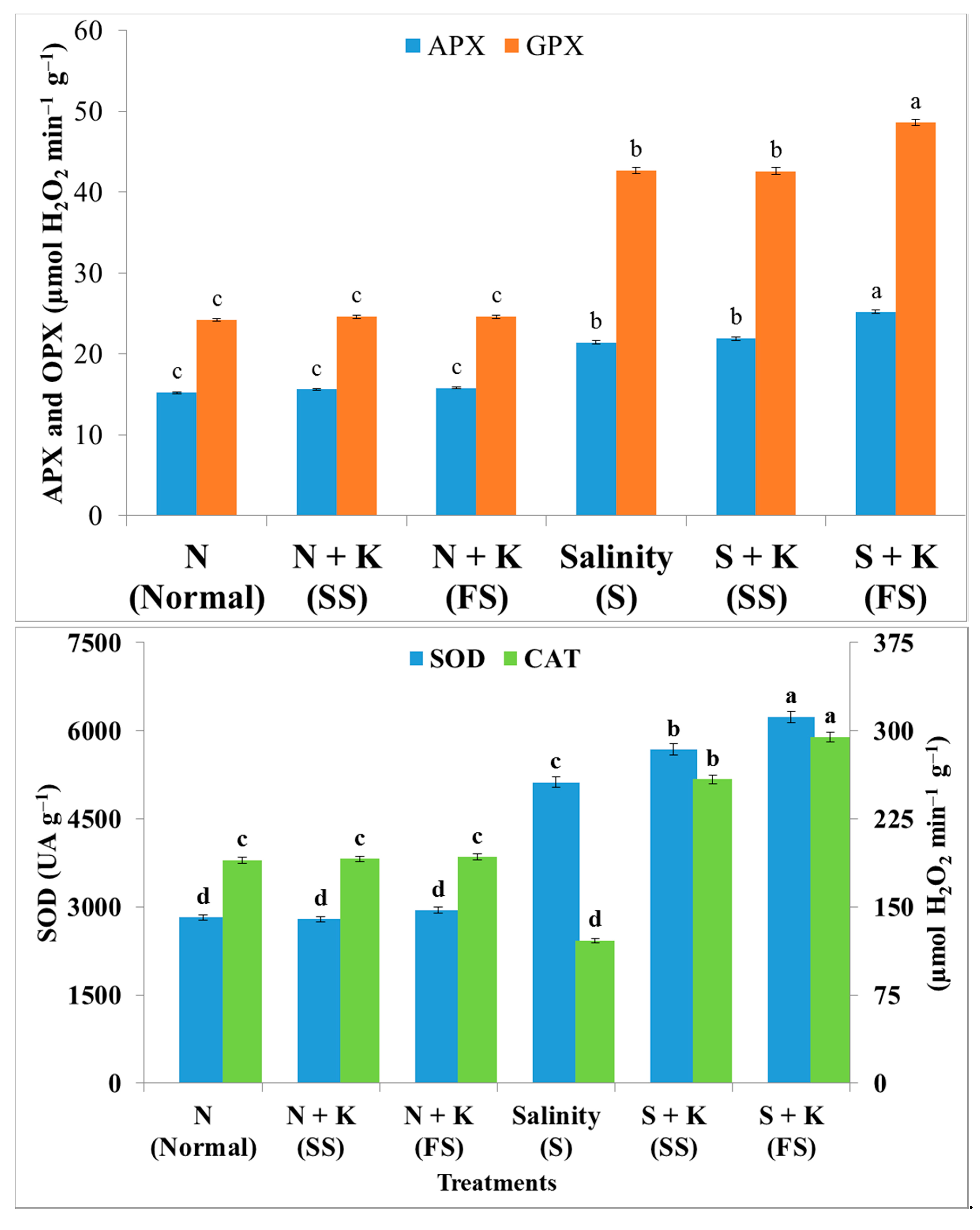
| SOD activity | d | −1.1d | +4.3d | +81.6c | +101.4b | +121.3a |
| CAT activity | c | +0.5c | +1.4c | −36.2d | +36.2b | +55.1a |
| APX activity | c | +2.6c | +3.9c | +40.8b | +44.1b | +65.8a |
| GPX activity | c | +1.7c | +1.7c | +76.4b | +76.0b | +100.8a |
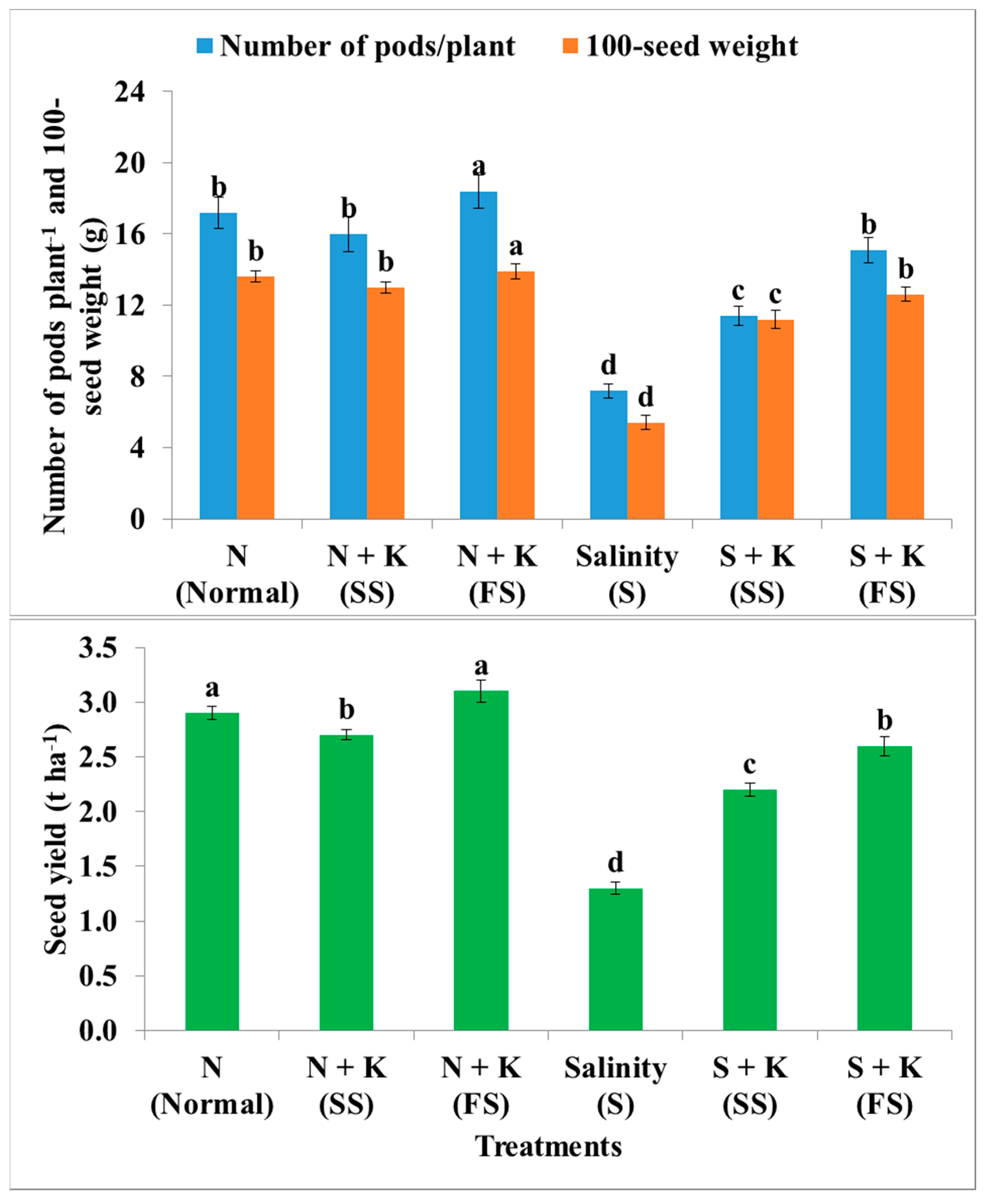
| Number of pods plant‒1 | b | −7.0c | +7.0a | −58.1e | −33.7d | −12.2c |
| 100-seed weight | a | −4.4ab | +2.2a | −60.3d | −17.6c | −7.4b |
| Seed yield | a | −6.9b | +6.9a | −55.2d | −24.1c | −13.8b |
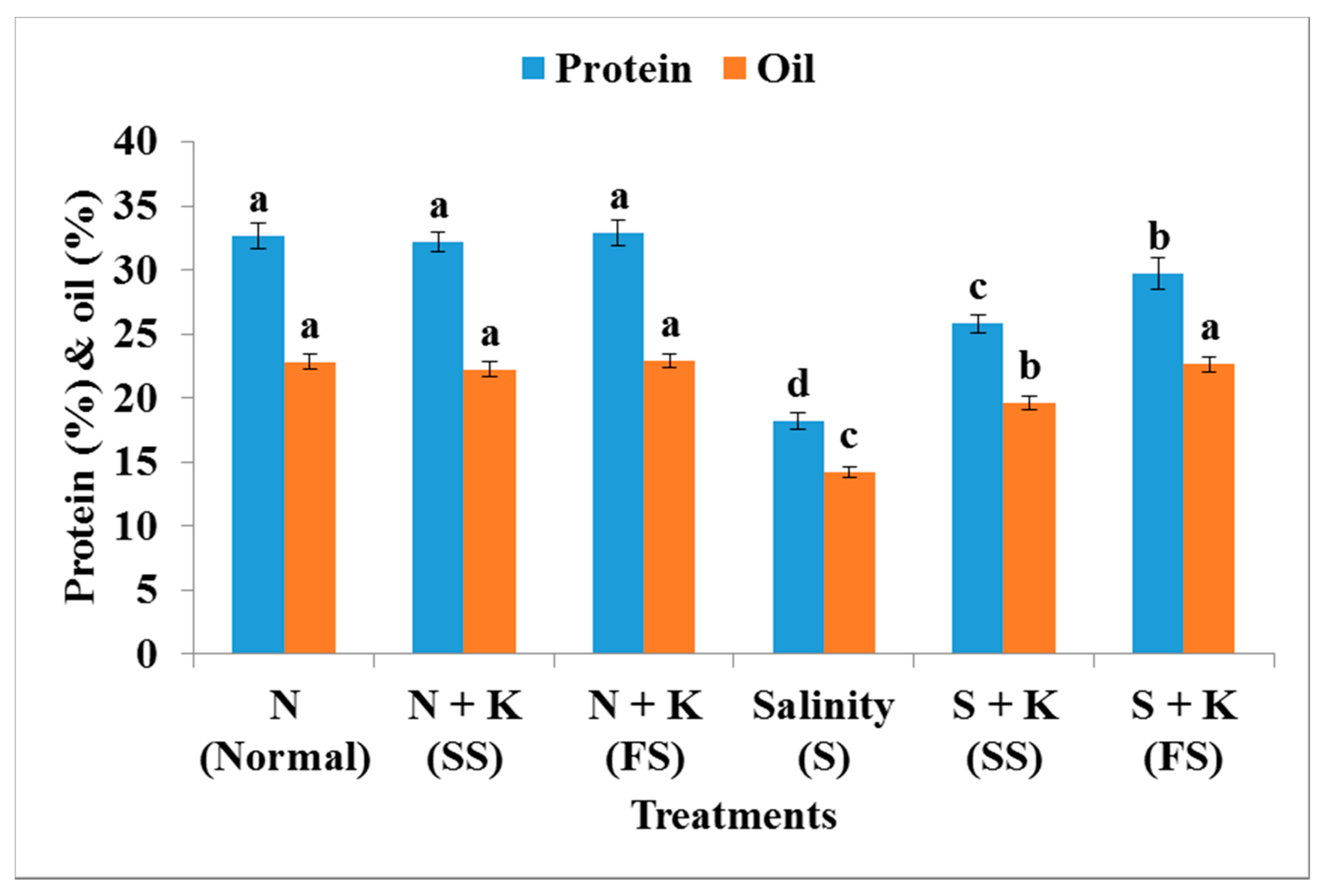
| Protein content (%) | a | −1.2a | +0.9a | −44.2d | −20.9c | −8.9b |
| Oil content (%) | a | −2.6a | +0.4a | −37.7c | −14.0b | −0.9a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
S. Taha, R.; Seleiman, M.F.; Alotaibi, M.; Alhammad, B.A.; Rady, M.M.; H. A. Mahdi, A. Exogenous Potassium Treatments Elevate Salt Tolerance and Performances of Glycine max L. by Boosting Antioxidant Defense System under Actual Saline Field Conditions. Agronomy 2020, 10, 1741. https://doi.org/10.3390/agronomy10111741
S. Taha R, Seleiman MF, Alotaibi M, Alhammad BA, Rady MM, H. A. Mahdi A. Exogenous Potassium Treatments Elevate Salt Tolerance and Performances of Glycine max L. by Boosting Antioxidant Defense System under Actual Saline Field Conditions. Agronomy. 2020; 10(11):1741. https://doi.org/10.3390/agronomy10111741
Chicago/Turabian StyleS. Taha, Ragab, Mahmoud F. Seleiman, Majed Alotaibi, Bushra Ahmed Alhammad, Mostafa M. Rady, and Ayman H. A. Mahdi. 2020. "Exogenous Potassium Treatments Elevate Salt Tolerance and Performances of Glycine max L. by Boosting Antioxidant Defense System under Actual Saline Field Conditions" Agronomy 10, no. 11: 1741. https://doi.org/10.3390/agronomy10111741