Evidence of the Role of QTL Epistatic Interactions in the Increase of Melon Fruit Flesh Content during Domestication

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Genotyping
2.3. Experimental Designs
2.4. Data Analysis
3. Results
3.1. QTL Analysis in the 14M2-F2 Population
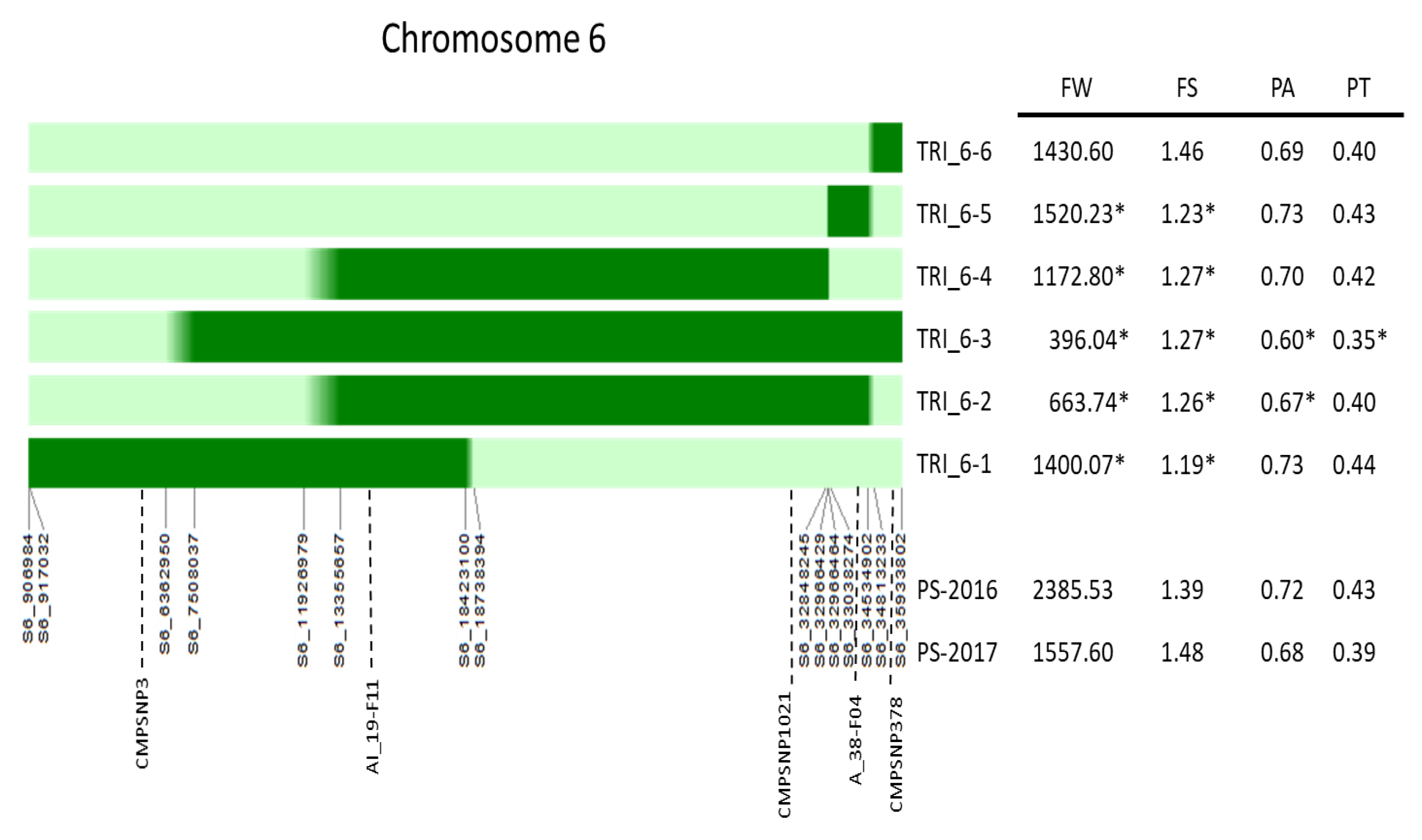
3.2. Analysis of Recombinant Introgression Lines
3.3. Effects of Other Genomic Regions on Flesh Content
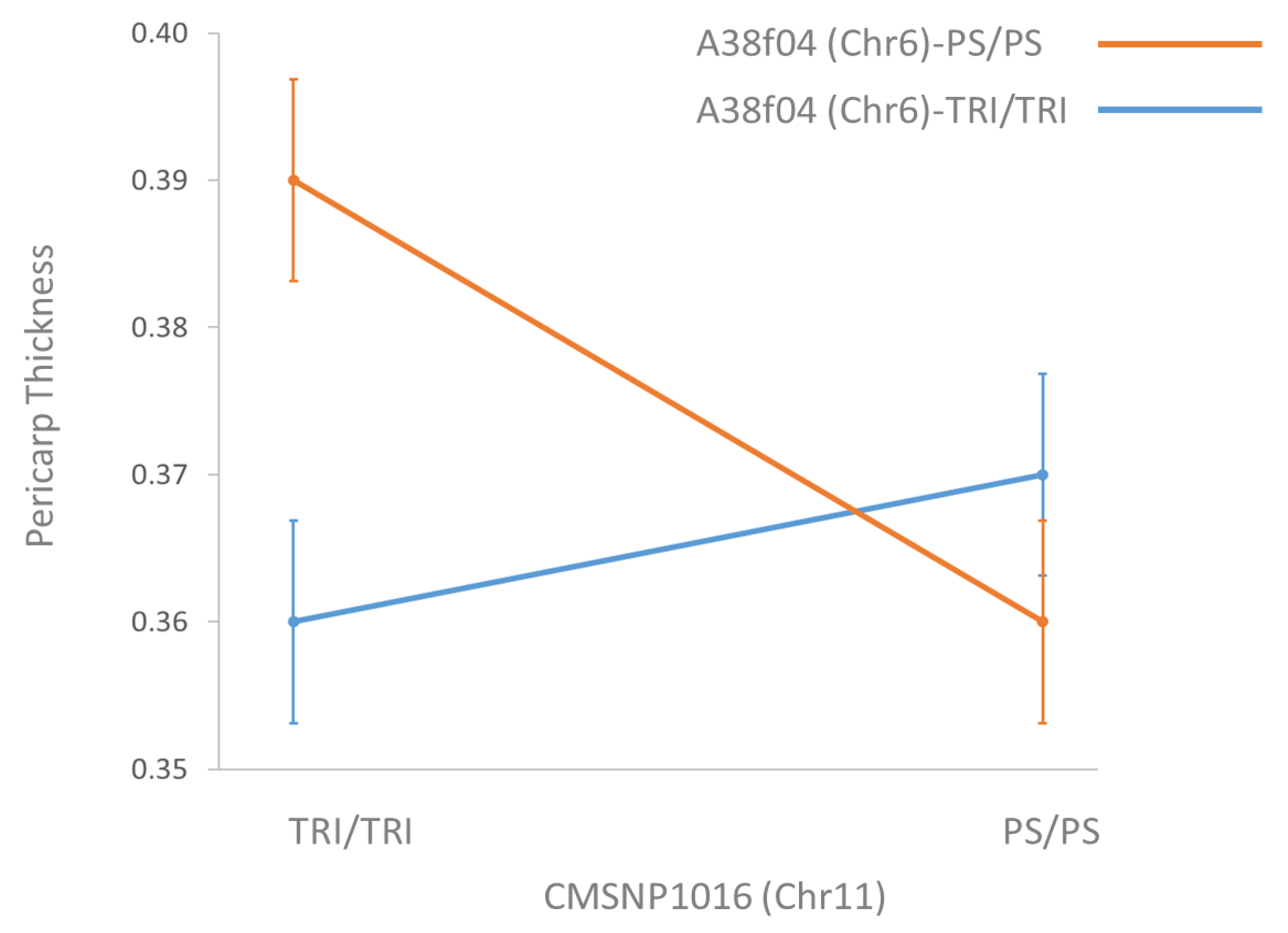
3.4. Verification of Epistatic Interaction
4. Discussion
4.1. paqt6.1 Effects Are Modulated by an Epistatic Locus
4.2. Epistasis and Melon Domestication
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Nomenclature
| ANOVA | analysis of variance |
| bHLH | basic helix-loop-helix |
| CV | coefficient of variation |
| FS | fruit shape |
| FW | fruit weight |
| GBS | genotyping by sequencing |
| IL | introgression line |
| LOD | logarithm of odds |
| PA | pericarp area |
| PS | Piel de Sapo |
| PT | pericarp thickness |
| QTL | quantitative trait locus |
| SNP | single nucleotide polymorphism |
| TRI | trigonus |
References
- Pitrat, M. Melon genetic resources: Phenotypic diversity and horticultural taxonomy. In Genetics and Genomics of Cucurbitaceae; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer Nature: Cham, Switzerland, 2017; pp. 25–60. [Google Scholar]
- Gonzalo, M.J.; Diaz, A.; Dhillon, N.P.S.; Reddy, U.K.; Pico, B.; Monforte, A.J. Re-evaluation of the role of Indian germplasm as center of melon diversification based on genotyping-by-sequencing analysis. BMC Genom. 2019, 20, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renner, S.S.; Schaefer, H. Phylogeny and evolution of the Cucurbitaceae. In Genetics and Genomics of Cucurbitaceae; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer Nature: Cham, Switzerland, 2017; pp. 13–24. [Google Scholar]
- Whitaker, T.W.; Davis, G.N. Cucurbits: Botany, Cultivation, and Utilization; Interscience Publishers: New York, NY, USA, 1962. [Google Scholar]
- Telford, I.R.H.; Schaefer, H.; Greuter, W.; Renner, S.S. A new Australian species of Luffa (Cucurbitaceae) and typification of two Australian Cucumis names, all based on specimens collected by Ferdinand Mueller in 1856. Phytokeys 2011, 5, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Telford, I.R.H.; Sebastian, P.; Bruhl, J.J.; Renner, S.S. Cucumis (Cucurbitaceae) in Australia and Eastern Malesia, including newly recognized species and the sister species to C. melo. Syst. Bot. 2011, 36, 376–389. [Google Scholar] [CrossRef]
- Filipowicz, N.; Schaefer, H.; Renners, S.S. Revisiting Luffa (Cucurbitaceae) 25 years after c. Heiser: Species boundaries and application of names tested with plastid and nuclear DNA sequences. Syst. Bot. 2014, 39, 205–215. [Google Scholar] [CrossRef]
- Clarke, A.C.; Burtenshaw, M.K.; McLenachan, P.A.; Erickson, D.L.; Penny, D. Reconstructing the origins and dispersal of the polynesian bottle gourd (Lagenaria siceraria). Mol. Biol. Evol. 2006, 23, 893–900. [Google Scholar] [CrossRef]
- Kistler, L.; Montenegro, A.; Smith, B.D.; Gifford, J.A.; Green, R.E.; Newsom, L.A.; Shapiro, B. Transoceanic drift and the domestication of African bottle gourds in the Americas. Proc. Natl. Acad. Sci. USA 2014, 111, 2937–2941. [Google Scholar] [CrossRef] [Green Version]
- Levi, A.; Jarret, R.; Kousik, S.; Wechter, W.P.; Nimmakayala, P.; Reddy, U.K. Genetic resources of watermelon. In Genetics and Genomics of Cucurbitaceae; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer Nature: Cham, Switzerland, 2017; pp. 87–110. [Google Scholar]
- Paris, S.P. Genetic resources of pumpkins and squash, Cucurbita spp. In Genetics and Genomics of Cucurbitaceae; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer Nature: Cham, Switzerland, 2017; pp. 111–154. [Google Scholar]
- Sebastian, P.; Schaefer, H.; Telford, I.R.H.; Renner, S.S. Cucumber (Cucumis sativus) and melon (C. melo) have numerous wild relatives in Asia and Australia, and the sister species of melon is from Australia. Proc. Natl. Acad. Sci. USA 2010, 107, 14269–14273. [Google Scholar] [CrossRef] [Green Version]
- Endl, J.; Achigan-Dako, E.G.; Pandey, A.K.; Monforte, A.J.; Pico, B.; Schaefer, H. Repeated domestication of melon (Cucumis melo) in Africa and Asia and a new close relative from India. Am. J. Bot. 2018, 105, 1662–1671. [Google Scholar] [CrossRef] [Green Version]
- Esteras, C.; Formisano, G.; Roig, C.; Díaz, A.; Blanca, J.; Garcia-Mas, J.; Gomez-Guillamon, M.L.; Lopez-Sese, A.I.; Lazaro, A.; Monforte, A.J.; et al. SNP genotyping in melons: Genetic variation, population structure, and linkage disequilibrium. Theor. Appl. Genet. 2013, 126, 1285–1303. [Google Scholar] [CrossRef]
- Zhao, G.W.; Lian, Q.; Zhang, Z.H.; Fu, Q.S.; He, Y.H.; Ma, S.; Ruggieri, V.; Monforte, A.J.; Wang, P.Y.; Julca, I.; et al. A comprehensive genome variation map of melon identifies multiple domestication events and loci influencing agronomic traits. Nat. Genet. 2019, 51, 1607–1615. [Google Scholar] [CrossRef]
- Roy, A.; Bal, S.S.; Fergany, M.; Kaur, S.; Singh, H.; Malik, A.A.; Singh, J.; Monforte, A.J.; Dhillon, N.P.S. Wild melon diversity in India (Punjab State). Gen. Res. Crop Evol. 2012, 59, 755–767. [Google Scholar] [CrossRef]
- Díaz, A.; Martin-Hernandez, A.M.; Dolcet-Sanjuan, R.; Garces-Claver, A.; Alvarez, J.M.; Garcia-Mas, J.; Pico, B.; Monforte, A.J. Quantitative trait loci analysis of melon (Cucumis melo L.) domestication-related traits. Theor. Appl. Genet. 2017, 130, 1837–1856. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mas, J.; Monforte, A.J.; Arús, P. Phylogenetic relationships among Cucumis species based on the ribosomal internal transcribed spacer sequence and microsatellite markers. Plant Syst. Evol. 2004, 248, 191–203. [Google Scholar] [CrossRef]
- Studer, A.J.; Zhao, Q.; Ross-Ibarra, J.; Doebley, J. Identification of a functional transposon insertion in the maize domestication gene tb1. Nat. Genet. 2011, 43, 1160–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.B.; Zhou, A.L.; Sang, T. Rice domestication by reducing shattering. Science 2006, 311, 1936–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frary, A.; Nesbitt, T.C.; Grandillo, S.; van der Knaap, E.; Cong, B.; Liu, J.P.; Meller, J.; Elber, R.; Alpert, K.B.; Tanksley, S.D. fw2.2: A quantitative trait locus key to the evolution of tomato fruit size. Science 2000, 289, 85–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Sanseverino, W.; Henaff, E.; Vives, C.; Pinosio, S.; Burgos-Paz, W.; Morgante, M.; Ramos-Onsins, S.E.; Garcia-Mas, J.; Casacuberta, J. Transposon insertions, structural variations, and SNPs contribute to the evolution of the melon genome. Mol. Biol. Evol. 2015, 32, 2760–2774. [Google Scholar] [CrossRef] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Mas, J.; Benjak, A.; Sanseverino, W.; Bourgeois, M.; Mir, G.; Gonzalez, V.M.; Henaff, E.; Camara, F.; Cozzuto, L.; Lowy, E.; et al. The genome of melon (Cucumis melo L.). Proc. Natl. Acad. Sci. USA 2012, 109, 11872–11877. [Google Scholar] [CrossRef] [Green Version]
- Brewer, M.T.; Lang, L.X.; Fujimura, K.; Dujmovic, N.; Gray, S.; van der Knaap, E. Development of a controlled vocabulary and software application to analyze fruit shape variation in tomato and other plant species. Plant Phys. 2006, 141, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lander, E.S.; Botstein, D. Mapping Mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics 1989, 121, 185–199. [Google Scholar] [PubMed]
- Kosambi, D.D. The estimation of map distances from recombination values. Ann. Eug. 1943, 12, 172–175. [Google Scholar] [CrossRef]
- Zeng, Z.B. Precision mapping of quantitative trait loci. Genetics 1994, 136, 1457–1468. [Google Scholar] [PubMed]
- Wang, S.; Basten, C.J.; Zeng, Z.B. Windows QTL Cartographer V2.5_011. Available online: http://statgen.ncsu.edu/qtlcart/WQTLCart.htm (accessed on 22 July 2020).
- Argyris, J.M.; Pujol, M.; Martin-Hernandez, A.M.; Garcia-Mas, J. Combined use of genetic and genomics resources to understand virus resistance and fruit quality traits in melon. Phys. Plant 2015, 155, 4–11. [Google Scholar] [CrossRef]
- Sun, S.; Wang, X.T.; Wang, K.T.; Cui, X. Dissection of complex traits of tomato in the post-genome era. Theor. Appl. Genet. 2020, 133, 1763–1776. [Google Scholar] [CrossRef]
- Bateson, W. Heredity and variation in modern lights. In Darwin and Modern Science; Seward, A.C., Ed.; Cambridge University Press: Cambridge, UK, 1909; pp. 85–101. [Google Scholar]
- Fisher, R.A. The correlation between relatives on the supposition of Mendelian inheritance. Trans. R. Soc. Edinb. 1918, 52, 399–433. [Google Scholar] [CrossRef] [Green Version]
- Eshed, Y.; Zamir, D. Less-than-additive epistatic interactions of quantitative trait loci in tomato. Genetics 1996, 143, 1807–1817. [Google Scholar]
- Causse, M.; Chaib, J.; Lecomte, L.; Buret, M.; Hospital, F. Both additivity and epistasis control the genetic variation for fruit quality traits in tomato. Theor. Appl. Genet. 2007, 115, 429–442. [Google Scholar] [CrossRef]
- Shang, L.; Liang, Q.; Wang, Y.; Zhao, Y.; Wang, K.; Hua, J. Epistasis together with partial dominance, over-dominance and QTL by environment interactions contribute to yield heterosis in upland cotton. Theor. Appl. Genet. 2016, 129, 1429–1446. [Google Scholar] [CrossRef]
- Würschum, T.; Maurer, H.P.; Schulz, B.; Moehring, J.; Reif, J.C. Genome-wide association mapping reveals epistasis and genetic interaction networks in sugar beet. Theor. Appl. Genet. 2011, 123, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Monforte, A.J.; Oliver, M.; Gonzalo, M.J.; Alvarez, J.M.; Dolcet-Sanjuan, R.; Arus, P. Identification of quantitative trait loci involved in fruit quality traits in melon (Cucumis melo L.). Theor. Appl. Genet. 2004, 108, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Clayberg, C. Interaction and linkage test of flesh color genes in Cucumis melo L. Rep. Cucurbit Genet. Coop. 1992, 15, 53. [Google Scholar]
- Vegas, J.; Garcia-Mas, J.; Monforte, A.J. Interaction between QTLs induces an advance in ethylene biosynthesis during melon fruit ripening. Theor. Appl. Genet. 2013, 126, 1531–1544. [Google Scholar] [CrossRef]
- Guiu-Aragones, C.; Monforte, A.J.; Saladie, M.; Correa, R.X.; Garcia-Mas, J.; Martin-Hernandez, A.M. The complex resistance to cucumber mosaic cucumovirus (CMV) in the melon accession PI161375 is governed by one gene and at least two quantitative trait loci. Mol. Breed. 2014, 34, 351–362. [Google Scholar] [CrossRef]
- Saez, C.; Esteras, C.; Martinez, C.; Ferriol, M.; Dhillon, N.P.S.; Lopez, C.; Pico, B. Resistance to tomato leaf curl New Delhi virus in melon is controlled by a major QTL located in chromosome 11. Plant Cell Rep. 2017, 36, 1571–1584. [Google Scholar] [CrossRef]
- Perchepied, L.; Dogimont, C.; Pitrat, M. Strain-specific and recessive QTLs involved in the control of partial resistance to Fusarium oxysporum f. sp. melonis race 1.2 in a recombinant inbred line population of melon. Theor. Appl. Genet. 2005, 111, 65–74. [Google Scholar] [CrossRef]
- Díaz, A.; Zarouri, B.; Fergany, M.; Eduardo, I.; Alvarez, J.M.; Picó, B.; Monforte, A.J. Mapping and introgression of QTL involved in fruit shape transgressive segregation into ‘Piel de Sapo’ Melon (Cucumis melo L.). PLoS ONE 2014, 9, e104188. [Google Scholar]
- Oren, E.; Tzuri, G.; Dafna, A.; Meir, A.; Kumar, R.; Katzir, N.; Elkind, Y.; Freilich, S.; Schaffer, A.A.; Tadmor, Y.; et al. High-density NGS-based map construction and genetic dissection of fruit shape and rind netting in Cucumis melo. Theor. Appl. Genet. 2020, 133, 1927–1945. [Google Scholar] [CrossRef]
- Wallace, J.G.; Larsson, S.J.; Buckler, E.S. Entering the second century of maize quantitative genetics. Heredity 2014, 112, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Stitzer, M.C.; Ross-Ibarra, J. Maize domestication and gene interaction. New Phytol. 2018, 220, 395–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, A.L.; Briggs, W.H.; Rucker, J.; Baltazar, B.M.; Sanchez-Gonzalez, J.D.; Feng, P.; Buckler, E.S.; Doebley, J. The genetic architecture of complex traits in teosinte (Zea mays ssp parviglumis): New evidence from association mapping. Genetics 2008, 180, 1221–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Studer, A.J.; Doebley, J.F. Do Large Effect QTL Fractionate? A Case Study at the Maize Domestication QTL teosinte branched1. Genetics 2011, 188, 673–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.G.; Liang, H.L.; Chen, G.P.; Li, F.F.; Wang, Y.S.; Liao, C.G.; Hui, Z.L. The bHLH transcription factor SlPRE2 regulates tomato fruit development and modulates plant response to gibberellin. Plant Cell Rep. 2019, 38, 1053–1064. [Google Scholar] [CrossRef]
- Mu, Q.; Huang, Z.J.; Chakrabarti, M.; Illa-Berenguer, E.; Liu, X.X.; Wang, Y.P.; Ramos, A.; van der Knaap, E. Fruit weight is controlled by Cell Size Regulator encoding a novel protein that is expressed in maturing tomato fruits. PLoS Genet. 2017, 13, e1006930. [Google Scholar] [CrossRef]
- Czerednik, A.; Busscher, M.; Angenent, G.C.; de Maagd, R.A. The cell size distribution of tomato fruit can be changed by overexpression of CDKA1. Plant Biotech. J. 2015, 13, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Czerednik, A.; Busscher, M.; Bielen, B.A.; Wolters-Arts, M.; de Maagd, R.A.; Angenent, G.C. Regulation of tomato fruit pericarp development by an interplay between CDKB and CDKA1 cell cycle genes. J. Exp. Bot. 2012, 63, 2605–2617. [Google Scholar] [CrossRef] [Green Version]
- Doebley, J.; Stec, A.; Gustus, C. Teosinte branched1 and the origin of maize—Evidence for epistasis and the evolution of dominance. Genetics 1995, 141, 333–346. [Google Scholar]
- von Korff, M.; Leon, J.; Pillen, K. Detection of epistatic interactions between exotic alleles introgressed from wild barley (H. vulgare ssp. spontaneum). Theor. Appl. Genet. 2010, 8, 1455–1464. [Google Scholar] [CrossRef]
- Azhacuvel, P.; Vidya-Saraswathi, D.; Komatsuda, T. High-resolution linkage mapping for the non-brittle rachis locus btr1 in cultivated x wild barley (Hordeum vulgare). Plant Sci. 2006, 170, 1087–1094. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, S.; Salomon, B.; Komatsuda, T. The domestication syndrome genes responsible for the major changes in plant form in the Triticeae crops. Plant Cell Phys. 2011, 52, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monforte, A.J.; Friedman, E.; Zamir, D.; Tanksley, S.D. Comparison of a set of allelic QTL-NILs for chromosome 4 of tomato: Deductions about natural variation and implications for germplasm utilization. Theor. Appl. Genet. 2001, 102, 572–590. [Google Scholar] [CrossRef]
- Gur, A.; Zamir, D. Unused natural variation can lift yield barriers in plant breeding. PLoS Biol. 2004, 2, 1610–1615. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.J.; McCouch, S.R. Leveraging natural diversity: Back through the bottleneck. Curr. Opin. Plant Biol. 2008, 11, 193–200. [Google Scholar] [CrossRef]
- Doust, A.N.; Lukens, L.; Olsen, K.M.; Mauro-Herrera, M.; Meyer, A.; Rogers, K. Beyond the single gene: How epistasis and gene-by-environment effects influence crop domestication. Proc. Natl. Acad. Sci. USA 2014, 111, 6178–6183. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Piel de Sapo | 14M2-F2 | |||
|---|---|---|---|---|
| Trait | Mean ± sd | CV | Mean ± sd | CV |
| Fruit Shape (FS) | 1.30 ± 0.02 | 0.015 | 1.30 ± 0.15 | 0.115 |
| Pericarp Area (PA) | 0.68 ± 0.05 | 0.074 | 0.65 ± 0.082 | 0.126 |
| Pericarp Thickness (PT) | 0.40 ± 0.04 | 0.100 | 0.41 ± 0.04 | 0.098 |
| Fruit Weight (FW) | 2485.00 ± 356.38 | 0.143 | 1273.9 ± 402.9 | 0.316 |
| Trait | Marker | LOD Threshold | LOD | Genetic Position (cM) | Physical Position (bp) | Confidence Interval | a | d | d/[a] | R2 |
|---|---|---|---|---|---|---|---|---|---|---|
| Fruit Shape | Sca21-3371186 | 2.1 | 3.29 | 3.12 | 32,568,674 | 0.0–13.5 | −0.07 | 0.00 | 0.05 | 0.11 |
| Fruit Weight | Sca21-2801185 | 2.2 | 2.77 | 18.04 | 33,138,675 | 5.98–20.07 | −191.94 | −21.82 | −0.11 | 0.13 |
| Fruit Weight | Pericarp Area | Pericarp Thickness | ||||
| mean | 1036.37 | 0.63 | 0.36 | |||
| TRI_6-3-F2 | sd | 424.09 | 0.03 | 0.02 | ||
| PS | mean | 1557.61 | 0.68 | 0.39 | ||
| sd | 322.11 | 0.02 | 0.02 | |||
| SNP | Chromosome | Genotype | ||||
| a38f04 | 6 | PS/PS | 1426.21 | * | 0.64 | 0.37 |
| TRI/TRI | 828.91 | 0.63 | 0.36 |
| Family | Chr6 | Chr11 | Pericarp Area | Dunnet | 18M56 vs. 18M57 | Pericarp Thickness | Dunnet | 18M56 vs. 18M57 | Fruit Weight | Dunnet | 18M56 vs. 18M57 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TRI_6-3 | TRI | TRI | 0.58 ± 0.02 | * | 0.33 ± 0.01 | * | 295.7 ± 33.55 | * | |||
| 18M55 | PS | PS | 0.66 ± 0.05 | 0.38 ± 0.01 | 953.67 ± 259.2 | * | |||||
| 18M56 | TRI | PS | 0.61 ± 0.04 | * | * | 0.34 ± 0.01 | * | * | 499.48 ± 55.09 | * | n.s |
| 18M57 | TRI | TRI | 0.55 ± 0.04 | * | * | 0.30 ± 0.01 | * | * | 415.03 ± 105.78 | * | n.s |
| 18M60 | PS | TRI | 0.66 ± 0.03 | 0.38 ± 0.01 | 1252.75 ± 118.27 | * | |||||
| PS | PS | PS | 0.68 ± 0.01 | 0.39 ± 0.01 | 1578.25 ± 150.36 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riahi, C.; Reig-Valiente, J.L.; Picó, B.; Díaz, A.; Gonzalo, M.J.; Monforte, A.J. Evidence of the Role of QTL Epistatic Interactions in the Increase of Melon Fruit Flesh Content during Domestication. Agronomy 2020, 10, 1064. https://doi.org/10.3390/agronomy10081064
Riahi C, Reig-Valiente JL, Picó B, Díaz A, Gonzalo MJ, Monforte AJ. Evidence of the Role of QTL Epistatic Interactions in the Increase of Melon Fruit Flesh Content during Domestication. Agronomy. 2020; 10(8):1064. https://doi.org/10.3390/agronomy10081064
Chicago/Turabian StyleRiahi, Chaymaa, Juan Luis Reig-Valiente, Belén Picó, Aurora Díaz, Maria José Gonzalo, and Antonio José Monforte. 2020. "Evidence of the Role of QTL Epistatic Interactions in the Increase of Melon Fruit Flesh Content during Domestication" Agronomy 10, no. 8: 1064. https://doi.org/10.3390/agronomy10081064