Mild Water Stress Makes Apple Buds More Likely to Flower and More Responsive to Artificial Forcing— Impacts of an Unusually Warm and Dry Summer in Germany

 , ,
, ,
Abstract
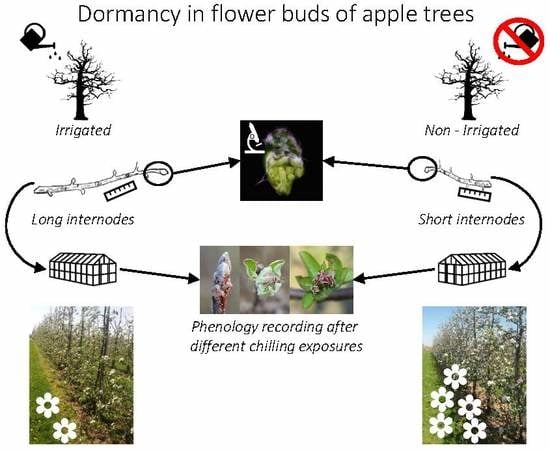
:
1. Introduction
2. Materials and Methods
2.1. Site and Plant Material
2.2. Morphological and Developmental Observations on Shoots and Apical Buds in the Field
2.3. Phenology Observations under Greenhouse and Field Conditions
2.4. Proportion of the Buds that Generated a Cyme
2.5. Chill and Heat Quantification and Chill Requirement Estimation
2.6. Starch Evaluation on King Flower Primordia
2.7. Statistical Analysis
3. Results
3.1. Morphological Observations on Shoots and Microscopic Monitoring of Apical Buds
3.2. Bud Development on One-Year-Old Shoots under Greenhouse Conditions
3.3. Phenology Progression and Flowering Rates under Field Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deutscher Wetterdienst. Zeitreihen und Trends. Available online: https://www.dwd.de/DE/leistungen/zeitreihen/zeitreihen.html#buehneTop (accessed on 17 November 2019).
- Jacob, D.; Kottmeier, C.; Petersen, J.; Rechid, D.; Teichmann, C. Regionale klimamodellierung. In Klimawandel in Deutschland: Entwicklung, Folgen, Risiken und Perspektiven; Brasseur, G.P., Jacob, D., Schuck-Zöller, S., Eds.; Springer: Berlin, Germany, 2017; pp. 27–35. [Google Scholar] [CrossRef]
- Van Oldenborgh, G.J.; Collins, M.; Arblaster, J.; Christensen, J.; Marotzke, J.; Power, S.; Rummukainen, M.; Zhou, T. Annex I: Atlas of global and regional climate projections. In Climate Change 2013: The physical science basis. Contribution of working group I to the fifth assessment report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Wilkie, J.D.; Sedgley, M.; Olesen, T. Regulation of floral initiation in horticultural trees. J. Exp. Bot. 2008, 59, 3215–3228. [Google Scholar] [CrossRef] [Green Version]
- Chai, Q.; Gan, Y.; Zhao, C.; Xu, H.; Waskom, R.M.; Niu, Y.; Siddique, K.H.M. Regulated deficit irrigation for crop production under drought stress. A review. Agron. Sustain. Dev. 2015, 36, 3. [Google Scholar] [CrossRef] [Green Version]
- Behboudian, M.H.; Mills, T.M. Deficit irrigation in deciduous orchards. Hortic. Rev. 1997, 21, 105–131. [Google Scholar] [CrossRef]
- Ebel, R.C.; Proebsting, E.L.; Patterson, M.E. Regulated deficit irrigation may alter apple maturity, quality, and storage life. HortScience 1993, 28, 141–143. [Google Scholar] [CrossRef] [Green Version]
- Ebel, R.C.; Proebsting, E.L.; Evans, R.G. Deficit irrigation to control vegetative growth in apple and monitoring fruit growth to schedule irrigation. HortScience 1995, 30, 1229–1232. [Google Scholar] [CrossRef] [Green Version]
- Berman, M.E.; DeJong, T.M. Water stress and crop load effects on fruit fresh and dry weights in peach (Prunus persica). Tree Physiol. 1996, 16, 859–864. [Google Scholar] [CrossRef]
- Bolat, I.; Dikilitas, M.; Ercisli, S.; Ikinci, A.; Tonkaz, T. The effect of water stress on some morphological, physiological, and biochemical characteristics and bud success on apple and quince rootstocks. Sci. World J. 2014, 1–8. [Google Scholar] [CrossRef]
- Naor, A.; Naschitz, S.; Peres, M.; Gal, Y. Responses of apple fruit size to tree water status and crop load. Tree Physiol. 2008, 28, 1255–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Failla, O.; Zocchi, G.; Treccani, C.; Cocucci, S. Growth, development and mineral-content of apple fruit in different water status conditions. J. Hortic. Sci. 1992, 67, 265–271. [Google Scholar] [CrossRef]
- Kramer, P.J. 2-Cell Water Relations. In Water Relations of Plants; Kramer, P.J., Ed.; Academic Press: New York, NY, USA, 1983; pp. 23–56. [Google Scholar] [CrossRef]
- Naschitz, S.; Naor, A.; Genish, S.; Wolf, S.; Goldschmidt, E.E. Internal management of non-structural carbohydrate resources in apple leaves and branch wood under a broad range of sink and source manipulations. Tree Physiol. 2010, 30, 715–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.Y.; Pallas, B.; Martinez, S.; Wang, Y.K.; Costes, E. Neoformation and summer arrest are common sources of tree plasticity in response to water stress of apple cultivars. Ann. Bot. 2019, 123, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Sobeih, W.Y.; Dodd, I.C.; Bacon, M.A.; Grierson, D.; Davies, W.J. Long-distance signals regulating stomatal conductance and leaf growth in tomato (Lycopersicon esculentum) plants subjected to partial root-zone drying. J. Exp. Bot. 2004, 55, 2353–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohde, A.; Bhalerao, R.P. Plant dormancy in the perennial context. Trends Plant Sci. 2007, 12, 217–223. [Google Scholar] [CrossRef]
- Cooke, J.E.K.; Eriksson, M.E.; Junttila, O. The dynamic nature of bud dormancy in trees: Environmental control and molecular mechanisms. Plant Cell Environ. 2012, 35, 1707–1728. [Google Scholar] [CrossRef]
- Lang, G.A. Dormancy-a new universal terminology. HortScience 1987, 22, 817–820. [Google Scholar]
- Faust, M.; Erez, A.; Rowland, L.J.; Wang, S.Y.; Norman, H.A. Bud dormancy in perennial fruit trees: Physiological basis for dormancy induction, maintenance, and release. HortScience 1997, 32, 623–629. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E. Climate change impacts on winter chill for temperate fruit and nut production: A review. Sci. Hortic. 2012, 144, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Campoy, J.A.; Ruiz, D.; Nortes, M.D.; Egea, J. Temperature efficiency for dormancy release in apricot varies when applied at different amounts of chill accumulation. Plant Biol. 2013, 15, 28–35. [Google Scholar] [CrossRef]
- Benmoussa, H.; Ghrab, M.; Ben Mimoun, M.; Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agric. For. Meteorol. 2017, 239, 34–46. [Google Scholar] [CrossRef]
- Guo, L.; Dai, J.H.; Wang, M.C.; Xu, J.C.; Luedeling, E. Responses of spring phenology in temperate zone trees to climate warming: A case study of apricot flowering in China. Agric. For. Meteorol. 2015, 201, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Alburquerque, N.; Garcia-Montiel, F.; Carrillo, A.; Burgos, L. Chilling and heat requirements of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements. Environ. Exp. Bot. 2008, 64, 162–170. [Google Scholar] [CrossRef]
- Harrington, C.A.; Gould, P.J.; St Clair, J.B. Modeling the effects of winter environment on dormancy release of Douglas-fir. For. Ecol. Manage. 2010, 259, 798–808. [Google Scholar] [CrossRef]
- Pope, K.S.; Da Silva, D.; Brown, P.H.; DeJong, T.M. A biologically based approach to modeling spring phenology in temperate deciduous trees. Agric. For. Meteorol. 2014, 198, 15–23. [Google Scholar] [CrossRef]
- Fadón, E.; Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environ. Exp. Bot. 2018, 152, 28–36. [Google Scholar] [CrossRef]
- Luedeling, E.; Kunz, A.; Blanke, M.M. Identification of chilling and heat requirements of cherry trees-a statistical approach. Int. J. Biometeorol. 2013, 57, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Kurokura, T.; Mimida, N.; Battey, N.H.; Hytönen, T. The regulation of seasonal flowering in the Rosaceae. J. Exp. Bot. 2013, 64, 4131–4141. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, E.; Baird, G.; Farias, D.; Oyanedel, E.; Olaeta, J.A.; Brown, P.; Zwieniecki, M.; Tixier, A.; Saa, S. Fruit load in almond spurs define starch and total soluble carbohydrate concentration and therefore their survival and bloom probabilities in the next season. Sci. Hortic. 2018, 237, 269–276. [Google Scholar] [CrossRef]
- Dietze, M.C.; Sala, A.; Carbone, M.S.; Czimczik, C.I.; Mantooth, J.A.; Richardson, A.D.; Vargas, R. Nonstructural carbon in woody plants. In Annual Review of Plant Biology; Merchant, S.S., Ed.; Annual Reviews: Palo Alto, CA, USA; Volume 65, 2014; pp. 667–687. [Google Scholar]
- Rodrigo, J.; Hormaza, J.I.; Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiol. Plant. 2000, 108, 35–41. [Google Scholar] [CrossRef]
- Fernandez, E.; Cuneo, I.F.; Luedeling, E.; Alvarado, L.; Farias, D.; Saa, S. Starch and hexoses concentrations as physiological markers in dormancy progression of sweet cherry twigs. Trees Struct. Funct. 2019, 33, 1187–1201. [Google Scholar] [CrossRef]
- Hedhly, A.; Vogler, H.; Schmid, M.; Pazmino, D.; Gagliardini, V.; Santelia, D.; Grossniklaus, U. Starch turnover and metabolism during flower and early embryo development. Plant Physiol. 2016, 172, 2388–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimienta, E.; Polito, V.S. Ovule abortion in `Nonpareil’ almond (Prunus dulcis [Mill.] D. A. Webb). Am. J. Bot. 1982, 69, 913–920. [Google Scholar] [CrossRef]
- Rodrigo, J.; Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sex. Plant Reprod. 1998, 11, 86–93. [Google Scholar] [CrossRef]
- Alcaraz, M.L.; Hormaza, J.I.; Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS ONE 2013, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Herrero, M.; Dickinson, H.G. Pollen-pistil incompatibility in Petunia hybrida: Changes in the pistil following compatible and incompatible intraspecific crosses. J. Cell Sci. 1979, 36, 1–18. [Google Scholar]
- Rodrigo, J.; Herrero, M.; Hormaza, J.I. Pistil traits and flower fate in apricot (Prunus armeniaca). Ann. Appl. Biol. 2009, 154, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, R.; García-Luis, A.; Monerri, C.; Guardiola, J.L. Carbohydrate availability in relation to fruitlet abscission in Citrus. Ann. Bot. 2001, 87, 805–812. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, D.J.; Tadeo, F.R.; Primo-Millo, E.; Talon, M. Fruit set dependence on carbohydrate availability in citrus trees. Tree Physiol. 2003, 23, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. Global warming and sexual plant reproduction. Trends Plant. Sci. 2009, 14, 30–36. [Google Scholar] [CrossRef]
- Statistisches Bundesamt (Destatis). Areas and quantities harvested (production for the market). Available online: https://www.destatis.de/EN/Themes/Economic-Sectors-Enterprises/Agriculture-Forestry-Fisheries/Fruit-Vegetables-Horticulture/Tables/2-4-areas-quantities-harvested.html (accessed on 8 November 2019).
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Koppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration. Guidelines for computing crop water requirements. In Irrigation and Drainage; Paper No. 56; FAO: Rome, Italy, 1998; p. 300. [Google Scholar]
- Meier, U. Growth Stages of Mono-and Dicotyledonous Plants, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry, Oxford: Berlin, Germany, 2001. [Google Scholar]
- Martinez, R.; Legua, P.; Martinez-Nicolas, J.J.; Melgarejo, P. Phenological growth stages of “Pero de Cehegin” (Malus domestica Borkh): Codification and description according to the BBCH scale. Sci. Hortic. 2019, 246, 826–834. [Google Scholar] [CrossRef]
- Luedeling, E. chillR: Statistical methods for phenology analysis in temperate fruit trees, R package version 0.70.21. 2019.
- Erez, A.; Fishman, S.; Linsley-Noakes, G.C.; Allan, P. The dynamic model for rest completion in peach buds. Acta Hort. 1990, 276, 165–174. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature-dependence of dormancy breaking in plants-computer-simulation of processes studied under controlled temperatures. J. Theor. Biol. 1987, 126, 309–321. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature-dependence of dormancy breaking in plants-mathematical-analysis of a 2-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Zhang, J.L.; Taylor, C. The Dynamic Model provides the best description of the chill process on ‘Sirora’ pistachio trees in Australia. HortScience 2011, 46, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E.; Kunz, A.; Blanke, M. Mehr Chilling für Obstbäume in wärmeren Wintern? Erwerbs Obstbau 2011, 53, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.L.; Richardson, E.A.; Kesner, C.D. Validation of chill unit and flower bud phenology models for ‘montmorency’ sour cherry. Acta Horticulturae ISHS 1986, 184, 71–78. [Google Scholar] [CrossRef]
- Herr, J.M. A new clearing-squash technique for the study of ovule development in Angiosperms. Am. J. Bot. 1971, 58, 785–790. [Google Scholar] [CrossRef]
- Herr, J.M. Applications of a new clearing technique for the investigation of vascular plant morphology. J. Elisha Mitchell Sci. Soc. 1972, 88, 137–143. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, 3.5.3; R Core Team R: Vienna, Austria, 2019.
- Hollander, M.A.; Wolfe, D.; Chicken, E. Ranked Set Sampling. In Nonparametric Statistical Methods, 3rd ed.; Hollander, M.A., Wolfe, D., Chicken, E., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Bonferroni, C. Teoria statistica delle classi e calcolo delle probabilita’. Pubbl. R Ist. Sup. Sci. Econ. Commer. Fir. 1936, 8, 3–62. [Google Scholar]
- Berhe, D.F.; Taxis, K.; Haaijer-Ruskamp, F.M.; Mulugeta, A.; Mengistu, Y.T.; Burgerhof, J.G.M.; Mol, P.G.M. Impact of adverse drug events and treatment satisfaction on patient adherence with antihypertensive medication-a study in ambulatory patients. Br. J. Clin. Pharmacol. 2017, 83, 2107–2117. [Google Scholar] [CrossRef] [PubMed]
- Tukey, J.W. Exploratory Data Analysis; Limited Preliminary Ed.; Addison-Wesley Publishing Company: Boston, MA, USA, 1970. [Google Scholar]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones enhanced drought tolerance in plants: a coping strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.; Johnston, R.; Seleznyova, A. A morphological and quantitative characterization of early floral development in apple (Malus x domestica Borkh.). Ann. Bot. 2003, 92, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadon, E.; Rodrigo, J.; Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees-Struct. Funct. 2018, 32, 1583–1594. [Google Scholar] [CrossRef]
- Felker, F.C.; Robitaille, H.A.; Hess, F.D. Morphological and ultrastructural development and starch accumulation during chilling of sour cherry flower buds. Am. J. Bot. 1983, 70, 376–386. [Google Scholar] [CrossRef]
- Fadon, E.; Herrero, M.; Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Front. Plant Sci. 2018, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Fadon, E.; Herrero, M.; Rodrigo, J. Anther and pollen development in sweet cherry (Prunus avium L.) in relation to winter dormancy. Protoplasma 2019, 256, 733–744. [Google Scholar] [CrossRef]
- Nakajima, Y.; Susanto, S.; Hasegawa, K. Influence of water-stress in autumn on flower induction and fruiting in young pomelo trees (Citrus grandis (l) Osbeck). J. Jpn. Soc. Hortic. Sci. 1993, 62, 15–20. [Google Scholar] [CrossRef]
- Sanyal, D.; Bangerth, F. Stress induced ethylene evolution and its possible relationship to auxin-transport, cytokinin levels, and flower bud induction in shoots of apple seedlings and bearing apple trees. Plant. Growth Regul. 1998, 24, 127–134. [Google Scholar] [CrossRef]
- Beauvieux, R.; Wenden, B.; Dirlewanger, E. Bud dormancy in perennial fruit tree species: A pivotal role for oxidative cues. Front. Plant Sci. 2018, 9, 13. [Google Scholar] [CrossRef]
- Erez, A. Bud dormancy phenomenon, problems and solutions in the tropics and subtropics. In Temperate Fruit Crops in Warm Climates; Erez, A., Ed.; Springer: Dordrecht, The Netherlands, 2000; pp. 17–48. [Google Scholar] [CrossRef]
- Shaltout, A.D.; Unrath, C.R. Rest completion prediction model for starkrimson delicious apples. J. Am. Soc. Hort. Sci. 1983, 108, 957–961. [Google Scholar]
- Luedeling, E.; Brown, P.H. A global analysis of the comparability of winter chill models for fruit and nut trees. Int. J. Biometeorol. 2011, 55, 411–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luedeling, E.; Girvetz, E.H.; Semenov, M.A.; Brown, P.H. Climate change affects winter chill for temperate fruit and nut trees. PLoS ONE 2011, 6, 13. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date of Sampling | Chill Received in the Field (in CP) |
|---|---|
| 7th December 2018 | 24.9 |
| 14th December 2018 | 29.7 |
| 21st December 2018 | 34.4 |
| 28th December 2018 | 39.4 |
| 4th January 2019 | 44.9 |
| 11th January 2019 | 49.6 |
| 18th January 2019 | 55.5 |
| 25th January 2019 | 56.3 |
| 2nd February 2019 | 63.7 |
| 8th February 2019 | 68.6 |
| Transition Point (from Stage x to Stage y) | Difference in Threshold Estimates (Non-Irrigated – Irrigated) | Confidence Interval for the Difference (95%) |
|---|---|---|
| BBCH 0 to BBCH 51-52 | −0.62 | −0.90 to −0.34 *** |
| BBCH 51-52 to BBCH 53-54 | −0.59 | −0.88 to −0.30 *** |
| BBCH 53-54 to BBCH 55-56 | −0.27 | −0.57 to −0.05 n.s. |
| BBCH 55-56 to BBCH 57-59 | −0.52 | −0.91 to −0.13 *** |
| BBCH 57-59 to BBCH 60 | −0.09 | −0.61 to −0.42 n.s. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez, E.; Luedeling, E.; Behrend, D.; Van de Vliet, S.; Kunz, A.; Fadón, E. Mild Water Stress Makes Apple Buds More Likely to Flower and More Responsive to Artificial Forcing— Impacts of an Unusually Warm and Dry Summer in Germany. Agronomy 2020, 10, 274. https://doi.org/10.3390/agronomy10020274
Fernandez E, Luedeling E, Behrend D, Van de Vliet S, Kunz A, Fadón E. Mild Water Stress Makes Apple Buds More Likely to Flower and More Responsive to Artificial Forcing— Impacts of an Unusually Warm and Dry Summer in Germany. Agronomy. 2020; 10(2):274. https://doi.org/10.3390/agronomy10020274
Chicago/Turabian StyleFernandez, Eduardo, Eike Luedeling, Dominik Behrend, Stijn Van de Vliet, Achim Kunz, and Erica Fadón. 2020. "Mild Water Stress Makes Apple Buds More Likely to Flower and More Responsive to Artificial Forcing— Impacts of an Unusually Warm and Dry Summer in Germany" Agronomy 10, no. 2: 274. https://doi.org/10.3390/agronomy10020274