
The Profile of Bioactive Compounds in Two Tomato Lines (hp-2 and Atv_aft) When Grown under Low-Input Farming Conditions with or without Mulching and Intercropping

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
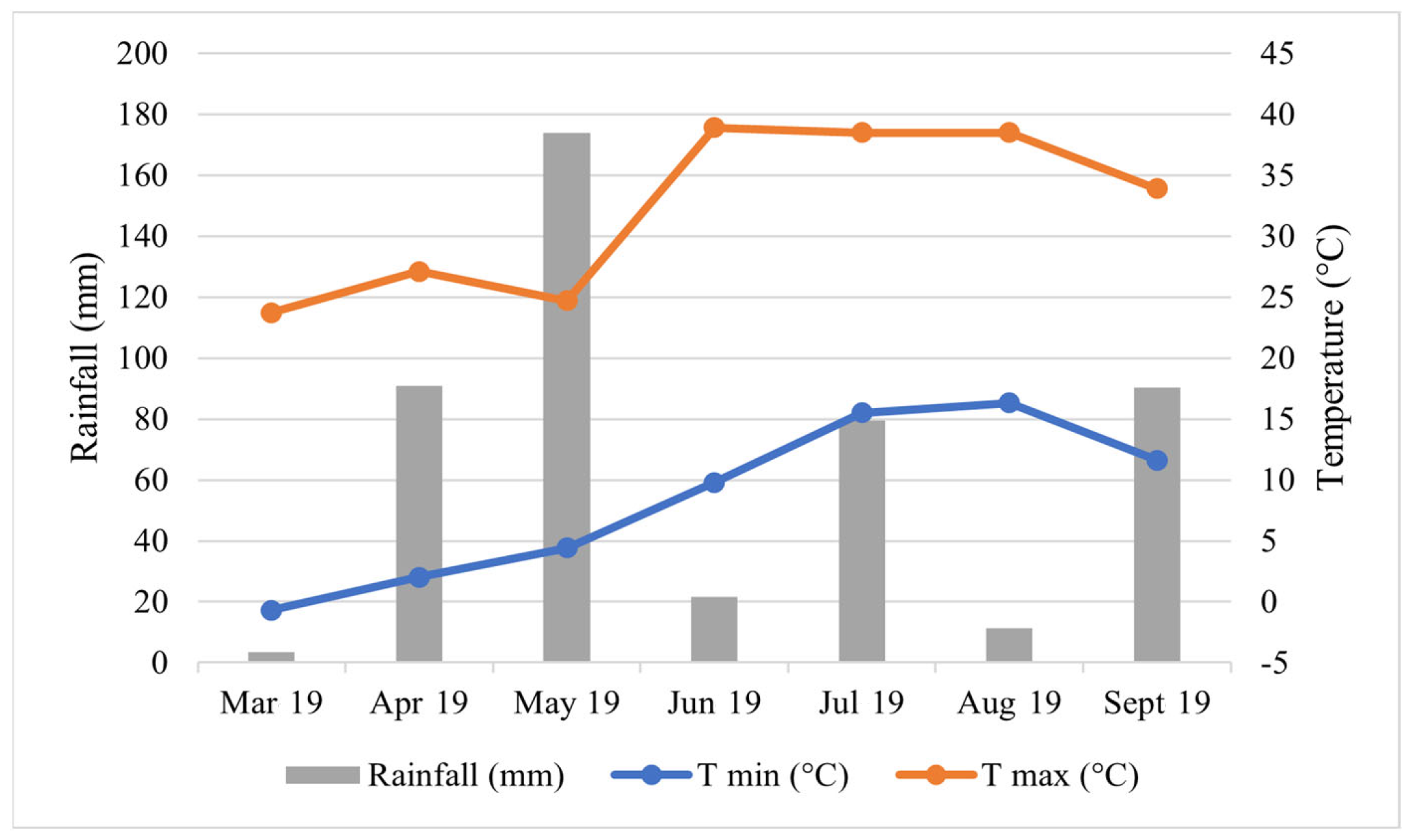
2.2. Field Experiment Description
2.3. Field Measurements
- (1)
- Fruit set (FS%). The ratio (%) between the number of fruits (TNF) and the number of flowers (NFL) observed before the first harvesting on the first and second truss of the tagged plants;
- (2)
- The total fruit number (TNF) and total weight of fruits (TWF) per plant. The weight was measured by a precision balance Kern (Stuttgart, Germany) mod. KB 10000-1N (d = 0.1 g);
- (3)
- The total soluble solid content (°Brix) was measured with an optical refractometer. The reading was made by placing one–two droplets of the juice extracted from five red ripe fruits randomly chosen in each single plot;
- (4)
- The pH value was measured on the juice (pHmeter Crison mod. 50 50 T, Barcelona, Spain).
2.4. Analysis of Bioactive Substances
2.5. Statistical Analysis
3. Results
3.1. Field Measurements
3.2. Analysis of Bioactive Compounds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Commission. COM(2020) 381 Final. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. A Farm to Fork Strategy for a Fair, Healthy and Environmentally-Friendly Food Syst; European Commission: Brussels, Belgium, 2020. [Google Scholar]
- Oberč, B.P.; Schnell, A.A. Approaches to Sustainable Agriculture. Exploring the Pathways towards the Future of Farming; IUCN European: Brussels, Belgium, 2020. [Google Scholar]
- Code, J. Alternative Agriculture. Innovations for Growing and Cultivating Diverse. In Routledge Handbook of Landscape and Food; Routledge: London, UK, 2018; pp. 125–137. [Google Scholar]
- Hirschfeld, S.; Van Acker, R. Permaculture Farmers Consistently Cultivate Perennials, Crop Diversity, Landscape Heterogeneity and Nature Conservation. Renew. Agric. Food Syst. 2019, 35, 342–351. [Google Scholar] [CrossRef]
- Biala, K.; Terres, J.-M.; Pointereau, P.; Paracchini, M.L. Low Input Farming Systems: An Opportunity to Develop Sustainable Agriculture. In Proceedings of the JRC Summer University, Ranco, Italy, 2–5 July 2007; European Commission, Joint Research Centre, Institute for Environment and Sustainability: Luxembourg, 2007; p. 127. [Google Scholar] [CrossRef]
- ISTAT. Agriculture. 2022. Available online: http://dati.istat.it/Index.aspx?DataSetCode=DCSP_COLTIVAZIONI (accessed on 4 January 2022).
- Martí, R.; Roselló, S.; Cebolla-Cornejo, J. Tomato as a Source of Carotenoids and Polyphenols Targeted to Cancer Prevention. Cancers 2016, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caseiro, M.; Ascenso, A.; Costa, A.; Creagh-Flynn, J.; Johnson, M.; Simões, S. Lycopene in Human Health. LWT 2020, 127, 109323. [Google Scholar] [CrossRef]
- Dono, G.; Picarella, M.E.; Pons, C.; Santangelo, E.; Monforte, A.; Granell, A.; Mazzucato, A. Characterization of a Repertoire of Tomato Fruit Genetic Variants in the San Marzano Genetic Background. Sci. Hortic. 2020, 261, 108927. [Google Scholar] [CrossRef]
- Ilahy, R.; Hdider, C.; Lenucci, M.S.; Tlili, I.; Dalessandro, G. Phytochemical Composition and Antioxidant Activity of High-Lycopene Tomato (Solanum lycopersicum L.) Cultivars Grown in Southern Italy. Sci. Hortic. 2011, 127, 255–261. [Google Scholar] [CrossRef]
- Mes, P.J.; Boches, P.; Myers, J.R.; Durst, R. Characterization of Tomatoes Expressing Anthocyanin in the Fruit. J. Amer. Soc. Hort. Sci. 2008, 133, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Ronen, G.; Carmel-Goren, L.; Zamir, D.; Hirschberg, J. An Alternative Pathway to Beta-Carotene Formation in Plant Chromoplasts Discovered by Map-Based Cloning of Beta and Old-Gold Color Mutations in Tomato. Proc. Natl. Acad. Sci. USA 2000, 97, 11102–11107. [Google Scholar] [CrossRef] [Green Version]
- Rosati, C.; Aquilani, R.; Dharmapuri, S.; Pallara, P.; Marusic, C.; Tavazza, R.; Bouvier, F.; Camara, B.; Giuliano, G. Metabolic Engineering of Beta-Carotene and Lycopene Content in Tomato Fruit. Plant J. 2000, 24, 413–419. [Google Scholar] [CrossRef]
- Isaacson, T.; Ronen, G.; Zamir, D.; Hirschberg, J. Cloning of Tangerine from Tomato Reveals a Carotenoid Isomerase Essential for the Production of B-Carotene and Xanthophylls in Plants. Plant Cell 2002, 14, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Levin, I.; De Vos, R.; Tadmor, Y.; Bovy, A.; Lieberman, M.; Oren-Shamis, M.; Segev, O.; Kolotilin, I.; Keller, M.; Ovadia, R.; et al. High Pigment Tomato Mutants—More than Just Lycopene (a Review). Isr. J. Plant Sci. 2006, 54, 179–190. [Google Scholar] [CrossRef]
- Sapir, M.; Oren-Shamir, M.; Ovadia, R.; Reuveni, M.; Evenor, D.; Tadmor, Y.; Nahon, S.; Shlomo, H.; Chen, L.; Meir, A.; et al. Molecular Aspects of Anthocyanin Fruit Tomato in Relation to High Pigment-1. J. Hered. 2008, 99, 292–303. [Google Scholar] [CrossRef] [Green Version]
- Gonzali, S.; Mazzucato, A.; Perata, P. Purple as a Tomato: Towards High Anthocyanin Tomatoes. Trends Plant Sci. 2009, 14, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Ciriminna, R.; Fidalgo, A.; Meneguzzo, F.; Ilharco, L.M.; Pagliaro, M.; Pais, A.R. Lycopene: Emerging Production Methods and Applications of a Valued Carotenoid. ACS Sustain. Chem. Eng. 2015, 4, 643–650. [Google Scholar] [CrossRef]
- Mustilli, A.C.; Fenzi, F.; Ciliento, R.; Alfano, F.; Bowler, C. Phenotype of the Tomato High Pigment-2 Mutant Is Caused by a Mutation in the Tomato Homolog of DEETIOLATED1. Plant Cell 1999, 11, 145–158. [Google Scholar] [CrossRef] [Green Version]
- Mazzucato, A.; Willems, D.; Bernini, R.; Picarella, M.E.; Santangelo, E.; Ruiu, F.; Tilesi, F.; Piero, G. Novel Phenotypes Related to the Breeding of Purple-Fruited Tomatoes and Effect of Peel Extracts on Human Cancer Cell Proliferation. Plant Physiol. Biochem. 2013, 72, 125–133. [Google Scholar] [CrossRef]
- Beni, C.; Neri, U.; Papetti, P.; Altimari, A. Natural Horticultural Systems in Organic Farming as a Tool for Resilience: Improvement of Economic Performance and Prevention of Soil Erosion. Agroecol. Sustain. Food Syst. 2021, 45, 1375–1398. [Google Scholar] [CrossRef]
- Landi, M.; Guidi, L.; Pardossi, A.; Tattini, M.; Gould, K.S. Photoprotection by Foliar Anthocyanins Mitigates Effects of Boron Toxicity in Sweet Basil (Ocimum basilicum). Planta 2014, 240, 941–953. [Google Scholar] [CrossRef]
- Rosales, M.A.; Ruiz, J.M.; Hernàndez, J.; Soriano, T.; Castilla, N.; Romero, L. Antioxidant Content and Ascorbate Metabolism in Cherry Tomato Exocarp in Relation to Temperature and Solar Radiation. J. Sci. Food Agric. 2006, 86, 1545–1551. [Google Scholar] [CrossRef]
- Nagata, M.; Yamashita, I. Simple Method for Simultaneous Determination of Chlorophyll and Carotenoids in Tomato Fruit. Nippon Shokuhin Kogyo Gakkaishi 1992, 39, 925–928. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, E.; Carnevale, M.; Migliori, C.A.; Picarella, M.E.; Dono, G.; Mazzucato, A.; Gallucci, F. Evaluation of Tomato Introgression Lines Diversified for Peel Color as a Source of Functional Biocompounds and Biomass for Energy Recovery. Biomass Bioenergy 2020, 141, 105735. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 17th ed.; AOAC: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Ilahy, R.; Siddiqui, M.W.; Tlili, I.; Montefusco, A.; Piro, G.; Hdider, C.; Lenucci, M.S. When Color Really Matters: Horticultural Performance and Functional Quality of High-Lycopene Tomatoes. Crit. Rev. Plant Sci. 2018, 37, 15–53. [Google Scholar] [CrossRef]
- Ronga, D.; Zaccardelli, M.; Lovelli, S.; Perrone, D.; Francia, E.; Milc, J.; Ulrici, A.; Pecchioni, N. Biomass Production and Dry Matter Partitioning of Processing Tomato under Organic vs Conventional Cropping Systems in a Mediterranean Environment. Sci. Hortic. 2017, 224, 163–170. [Google Scholar] [CrossRef]
- Ronga, D.; Francia, E.; Rizza, F.; Badeck, F.W.; Caradonia, F.; Montevecchi, G.; Pecchioni, N. Changes in Yield Components, Morphological, Physiological and Fruit Quality Traits in Processing Tomato Cultivated in Italy since the 1930’s. Sci. Hortic. 2019, 257, 108726. [Google Scholar] [CrossRef]
- Morra, L.; Cozzolino, E.; Salluzzo, A.; Modestia, F.; Bilotto, M.; Baiano, S.; del Piano, L. Plant Growth, Yields and Fruit Quality of Processing Tomato (Solanum lycopersicon L.) as Affected by the Combination of Biodegradable Mulching and Digestate. Agronomy 2021, 11, 100. [Google Scholar] [CrossRef]
- Massantini, R.; Radicetti, E.; Frangipane, M.T.; Campiglia, E. Quality of Tomato (Solanum lycopersicum L.) Changes under Different Cover Crops, Soil Tillage and Nitrogen Fertilization Management. Agriculture 2021, 11, 106. [Google Scholar] [CrossRef]
- Pavlů, L.; Kodešová, R.; Fér, M.; Nikodem, A.; Němec, F.; Prokeš, R. The Impact of Various Mulch Types on Soil Properties Controlling Water Regime of the Haplic Fluvisol. Soil Tillage Res. 2021, 205, 104748. [Google Scholar] [CrossRef]
- Rathinasabapathi, B.; Ferguson, J.; Gal, M. Evaluation of Allelopathic Potential of Wood Chips for Weed Suppression in Horticultural Production Systems. HortScience 2005, 40, 711–713. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.; Lammer, D.; Lyon, S.; Carter, B.; Jones, S.S. Breeding for Organic and Low-Input Farming Systems: An Evolutionary–Participatory Breeding Method for Inbred Cereal Grains. Renew. Agric. Food Syst. 2005, 20, 48–55. [Google Scholar] [CrossRef]
- Crespo-Herrera, L.A.; Ortiz, R. Plant Breeding for Organic Agriculture: Something New? Agric. Food Secur. 2015, 4, 25. [Google Scholar] [CrossRef]
- Bino, R.J.; De Vos, C.H.R.; Lieberman, M.; Hall, R.D.; Bovy, A.; Jonker, H.H.; Tikunov, Y.; Lommen, A.; Moco, S.; Levin, I. The Light-Hyperresponsive High Pigment-2dg Mutation of Tomato: Alterations in the Fruit Metabolome. New Phytol. 2005, 166, 427–438. [Google Scholar] [CrossRef]
- Lenucci, M.S.; Cadinu, D.; Taurino, M.; Piro, G.; Dalessandro, G. Antioxidant Composition in Cherry and High-Pigment Tomato Cultivars. J. Agric. Food Chem. 2006, 54, 2606–2613. [Google Scholar] [CrossRef]
- Toor, R.K.; Savage, G.P. Antioxidant Activity in Different Fractions of Tomato. Food Res. Int. 2005, 38, 487–494. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Chiou, A.; Pyriochou, V.; Peristeraki, A.; Karathanos, V.T. Bioactive Phytochemicals in Industrial Tomatoes and Their Processing Byproducts. LWT-Food Sci. Technol. 2012, 49, 213–216. [Google Scholar] [CrossRef]
- Szabo, K.; Cătoi, A.F.; Vodnar, D.C. Bioactive Compounds Extracted from Tomato Processing By-Products as a Source of Valuable Nutrients. Plant Foods Hum. Nutr. 2018, 73, 268–277. [Google Scholar] [CrossRef]
- Boccia, F.; Di Donato, P.; Covino, D.; Poli, A. Food Waste and Bio-Economy: A Scenario for the Italian Tomato Market. J. Clean. Prod. 2019, 227, 424–433. [Google Scholar] [CrossRef]
- Ballester, A.-R.; Molthoff, J.; de Vos, R.; Hekkert, B.t.L.; Orzaez, D.; Fernandez-Moreno, J.-P.; Tripodi, P.; Grandillo, S.; Martin, C.; Heldens, J.; et al. Biochemical and Molecular Analysis of Pink Tomatoes: Deregulated Expression of the Gene Encoding Transcription Factor SlMYB12 Leads to Pink Tomato Fruit Color. Plant Physiol. 2010, 152, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.; Rodríguez, E.; Dìaz, C. Free Hydroxycinnamic Acids, Lycopene, and Color Parameters in Tomato Free Hydroxycinnamic Acids, Lycopene, and Color Parameters in Tomato Cultivars. J. Agric. Food Chem. 2007, 55, 8604–8615. [Google Scholar] [CrossRef]
- Anton, D.; Matt, D.; Pedastsaar, P.; Bender, I.; Kazimierczak, R.; Roasto, M.; Kaart, T.; Luik, A. Three-Year Comparative Study of Polyphenol Contents and Antioxidant Capacities in Fruits of Tomato (Lycopersicon esculentum Mill.) Cultivars Grown under Organic and Conventional Conditions. J. Agric. Food Chem. 2014, 62, 5173–5180. [Google Scholar] [CrossRef]
- Lahoz, I.; Leiva-brondo, M.; Martí, R.; Ignacio, J.; Campillo, C.; Roselló, S.; Cebolla-cornejo, J. Influence of High Lycopene Varieties and Organic Farming on the Production and Quality of Processing Tomato. Sci. Hortic. 2016, 204, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Evoli, L.D.; Lucarini, M.; Sánchez, J.; Aguzzi, A.; Gabrielli, P.; Gambelli, L.; Lombardi-Boccia, G. Phenolic Acids Content and Nutritional Quality of Conventional, Organic and Biodynamic Cultivations of the Tomato CXD271BIO Breeding Line (Solanum lycopersicum L.). Food Nutr. Sci. 2016, 7, 1112–1121. [Google Scholar] [CrossRef] [Green Version]
- Heimler, D.; Romani, A.; Ieri, F. Plant Polyphenol Content, Soil Fertilization and Agricultural Management: A Review. Eur. Food Res. Technol. 2017, 243, 1107–1115. [Google Scholar] [CrossRef]
- Durazzo, A.; Azzini, E.; Foddai, M.S.; Nobili, F.; Garaguso, I.; Raguzzini, A.; Finotti, E.; Tisselli, V.; Del Vecchio, S.; Piazza, C.; et al. Influence of Different Crop Management Practices on the Nutritional Properties and Benefits of Tomato -Lycopersicon esculentum Cv Perfectpeel-. Int. J. Food Sci. Technol. 2010, 45, 2637–2644. [Google Scholar] [CrossRef]
- Lu, Z.; Wang, J.; Gao, R.; Ye, F.; Zhao, G. Sustainable Valorisation of Tomato Pomace: A Comprehensive Review. Trends Food Sci. Technol. 2019, 86, 172–187. [Google Scholar] [CrossRef]
- Bennet, R.N.; Wallsgrove, R.M. Secondary Metabolites in Plant Defence Mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Agati, G.; Tattini, M. Multiple Functional Roles of Flavonoids in Photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line/Variety | Mutant Genes | Modified Bioactive Compound Class | Fruit Type | Code | Ref. |
|---|---|---|---|---|---|
| V711392 (Breeding line) | hp-2 | All | Round, flesh and peel red | 392 | [15] |
| V710446 (Solenero®) | Aft atv | Polyphenols (Plant and fruits) | Round, red flesh, black peel | 446 | [20] |
| Rio Grande (Processing variety) | - | None | Oval, flesh and peel red | RG |
| Soil Properties | U.M. * | |
|---|---|---|
| Sand | % | 25 |
| Silt | % | 38 |
| Clay | % | 37 |
| pH | 7.9 | |
| Total nitrogen (N) | % | 0.16 |
| Assimilable phosphorous (P) | mg kg−1 | 14.1 |
| Exchangeable potassium (K) | mg kg−1 | 365.3 |
| Product | Farming System | Dose | N. of Treatments |
|---|---|---|---|
| Copper sulphate | LI and LIMI | 3 g L−1 | 1 |
| NeemAzal ® | LI | 2 mL L−1 | 1 |
| Rapax ® (Bacillus thuringensis) | LI and LIMI | 2 mL L−1 | 3 |
| Tioflor | LI | 5 g L−1 | 2 |
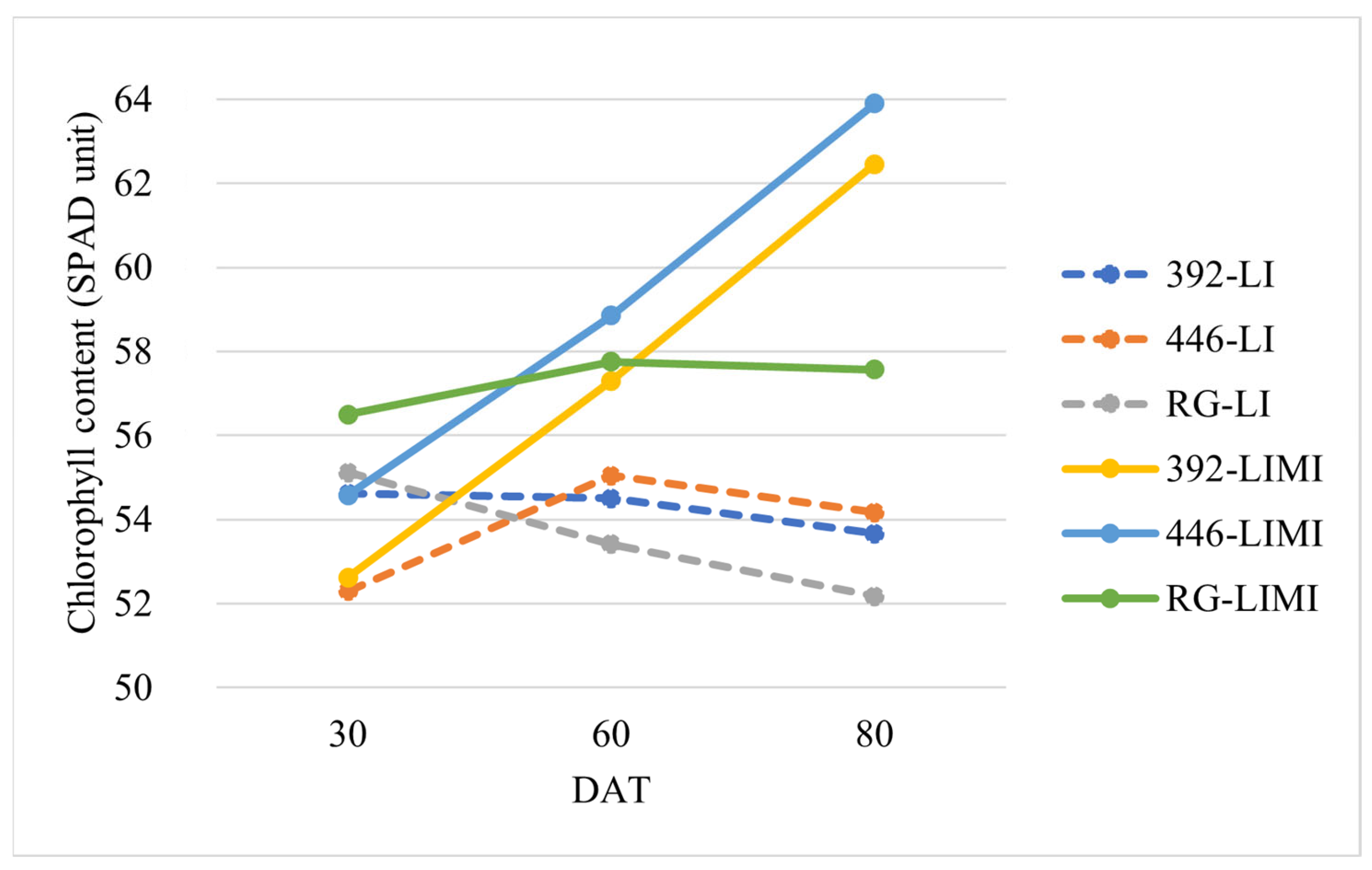
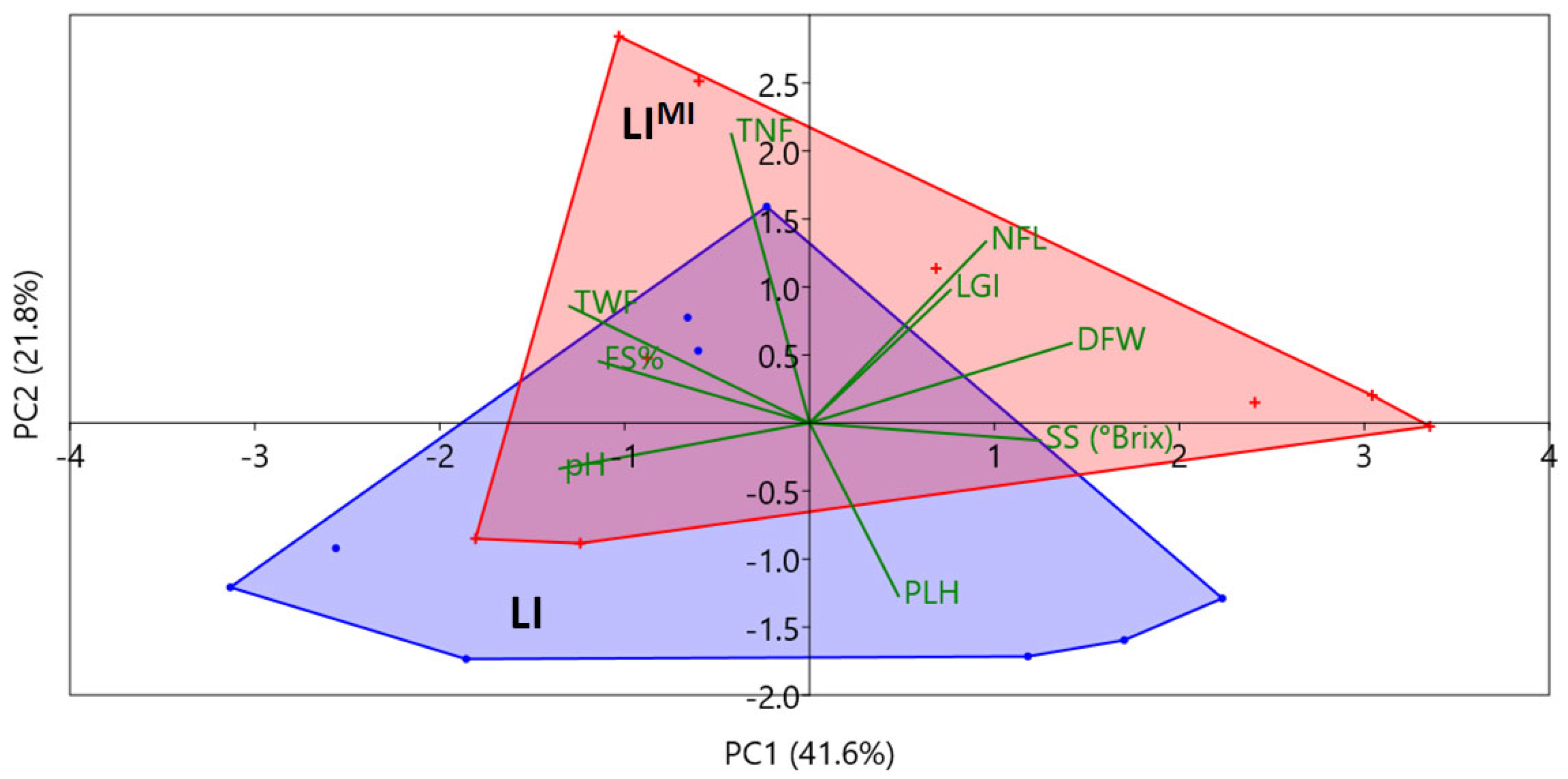
| Factor | PLH § (cm) | DFW (days) | NFL (n.) | LGI (Spad Unit) |
|---|---|---|---|---|
| System | ||||
| LI | 56.1 | 27 | 6.1 | 53.3 b |
| LIMI | 55.3 | 28 | 7.1 | 61.3 a |
| Genotype | ||||
| 392 | 49.3 | 29 B | 7.5 a | 58.1 |
| 446 | 61.0 | 33 A | 7.1 a | 59.0 |
| RG | 56.8 | 20 C | 5.3 b | 54.9 |
| Sys × Gen | ||||
| 392-LI | 49.8 | 29 | 7.8 | 53.7 |
| 446-LI | 62.6 | 32 | 6.0 | 54.2 |
| RG-LI | 56.0 | 20 | 4.7 | 52.2 |
| 392 LIMI | 48.8 | 29 | 7.2 | 62.5 |
| 446 LIMI | 59.4 | 34 | 8.1 | 63.9 |
| RG LIMI | 57.6 | 20 | 5.9 | 57.6 |
| Significance | ||||
| System | ns | ns | ns | * |
| Genotype | ns | ** | * | ns |
| Sys × Gen | ns | ns | ns | ns |
| Factor | FS (%) § | TNF (n.) | TWF (g) | pH | SS (°Brix) |
|---|---|---|---|---|---|
| System | |||||
| LI | 63.3 | 46 B | 2195.0 b | 4.46 a | 4.28 |
| LIMI | 52.1 | 70 A | 2716.6 a | 4.36 b | 4.31 |
| Genotype | |||||
| 392 | 65.7 A | 93 A | 2995.9 A | 4.40 AB | 4.18 B |
| 446 | 47.5 B | 34 B | 898.4 B | 4.24 B | 4.87 A |
| RG | 59.8 A | 46 B | 3473.1 A | 4.59 A | 3.83 B |
| Sys × Gen | |||||
| 392-LI | 66.6 A | 78 | 2780.0 | 4.49 | 4.40 bc |
| 446-LI | 51.6 B | 22 | 763.9 | 4.21 | 4.53 b |
| RG-LI | 71.7 A | 38 | 3041.2 | 4.68 | 3.90 bc |
| 392 LIMI | 64.8 A | 109 | 3211.8 | 4.32 | 3.97 bc |
| 446 LIMI | 43.4 B | 47 | 1032.9 | 4.26 | 5.20 a |
| RG LIMI | 48.0 B | 55 | 3905.0 | 4.50 | 3.77 c |
| Significance | |||||
| System | ns | ** | * | * | ns |
| Genotype | ** | ** | ** | ** | ** |
| Sys × Gen | ** | ns | ns | ns | * |
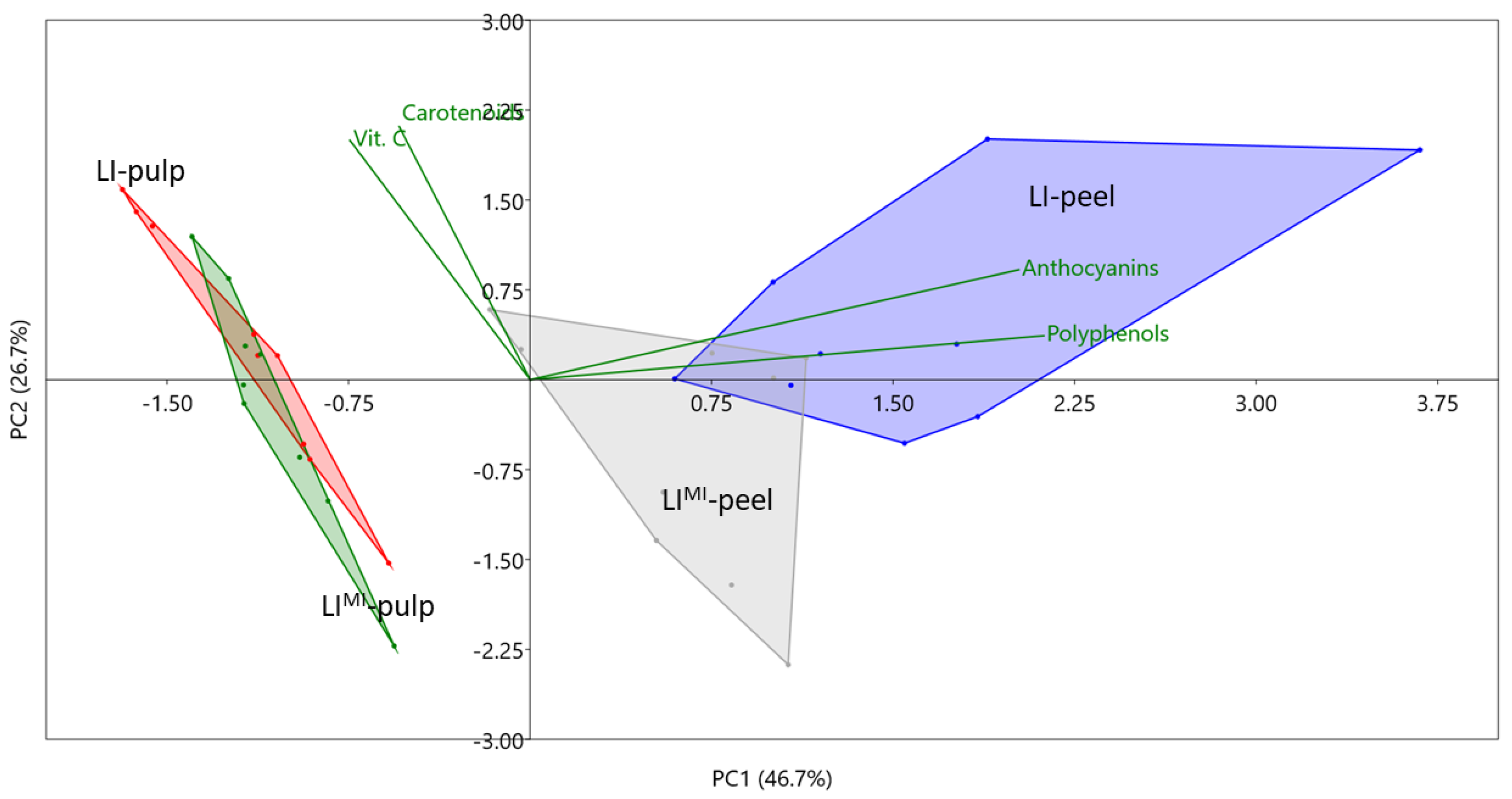
| Factor | Vit. C | Carotenoids | Polyphenols | Anthocyanins |
|---|---|---|---|---|
| System | ||||
| LI | 13.36 | 24.84 | 109.96 A | 0.91 a |
| LIMI | 11.02 | 23.25 | 79.76 B | 0.42 b |
| Genotype | ||||
| 392 | 10.49 B | 25.48 | 102.26 B | 0.26 b |
| 446 | 15.05 A | 22.40 | 107.83 A | 0.85 a |
| RG | 11.03 B | 24.27 | 74.50 C | 0.88 a |
| Sys × Gen | ||||
| 392 LI | 9.87 B | 25.70 | 118.54 B | 0.51 bc |
| 446 LI | 20.23 A | 24.65 | 127.37 A | 1.48 a |
| RG LI | 9.97 B | 24.18 | 83.98 C | 0.74 ac |
| 392 LIMI | 11.10 B | 25.25 | 85.97 C | 0.02 c |
| 446 LIMI | 9.87 B | 20.15 | 88.29 C | 0.21 c |
| RG LIMI | 12.08 B | 24.35 | 65.01 D | 1.02 ab |
| Significance | ||||
| System | ns | ns | ** | * |
| Genotype | ** | ns | ** | * |
| Sys × Gen | ** | ns | ** | * |
| Factor | Vit. C | Carotenoids | Polyphenols | Anthocyanins |
|---|---|---|---|---|
| System | ||||
| LI | 19.92 A | 24.69 | 33.31 a | 0.002 |
| LIMI | 13.83 B | 25.07 | 30.00 b | 0.002 |
| Genotype | ||||
| 392 | 13.91 B | 25.10 | 30.44 b | 0.002 |
| 446 | 27.43 A | 25.28 | 34.43 a | 0.003 |
| RG | 9.29 C | 24.27 | 30.18 b | 0.002 |
| Sys × Gen | ||||
| 392-LI | 13.90 C | 25.51 | 34.02 a | 0.001 |
| 446-LI | 34.27 A | 24.36 | 33.25 a | 0.004 |
| RG-LI | 11.60 CD | 24.21 | 32.67 a | 0.002 |
| 392 LIMI | 13.92 C | 24.68 | 26.86 b | 0.002 |
| 446 LIMI | 20.60 B | 26.20 | 35.60 a | 0.001 |
| RG-LIMI | 6.97 D | 24.34 | 27.69 b | 0.002 |
| Significance | ||||
| System | ** | ns | * | ns |
| Genotype | ** | ns | * | ns |
| Sys × Gen | ** | ns | * | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santangelo, E.; Bergonzoli, S.; Papetti, P.; Mazzucato, A.; Del Giudice, A.; Scarfone, A.; Beni, C. The Profile of Bioactive Compounds in Two Tomato Lines (hp-2 and Atv_aft) When Grown under Low-Input Farming Conditions with or without Mulching and Intercropping. Agriculture 2023, 13, 280. https://doi.org/10.3390/agriculture13020280
Santangelo E, Bergonzoli S, Papetti P, Mazzucato A, Del Giudice A, Scarfone A, Beni C. The Profile of Bioactive Compounds in Two Tomato Lines (hp-2 and Atv_aft) When Grown under Low-Input Farming Conditions with or without Mulching and Intercropping. Agriculture. 2023; 13(2):280. https://doi.org/10.3390/agriculture13020280
Chicago/Turabian StyleSantangelo, Enrico, Simone Bergonzoli, Patrizia Papetti, Andrea Mazzucato, Angelo Del Giudice, Antonio Scarfone, and Claudio Beni. 2023. "The Profile of Bioactive Compounds in Two Tomato Lines (hp-2 and Atv_aft) When Grown under Low-Input Farming Conditions with or without Mulching and Intercropping" Agriculture 13, no. 2: 280. https://doi.org/10.3390/agriculture13020280