
Dietary Serine Supplementation Improves Growth Performance, Intramuscular Fat Content, and Composition of Gut Microbes and Metabolites in Growing–Finishing Pigs

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sample Collection
2.3. Biochemical Parameters
2.4. Meat Quality
2.5. Fecal Microbiota Profiling
2.6. Determination of Fecal Metabolites
2.7. Statistical Analysis
3. Results
3.1. Growth Performance and Serum Biochemical Parameters
3.2. Meat Quality
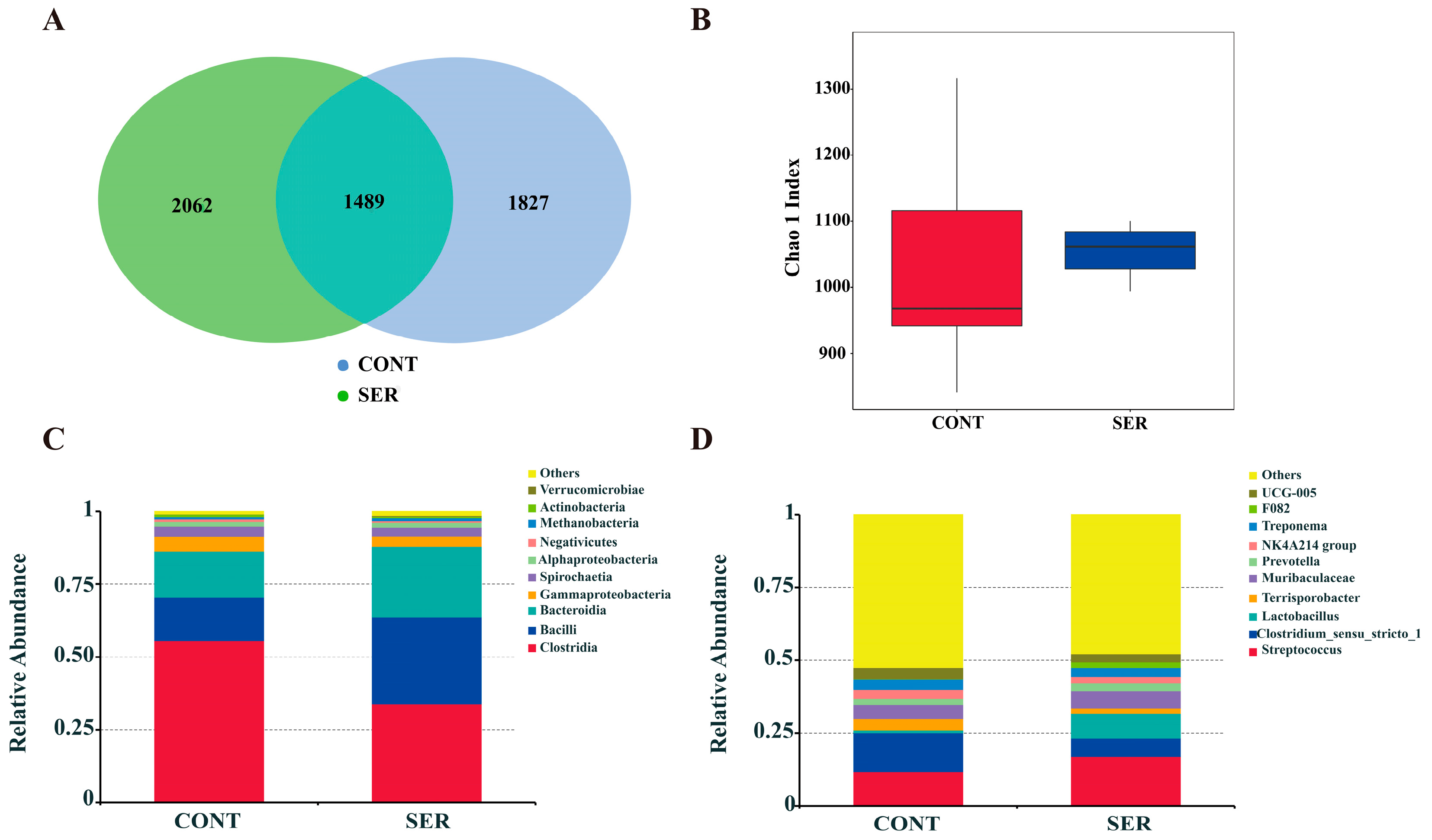
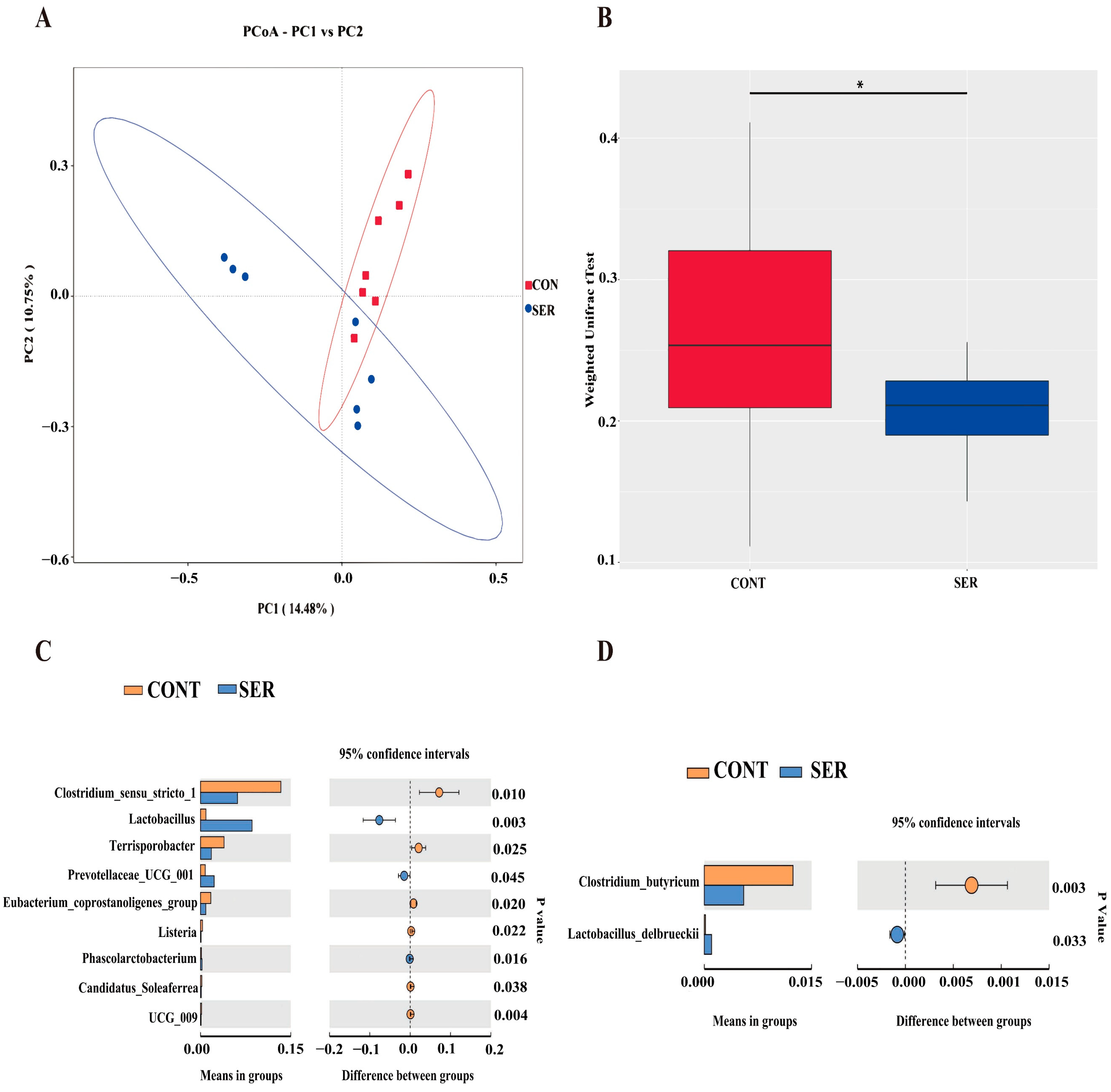
3.3. Fecal Microbiota Composition
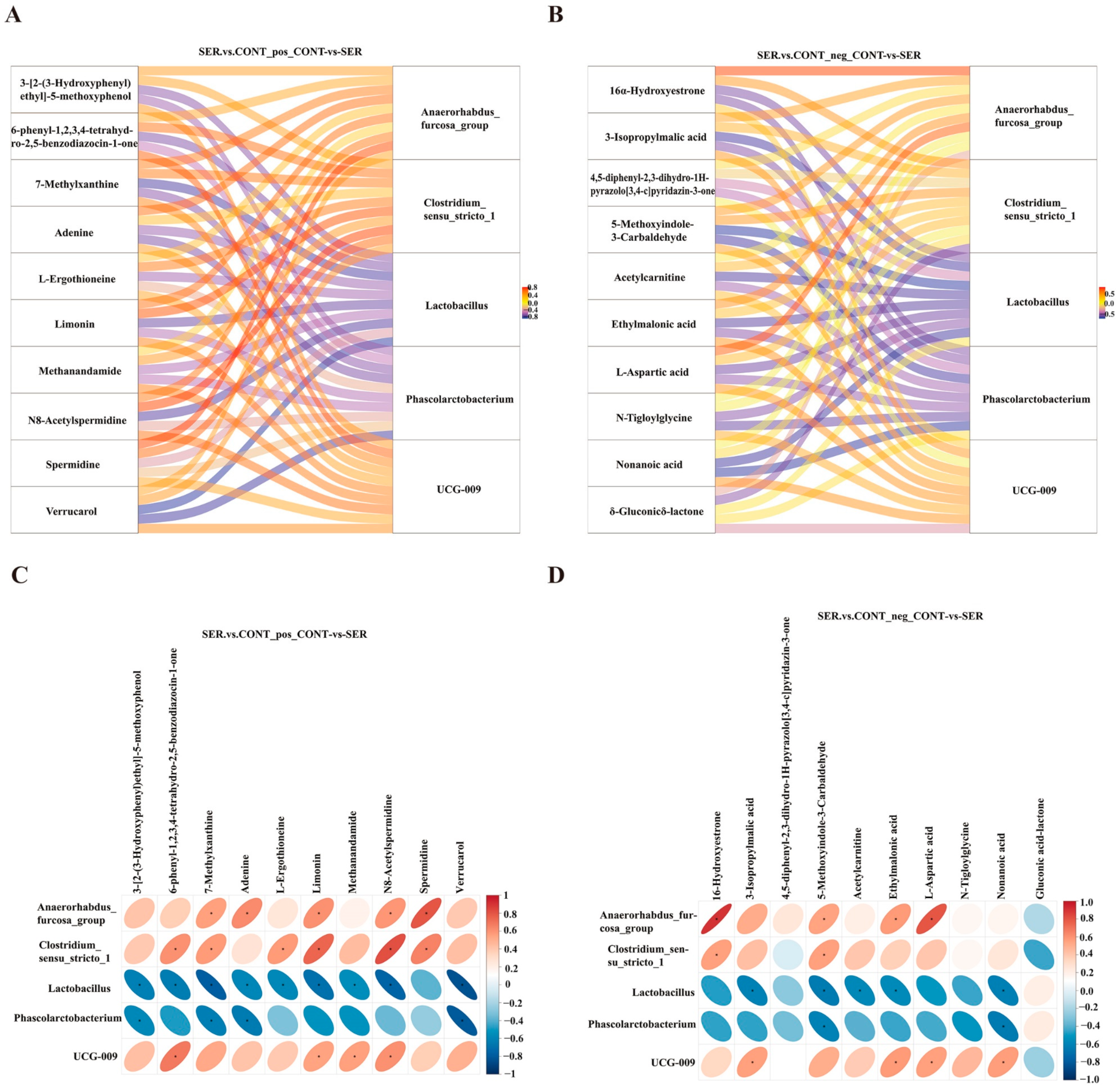
3.4. Fecal Metabolites
3.5. Correlation between Microbiota and Metabolites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, B.; Chen, Y.; Li, Q.; Zhong, Z.; Tan, Y.; Zhang, S.; Zhu, L. Effects of Breed and Gender Effect on Pork Quality Traits. Southwest. China J. Agric. Sci. 2019, 32, 2222–2225. [Google Scholar]
- Wang, Y.; Thakali, K.; Morse, P.; Shelby, S.; Chen, J.; Apple, J.; Huang, Y. Comparison of Growth Performance and Meat Quality Traits of Commercial Cross-Bred Pigs versus the Large Black Pig Breed. Animals 2021, 11, 200. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.L.; Li, D.Y.; Leng, D.; Kui, H.; Bai, X.; Wang, T. Gut microbiota and meat quality. Front. Microbiol. 2022, 13, 951726. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Zeng, D.; Cheng, L.; Mao, X.; Yu, J.; Yu, B.; Chen, D. Dietary beta-glucan supplementation improves growth performance, carcass traits and meat quality of finishing pigs. Anim. Nutr. 2019, 5, 380–385. [Google Scholar] [CrossRef]
- Tian, Z.; Deng, D.; Cui, Y.; Chen, W.; Yu, M.; Ma, X. Diet supplemented with fermented okara improved growth performance, meat quality, and amino acid profiles in growing pigs. Food Sci. Nutr. 2020, 8, 5650–5659. [Google Scholar] [CrossRef]
- Ma, X.; Yu, M.; Liu, Z.; Deng, D.; Cui, Y.; Tian, Z.; Wang, G. Effect of amino acids and their derivatives on meat quality of finishing pigs. J. Food Sci. Technol. 2020, 57, 404–412. [Google Scholar] [CrossRef]
- Hu, C.J.; Jiang, Q.Y.; Zhang, T.; Yin, Y.L.; Li, F.N.; Deng, J.P.; Wu, G.Y.; Kong, X.F. Dietary supplementation with arginine and glutamic acid modifies growth performance, carcass traits, and meat quality in growing-finishing pigs. J. Anim. Sci. 2017, 95, 2680–2689. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Long, J.; Zhou, X.; Liu, Y.; Li, T.; Wu, X. Serine is required for the maintenance of redox balance and proliferation in the intestine under oxidative stress. FASEB J. 2020, 34, 4702–4717. [Google Scholar] [CrossRef]
- Lionaki, E.; Gkikas, I.; Daskalaki, I.; Ioannidi, M.-K.; Klapa, M.I.; Tavernarakis, N. Mitochondrial protein import determines lifespan through metabolic reprogramming and de novo serine biosynthesis. Nat. Commun. 2022, 13, 651. [Google Scholar] [CrossRef]
- Waterhouse, H.N.; Scott, H.M. Effect of Different Proteins and Protein Levels on the Glycine Need of the Chick Fed Purified Diets. Poult. Sci. 1961, 40, 1160–1165. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, Y.; Zhang, L.; Kong, X.; Li, F. Serine-to-glycine ratios in low-protein diets regulate intramuscular fat by affecting lipid metabolism and myofiber type transition in the skeletal muscle of growing-finishing pigs. Anim. Nutr. 2021, 7, 384–392. [Google Scholar] [CrossRef]
- He, L.; Liu, Y.; Liu, D.; Feng, Y.; Yin, J.; Zhou, X. Exogenous and Endogenous Serine Deficiency Exacerbates Hepatic Lipid Accumulation. Oxid. Med. Cell. Longev. 2021, 2021, 4232704. [Google Scholar] [CrossRef]
- Ma, J.; Duan, Y.; Li, R.; Liang, X.; Li, T.; Huang, X.; Yin, Y.; Yin, J. Gut microbial profiles and the role in lipid metabolism in Shaziling pigs. Anim. Nutr. 2022, 9, 345–356. [Google Scholar] [CrossRef]
- Zhao, G.; Xiang, Y.; Wang, X.; Dai, B.; Zhang, X.; Ma, L.; Yang, H.; Lyu, W.; Jia, Z. Exploring the Possible Link between the Gut Microbiome and Fat Deposition in Pigs. Oxidative Med. Cell. Longev. 2022, 2022, 1098892. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Tian, Y.; Zhou, F.; Ma, J.; Xia, S.; Yang, T.; Ma, L.; Zeng, Q.; Liu, G.; et al. Obese Ningxiang pig-derived microbiota rewires carnitine metabolism to promote muscle fatty acid deposition in lean DLY pigs. Innovation 2023, 4, 100486. [Google Scholar] [CrossRef] [PubMed]
- Nguyen Cong, O.; Bernard, T.; Pham Kim, D.; Do Duc, L.; Nassim, M.; Nguyen Thi, H.; Nguyen Hoang, T.; Georges, D.; Jerome, B.; Vu Dinh, T.; et al. Growth performance, carcass quality characteristics and colonic microbiota profiles in finishing pigs fed diets with different inclusion levels of rice distillers’ by-product. Anim. Sci. J. Nihon Chikusan Gakkaiho 2019, 90, 948–960. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hua, R.; Zhang, B.; Zhang, X.; Yang, H.; Zhou, X. Serine Alleviates Dextran Sulfate Sodium-Induced Colitis and Regulates the Gut Microbiota in Mice. Front. Microbiol. 2018, 9, 3062. [Google Scholar] [CrossRef]
- Wang, Y.B.; Du, W.; Fu, A.K.; Zhang, X.P.; Huang, Y.; Lee, K.H.; Yu, K.; Li, W.F.; Li, Y.L. Intestinal microbiota and oral administration of Enterococcus faecium associated with the growth performance of new-born piglets. Benef. Microbes 2016, 7, 529–538. [Google Scholar] [CrossRef]
- Driessen, B.; Van Beirendonck, S.; Buyse, J. The Impact of Grouping on Skin Lesions and Meat Quality of Pig Carcasses. Animals 2020, 10, 544. [Google Scholar] [CrossRef] [PubMed]
- Henriquez-Rodriguez, E.; Tor, M.; Pena, R.N.; Estany, J. A polymorphism in the stearoyl-CoA desaturase gene promoter increases monounsaturated fatty acid content in dry-cured ham. Meat Sci. 2015, 106, 38–43. [Google Scholar] [CrossRef]
- Ogbuagu, N.E.; Ayo, J.O.; Aluwong, T.; Akor-Dewu, M.B. L-serine improves lipid profile, performance, carcass weight and intestinal parameters in feed restricted broiler chickens during the hot-dry season. Trop. Anim. Health Prod. 2022, 54, 12. [Google Scholar] [CrossRef]
- Lewis, C.A.; Parker, S.J.; Fiske, B.P.; McCloskey, D.; Gui, D.Y.; Green, C.R.; Vokes, N.I.; Feist, A.M.; Vander Heiden, M.G.; Metallo, C.M. Tracing Compartmentalized NADPH Metabolism in the Cytosol and Mitochondria of Mammalian Cells. Mol. Cell 2014, 55, 253–263. [Google Scholar] [CrossRef]
- Mao, Z.; Zhang, W. Role of mTOR in Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2018, 19, 2043. [Google Scholar] [CrossRef]
- Jiang, J.; Jin, F.; Lin, G.; Xiong, Y.L. Modulation of muscle antioxidant enzymes and fresh meat quality through feeding peptide-chelated trace minerals in swine production. Food Biosci. 2021, 42, 101191. [Google Scholar] [CrossRef]
- Zhou, X.; He, L.; Wu, C.; Zhang, Y.; Wu, X.; Yin, Y. Serine alleviates oxidative stress via supporting glutathione synthesis and methionine cycle in mice. Mol. Nutr. Food Res. 2017, 61, 1700262. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Xie, J.; Zhang, Y.; Wang, J.; Sun, X.; Zhang, H. Protective effects of probiotic Lactobacillus casei Zhang against endotoxin- and d-galactosamine-induced liver injury in rats via anti-oxidative and anti-inflammatory capacities. Int. Immunopharmacol. 2013, 15, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Fang, S.; Wei, H.; He, M.; Fu, H.; Xiong, X.; Zhou, Y.; Wu, J.; Gao, J.; Yang, H.; et al. Prevotella copri increases fat accumulation in pigs fed with formula diets. Microbiome 2021, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.; Bell, R.; Kiag, K.A.; Lee, S.H.; Soto, R.; Ghazaryan, A.; Buhrke, K.; Ekiz, H.A.; Ost, K.S.; Boudina, S.; et al. T cell-mediated regulation of the microbiota protects against obesity. Science 2019, 365, eaat9351. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Xin, Y.; Ma, Y.; Xu, X.; Zhao, S.; Cao, J. Screening of Microbes Associated With Swine Growth and Fat Deposition Traits Across the Intestinal Tract. Front. Microbiol. 2020, 11, 586776. [Google Scholar] [CrossRef] [PubMed]
- Izzo, L.T.; Trefely, S.; Demetriadou, C.; Drummond, J.M.; Mizukami, T.; Kuprasertkul, N.; Farria, A.T.; Nguyen, P.T.T.; Murali, N.; Reich, L.; et al. Acetylcarnitine shuttling links mitochondrial metabolism to histone acetylation and lipogenesis. Sci. Adv. 2023, 9, 20. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Sun, Y.; Chen, L.; Zhang, Y.; Wang, J.; Li, H.; Yan, X.; Xia, L.; Yao, G. Differences in meat quality between Angus cattle and Xinjiang brown cattle in association with gut microbiota and its lipid metabolism. Front. Microbiol. 2022, 13, 988984. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Y.J.; Zuo, S.X.; Peng, S.J.; Wang, Z.J.; Zhang, Y.J.; Luo, H.L. Untargeted and Targeted Metabolomics Profiling of Muscle Reveals Enhanced Meat Quality in Artificial Pasture Grazing Tan Lambs via Rescheduling the Rumen Bacterial Community. J. Agric. Food Chem. 2021, 69, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, P.; Habibi, M.; Roberts, K.; Sutton, J.; Shili, C.N.; Lin, D.; Pezeshki, A. Dietary Tryptophan Supplementation Alters Fat and Glucose Metabolism in a Low-Birthweight Piglet Model. Nutrients 2021, 13, 2561. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Goodarzi, P.; Pezeshki, A.; Hagen, D.E. RNA-seq reveals insights into molecular mechanisms of metabolic restoration via tryptophan supplementation in low birth weight piglet model. J. Anim. Sci. 2022, 100, skac156. [Google Scholar] [CrossRef] [PubMed]
- Zubiri-Gaitán, A.; Blasco, A.; Hernández, P. Plasma metabolomic profiling in two rabbit lines divergently selected for intramuscular fat content. Commun. Biol. 2023, 6, 893. [Google Scholar] [CrossRef]
- Niu, J.K.; Liu, X.; Xu, J.Y.; Li, F.; Wang, J.C.; Zhang, X.X.; Yang, X.; Wang, L.; Ma, S.; Li, D.F.; et al. Effects of Silage Diet on Meat Quality through Shaping Gut Microbiota in Finishing Pigs. Microbiol. Spectr. 2023, 11, 19. [Google Scholar] [CrossRef]
- Wu, C.; Lyu, W.; Hong, Q.; Zhang, X.; Yang, H.; Xiao, Y. Gut Microbiota Influence Lipid Metabolism of Skeletal Muscle in Pigs. Front. Nutr. 2021, 8, 675445. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Kwon, Y.; Kim, D.-K.; Jeon, J.; Jang, S.C.; Wang, T.; Ban, M.; Kim, M.-H.; Jeon, S.G.; Kim, M.-S.; et al. Gut microbe-derived extracellular vesicles induce insulin resistance, thereby impairing glucose metabolism in skeletal muscle. Sci. Rep. 2015, 5, 15878. [Google Scholar] [CrossRef]
- Virtue, A.T.; McCright, S.J.; Wright, J.M.; Jimenez, M.T.; Mowel, W.K.; Kotzin, J.J.; Joannas, L.; Basavappa, M.G.; Spencer, S.P.; Clark, M.L.; et al. The gut microbiota regulates white adipose tissue inflammation and obesity via a family of microRNAs. Sci. Transl. Med. 2019, 11, 13. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | 20~30 kg | 30~60 kg | 60~120 kg |
|---|---|---|---|
| Corn, % | 65.00 | 72.30 | 59.00 |
| Wheat bran, % | 6.00 | 3.00 | / |
| Rice, % | / | / | 20.00 |
| Soybean meal #, % | 25.00 | 19.50 | 14.00 |
| Soybean oil, % | / | 1.20 | 3.00 |
| Limestone, % | 1.00 | 0.80 | 0.80 |
| calcium hydrogen phosphate, % | 0.80 | 0.70 | 0.50 |
| NaCl, % | 0.40 | 0.45 | 0.30 |
| L-lysine hydrochloride, % | 0.30 | 0.60 | 0.25 |
| DL-methionine, % | 0.10 | 0.10 | / |
| L-threonine, % | 0.20 | 0.20 | / |
| L-tryptophan, % | / | 0.03 | / |
| Zeolite powder | 0.08 | / | 1.03 |
| Premix *, % | 1.12 | 1.12 | 1.12 |
| Total, % | 100 | 100 | 100 |
| Calculated nutrient level | |||
| DE (MJ/kg) | 13.86 | 14.28 | 14.03 |
| CP, % | 17.51 | 15.57 | 12.55 |
| Ca, % | 0.64 | 0.53 | 0.46 |
| Total P, % | 0.50 | 0.43 | 0.38 |
| P, % | 0.23 | 0.20 | 0.17 |
| Lysine, % | 1.03 | 1.12 | 0.71 |
| Methionine, % | 0.36 | 0.33 | 0.2 |
| Threonine, % | 0.73 | 0.65 | 0.37 |
| Tryptophan, % | 0.15 | 0.15 | 0.10 |
| CONT | SER | |
|---|---|---|
| Initial BW, kg | 20.95 ± 0.76 | 20.35 ± 0.38 |
| Final BW, kg | 116.8 ± 7.1 | 125.2 ± 4.9 |
| ADG, kg | 0.806 ± 0.059 a | 0.871 ± 0.026 b |
| ADFI, kg | 2.25 ± 0.16 | 2.29 ± 0.04 |
| F/G | 2.80 ± 0.05 | 2.63 ± 0.10 |
| CONT | SER | |
|---|---|---|
| GSH, ng/mL | 129.4 ± 17.4 a | 196.9 ± 19.1 b |
| SOD, ng/mL | 197.6 ± 29.3 a | 286.4 ± 17.2 b |
| MDA, nmol/mL | 4.06 ± 0.40 a | 1.91 ± 0.24 b |
| CONT | SER | |
|---|---|---|
| Color | ||
| L | 45.23 ± 1.56 | 44.11 ± 1.28 |
| a | 13.86 ± 0.33 | 14.48 ± 0.48 |
| b | 5.40 ± 0.57 | 5.08 ± 0.46 |
| pH45 min | 6.60 ± 0.09 | 6.42 ± 0.11 |
| pH24 h | 5.60 ± 0.07 | 5.53 ± 0.07 |
| Drip loss | 3.58 ± 0.50 a | 2.24 ± 0.33 b |
| Intramuscular fat | 2.26 ± 0.18 a | 2.94 ± 0.17 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; He, F.; Deng, Z.; Yin, J.; Guan, G.; Xie, Z.; Zhou, X. Dietary Serine Supplementation Improves Growth Performance, Intramuscular Fat Content, and Composition of Gut Microbes and Metabolites in Growing–Finishing Pigs. Agriculture 2024, 14, 349. https://doi.org/10.3390/agriculture14030349
Guo Y, He F, Deng Z, Yin J, Guan G, Xie Z, Zhou X. Dietary Serine Supplementation Improves Growth Performance, Intramuscular Fat Content, and Composition of Gut Microbes and Metabolites in Growing–Finishing Pigs. Agriculture. 2024; 14(3):349. https://doi.org/10.3390/agriculture14030349
Chicago/Turabian StyleGuo, Yiting, Fang He, Zhiying Deng, Jie Yin, Guiping Guan, Zhengjun Xie, and Xihong Zhou. 2024. "Dietary Serine Supplementation Improves Growth Performance, Intramuscular Fat Content, and Composition of Gut Microbes and Metabolites in Growing–Finishing Pigs" Agriculture 14, no. 3: 349. https://doi.org/10.3390/agriculture14030349