
Overexpression of CsGSH2 Alleviates Propamocarb Residues and Phytotoxicity in Cucumber by Enhancing Antioxidant and Glutathione Detoxification Properties

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Treatments
2.2. Determination of Propamocarb Residues
2.3. Lipid Peroxidation and ROS Levels
2.4. Real-Time Quantitative PCR
2.5. Subcellular Localization
2.6. CsGSH2 Gene Construction and Transformation
2.7. Determination of Enzyme Activities and AsA-GSH Cycle
2.8. Statistical Analysis
3. Results
3.1. Propamocarb Stress Induces Lipid Peroxidation and ROS Accumulation in Cucumber
3.2. CsGSH2 Regulates Glutathione Metabolism under Propamocarb Stress
3.3. CsGSH2 Is a Cytosolic Protein
3.4. Cloning and Bioinformatic Analysis of CsGSH2
3.5. Overexpression of CsGSH2 Alleviates Propamocarb Residues in Cucumber and Arabidopsis
3.6. Overexpression of CsGSH2 Helps Propamocarb Detoxification
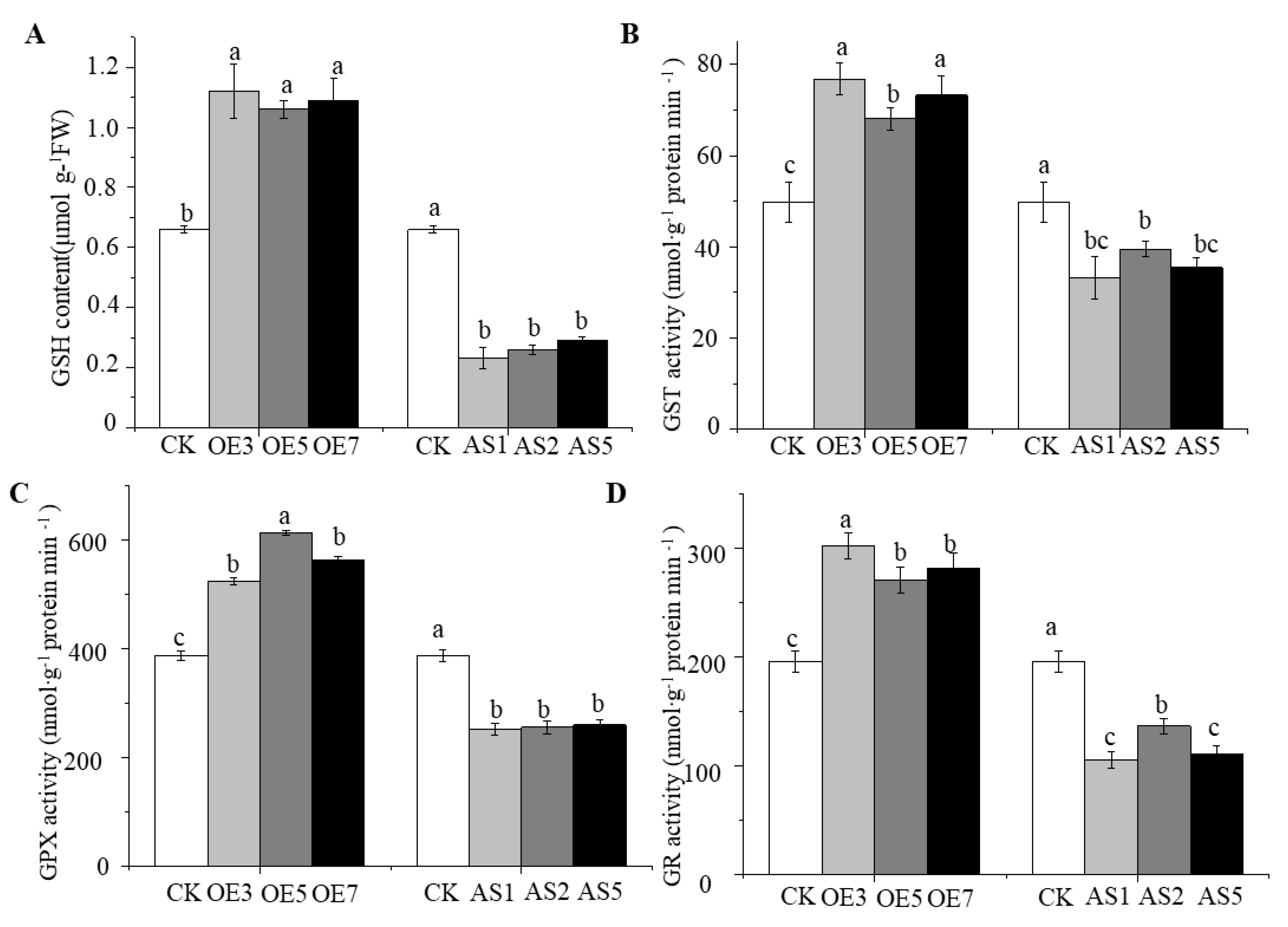
3.7. Overexpression of CsGSH2 Enhances Antioxidant Enzyme Activities in Response to Propamocarb Phytotoxicity
3.8. Overexpression of CsGSH2 Moderates the AsA-GSH Cycle to Reduce ROS Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Z.; Jiang, Y.; Peng, X.; Xu, S.; Zhang, H.; Gao, J.; Xi, Z. Exogenous 24-epibrassinolide regulates antioxidant and pesticide detoxification systems in grapevine after chlorothalonil treatment. Plant Growth Regul. 2016, 81, 455–466. [Google Scholar] [CrossRef]
- Li, R.; Zhao, J.; Wu, Y.H.; Hu, F.F. Residual trend of 66.5% propamocarb hydrochloride aqueous solutions in cucumber. J. Biosaf. 2020, 30, 132–136. [Google Scholar] [CrossRef]
- Gangemi, S.; Gofita, E.; Costa, C.; Teodoro, M.; Briguglio, G.; Nikitovic, D.; Tzanakakis, G.; Tsatsakis, A.M.; Wilks, M.F.; Spandidos, D.A.; et al. Occupational and environmental exposure to pesticides and cytokine pathways in chronic diseases. Int. J. Mol. Med. 2016, 38, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Li, S.N.; Xin, M.; Luan, J.; Liu, D.; Wang, C.H.; Liu, C.L.; Zhang, W.S.; Zhou, X.Y.; Qin, Z.W. Overexpression of CsHMGB alleviates phytotoxicity and propamocarb residues in cucumber. Front. Plant. Sci. 2020, 11, 738. [Google Scholar] [CrossRef]
- Yin, Y.L.; Zhou, Y.; Zhou, Y.H.; Shi, K.; Zhou, J.; Yu, Y.; Yu, J.Q.; Xia, X.J. Interplay between mitogen-activated protein kinase and nitric oxide in brassinosteroid–induced pesticide metabolism in Solanum lycopersicum. J. Hazard Mater. 2016, 316, 221–231. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, S.; Ding, F. Melatonin mitigates chilling-induced oxidative stress and photosynthesis inhibition in tomato plants. Antioxidants 2020, 9, 218. [Google Scholar] [CrossRef]
- Zhu, M.D.; Zhang, M.; Gao, D.J.; Zhou, K.; Tang, S.J.; Zhou, B.; Lv, Y.M. Rice OsHSFA3 gene improves drought tolerance by modulating polyamine biosynthesis depending on abscisic acid and ros levels. Int. J. Mol. Sci. 2020, 21, 1857. [Google Scholar] [CrossRef]
- Hou, L.Y.; Lehmann, M.; Geigenberger, P. Thioredoxin h2 and o1 Show Different Subcellular Localizations and Redox-Active Functions, and Are Extrachloroplastic Factors Influencing Photosynthetic Performance in Fluctuating Light. Antioxidants 2021, 10, 705. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Anee, T.I.; Parvin, K.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Guan, L.; Zhang, P.; Wang, X.K.; Ren, Y.P.; Guo, B.B.; Liu, F. Photodegradation of pyraclostrobin in water environment and microencapsulation effect on its photostability. J. Agro. Environ. Sci. 2015, 34, 1493–1497. [Google Scholar] [CrossRef]
- Liu, N.; Li, J.; Lv, J.; Yu, J.; Xie, J.; Wu, Y.; Tang, Z. Melatonin alleviates Imidacloprid phytotoxicity to cucumber (Cucumis sativus L.) through modulating redox homeostasis in plants and promoting its metabolism by enhancing glutathione dependent detoxification. Ecotoxicol. Environ. Saf. 2021, 217, 112248. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xia, X.; Yu, G.; Wang, J.; Wu, J.; Wang, M.; Yang, Y.; Shi, K.; Yu, Y.; Chen, Z. Brassino steroids play a critical role in the regulation of pesticide meta-bolism in crop plants. Sci. Rep. 2015, 5, 9018. [Google Scholar] [CrossRef] [PubMed]
- Chin, D.C.; Hsieh, C.C.; Lin, H.Y.; Yeh, K.W. Low Glutathione Redox State Couples with a Decreased Ascorbate Redox Ratio to Accelerate Flowering in Oncidium Orchid. Plant Cell Physiol. 2016, 57, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Bhardwaj, R.; Kumar, V.; Thukral, A.K. Gc-Ms Studies Reveal Stimulated Pesticide Detoxification by Brassinolide Application in Brassica juncea L. plants. Environ. Sci. Pollut. Res. 2016, 23, 14518–14525. [Google Scholar] [CrossRef]
- Bela, K.; Horváth, E.; Gallé, Á.; Szabados, L.; Tari, I.; Csiszár, J. Plant glutathione peroxidases: Emerging role of the antioxidant enzymes in plant development and stress responses. J. Plant Physiol. 2015, 176, 192–201. [Google Scholar] [CrossRef]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Verma, P.K.; Verma, S.; Meher, A.K.; Pande, V.; Mallick, S.; Bansiwal, A.K.; Tripathi, R.D.; Dhankher, O.M.; Chakrabarty, D. Overexpression of rice glutaredoxins (OsGrxs) significantly reduces arsenite accumulation by maintaining glutathione pool and modulating aquaporins in yeast. Plant Physiol. Biochem. 2016, 106, 208–217. [Google Scholar] [CrossRef]
- Jia, C.C.; Ji, J.; Wang, G.; Tian, X.W.; Du, X.L.; Guan, C.F.; Jin, C.; Wu, D.Y. Over-expression of Glutathione Synthetase Gene Enhances CadmiumTolerance in Transgenic Tobacco Plant. China Biotenol. 2014, 34, 79–86. [Google Scholar] [CrossRef]
- Liu, D.L.; Ma, L.B.; Guo, A.Y.; Yang, H.; Lu, Z.Q. Cloning of Beta vulgaris Glutathione Synthetase (BvGS) Gene under Cadmium Stress. Sugar Crops. China 2017, 39, 23–25. [Google Scholar] [CrossRef]
- Gullner, G.; Kömives, T.; Rennenberg, H. Enhanced tolerance of transgenic poplar plants overexpressing γ-glutamylcysteine synthetase towards chloroacetanilide herbicides. J. Exp. Bot. 2001, 52, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Qin, Z.; Zhao, W.; Zhou, X.Y.; Wu, T. Transcriptome Analysis Reveals Differentially Expressed Genes Associated with Propamocarb Response in Cucumber (Cucumis sativus L.) Fruit. Acta Physiol. Plant. 2013, 35, 2393–2406. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Zhao, Z.; Qian, C.; Sui, Y.; Malik, A.A.; Chen, J. Selection of appropriate reference genes for gene expression studies by quantitative real–time polymerase chain reaction in cucumber. Anal. Biochem. 2010, 399, 257–261. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Bechtold, N.; Pelletier, G. In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Molecular Biol. 1998, 82, 259–266. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Liu, B.; Wang, W.; Liu, X. A GAMYB homologue CsGAMYB1 regulates sex expression of cucumber via an ethylene independent pathway. J. Exp. Bot. 2014, 65, 3201–3213. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein using theprinciple of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Giannakoula, A.; Moustakas, M.; Syros, T.; Yupsanis, T. Aluminium stress induces up-regulation of an efficient antioxidant system in the Al-tolerant maize line but not in the Al-sensitive line. Environ. Exp. Bot. 2010, 67, 487–494. [Google Scholar] [CrossRef]
- Durner, J.; Klessig, D.F. Inhibition of ascorbate peroxidase by salicylic acid and 2,6-dichloroisonicotinic acid, two inducers of plant defense responses. Proc. Natl. Acad. Sci. USA 1995, 92, 11312–11316. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.V.; Davis, D.G. Abiotic stress alters transcript profiles and activity of glutathione S-transferase, glutathione peroxidase, and glutathione reductase in Euphorbia esula. Physiol. Plant. 2004, 120, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Yuan, L.Y.; Guo, S.R.; Sun, J.; Yuan, Y.H. Effects of exogenous spermine on chlorophyll fluorescence, antioxidant system and ultrastructure of chloroplasts in Cucumis sativus L. under salt stress. Plant Physiol. Biochem. 2013, 63, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef]
- Anderson, J.V.; Chevone, B.I.; Hess, J.L. Seasonal variation in the antioxidant system of eastern white pine needles: Evidence for thermal dependence. Plant Physiol. 1992, 98, 501–508. [Google Scholar] [CrossRef]
- Siddique, Z.; Malik, A.U.; Asi, M.R.; Anwar, R.; Inam Ur Raheem, M. Sonolytic-ozonation technology for sanitizing microbial contaminants and pesticide residues from spinach (Spinacia oleracea L.) leaves, at household level. Environ. Sci. Pollut. Res. 2021, 28, 52913–52924. [Google Scholar] [CrossRef]
- Pan, L.; Yu, Q.; Wang, J.; Han, H.; Mao, L.; Nyporko, A.; Maguza, A.; Fan, L.J.; Bai, L.Y.; Powles, S. An ABCC-type transporter endowing glyphosate resistance in plants. Proc. Natl. Acad. Sci. USA 2021, 118, 2100136118. [Google Scholar] [CrossRef]
- Yan, Y.; Sun, S.; Zhao, N.; Yang, W.; Shi, Q.; Gong, B. COMT1 overexpression resulting in increased melatonin biosynthesis contributes to the alleviation of carbendazim phytotoxicity and residues in tomato plants. Environ. Pollut. 2019, 252, 51–61. [Google Scholar] [CrossRef]
- Liu, C.; Qin, Z.; Zhou, X.; Xin, M.; Wang, C.; Liu, D.; Li, S. Expression and functional analysis of the propamocarb related gene CsDIR16 in cucumbers. BMC Plant Biol. 2018, 18, 16. [Google Scholar] [CrossRef]
- Wu, S.; Luo, T.; Wang, S.; Zhou, J.; Ni, Y.; Fu, Z.; Jin, Y. Chronic exposure to fungicide propamocarb induces bile acid metabolic disorder and increases trimethylamine in C57BL/6J mice. Sci. Total Environ. 2018, 642, 341–348. [Google Scholar] [CrossRef]
- Meng, J.J.; Qin, Z.W.; Zhou, X.Y.; Xin, M. An Atp-binding cassettetransporter gene from Cucumis sativus L., CsABC19, is involved in propamocarb stress in Arabidopsis thaliana. Plant Mol. Biol. Rep. 2016, 34, 947–960. [Google Scholar] [CrossRef]
- Zhang, F.; Xin, M.; Yu, S.Q.; Liu, D.; Zhou, X.Y.; Qin, Z.W. Expression and functional analysis of the propamocarb-related gene CsMCF in cucumber. Front. Plant Sci. 2019, 10, 871. [Google Scholar] [CrossRef] [PubMed]
- Marty, L.; Bausewein, D.; Müller, C.; Bangash, S.A.K.; Moseler, A.; Schwarzländer, M.; Müller–Schüssele, S.J.; Zechmann, B.; Riondet, C.; Balk, J.; et al. Arabidopsis glutathione reductase 2 is indispensable in plastids, while mitochondrial glutathione is safeguarded by additional reduction and transport systems. New Phytol. 2019, 224, 1569–1584. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.W.; Zhao, L.X.; Gao, S.; Leng, X.Y.; Yu, Y.; Fu, Y.; Ye, F. Quinoxaline derivatives as herbicide safeners by improving Zea mays tolerance. Pestic. Biochem. Physiol. 2021, 179, 104958. [Google Scholar] [CrossRef]
- Zhang, T.; Tsutsuki, H.; Li, X.; Sawa, T. New insights into the regulatory roles of glutathione in NLRP3-inflammasome-mediated immune and inflammatory responses. The J. Biochem. 2022, 171, 367–377. [Google Scholar] [CrossRef]
- Xing, C.; Li, J.; Lam, S.M.; Yuan, H.; Shui, G.; Yang, J. The role of glutathione-mediated triacylglycerol synthesis in the response to ultra-high cadmium stress in Auxenochlorella protothecoides. J. Environ. Sci. 2021, 108, 58–69. [Google Scholar] [CrossRef]
- Sun, C.; Liu, L.; Yu, Y.; Liu, W.; Lu, L.; Jin, C.; Lin, X. Nitric oxide alleviates aluminum-induced oxidative damage through regulating the ascorbate-glutathione cycle in roots of wheat. J. Integr. Plant Biol. 2015, 57, 550–561. [Google Scholar] [CrossRef]
- Wang, D.; Liu, Y.; Tan, X.; Liu, H.; Zeng, G.; Hu, X.; Jian, H.; Gu, Y. Effect of exogenous nitric oxide on antioxidative system and S-nitrosylation in leaves of Boehmeria nivea (L.) Gaud under cadmium stress. Environ. Sci. Pollut. Res. Int. 2015, 22, 3489–3497. [Google Scholar] [CrossRef]
- Li, J.S.; Suzui, N.; Nakai, Y.; Yin, Y.G.; Ishii, S.; Fujimaki, S.; Nao, K.; Hiroki, R.; Takashi, M.; Kanna, S.L.; et al. Shoot base responds to root-applied glutathione and functions as a critical region to inhibit cadmium translocation from the roots to shoots in oilseed rape (Brassica napus). Plant Sci. 2021, 305, 110822. [Google Scholar] [CrossRef]
- Zeng, F.; Mallhi, Z.I.; Khan, N.; Rizwan, M.; Ali, S.; Ahmad, A.; Hussain, A.; Alsahli, A.A.; Alyemeni, M.N. Combined citric acid and glutathione augments lead (Pb) stress tolerance and phytoremediation of castorbean through antioxidant machinery and Pb uptake. Sustainability 2021, 13, 4073. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Wang, Y.Q.; Mao, Q.; Wu, M.J.; Yan, Y.R.; Ren, J.; Wang, X.J.; Liu, A.R.; Chen, S.C. Dopamine alleviates bisphenol A-induced phytotoxicity by enhancing antioxidant and detoxification potential in cucumber. Environ. Pollut. 2020, 259, 113957. [Google Scholar] [CrossRef]
- Li, G.Z.; Zheng, Y.X.; Chen, S.J.; Liu, J.; Wang, P.F.; Wang, Y.H.; Guo, T.C.; Kang, G.Z. TaWRKY74 participates copper tolerance through regulation of TaGST1 expression and GSH content in wheat. Ecotoxicol. Environ. Saf. 2021, 221, 112469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Qin, Z.W.; Zhou, X.Y.; Xin, M.; Li, S.N.; Luan, J. Expression and functional analysis of the propamocarb-related gene CsMAPEG in cucumber. BMC Plant Biol. 2019, 19, 371. [Google Scholar] [CrossRef] [PubMed]
- Cengiz, K.; Muhammad, A. Sodium hydrosulfite together with silicon detoxifies arsenic toxicity in tomato plants by modulating the AsA-GSH cycle. Environ. Pollut. 2021, 294, 118608. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | TPM/Control | TPM/Propamocarb | log2 Ratio (Propamocarb/Control) | FDR (False Discovery Rate) | Blast Annotation | Enzyme NO. |
|---|---|---|---|---|---|---|
| Csa4M303130 | 47.19 | 111.91 | 2.04 | 1.63 × 10−12 | Glutathione S-transferase (GST) | 2.5.1.18 |
| Csa3M133380 | 8.56 | 35.24 | 1.25 | 1.56 × 10−11 | ||
| Csa5M409710 | 22.91 | 86.82 | 1.92 | 6.84 × 10−12 | ||
| Csa7M395820 | 11.04 | 40.59 | 1.88 | 6.45 × 10−12 | ||
| Csa3M597320 | 10.49 | 25.65 | 1.29 | 2.29 × 10−5 | Ribonucleoside-diphosphate reductase small chain (RDR) | 1.17.4.1 |
| Csa1M571280 | 2.21 | 13.81 | 2.64 | 5.71 × 10−6 | Glutathione synthetase (GSH) | 6.3.2.2 |
| Lines | AsA Content | DHA Content | Total Ascorbate | AsA/DHA | GSH Content | GSSG Content | Total Glutathione | GSH/GSSG |
|---|---|---|---|---|---|---|---|---|
| CK | 1.42 ± 0.015 b | 1.37 ± 0.031 b | 2.79 ± 0.016 c | 1.04 ± 0.037 b | 0.66 ± 0.029 b | 0.28 ± 0.009 b | 0.94 ± 0.024 b | 2.36 ± 0.052 c |
| OE3 | 2.21 ± 0.038 a | 1.01 ± 0.019 d | 3.22 ± 0.032 a | 2.19 ± 0.022 a | 1.08 ± 0.056 a | 0.31 ± 0.015 a | 1.39 ± 0.076 a | 3.48 ± 0.043 a |
| OE5 | 2.34 ± 0.013 a | 1.14 ± 0.021 c | 3.48 ± 0.047 a | 2.05 ± 0.016 a | 0.98 ± 0.033 a | 0.33 ± 0.007 a | 1.34 ± 0.046 a | 2.97 ± 0.032 b |
| OE7 | 2.15 ± 0.047 a | 1.02 ± 0.027 d | 3.17 ± 0.026 a | 2.11 ± 0.013 a | 1.02 ± 0.017 a | 0.32 ± 0.010 a | 1.34 ± 0.028 a | 3.19 ± 0.022 a |
| AS1 | 1.02 ± 0.011 c | 1.58 ± 0.020 a | 2.60 ± 0.009 b | 0.65 ± 0.045 c | 0.37 ± 0.008 c | 0.17 ± 0.004 c | 0.54 ± 0.014 c | 2.18 ± 0.045 d |
| AS2 | 0.92 ± 0.009 c | 1.43 ± 0.015 b | 2.35 ± 0.028 b | 0.64 ± 0.023 c | 0.39 ± 0.006 c | 0.19 ± 0.011 c | 0.58 ± 0.015 c | 2.05 ± 0.024 d |
| AS5 | 0.97 ± 0.026 c | 1.56 ± 0.008 a | 2.53 ± 0.018 b | 0.62 ± 0.021 c | 0.41 ± 0.016 c | 0.21 ± 0.012 c | 0.62 ± 0.031 c | 1.95 ± 0.033 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wu, Z.; Liu, C.; Fan, L.; He, Y.; Lu, K.; Liu, D.; Feng, G. Overexpression of CsGSH2 Alleviates Propamocarb Residues and Phytotoxicity in Cucumber by Enhancing Antioxidant and Glutathione Detoxification Properties. Agriculture 2022, 12, 1528. https://doi.org/10.3390/agriculture12101528
Li S, Wu Z, Liu C, Fan L, He Y, Lu K, Liu D, Feng G. Overexpression of CsGSH2 Alleviates Propamocarb Residues and Phytotoxicity in Cucumber by Enhancing Antioxidant and Glutathione Detoxification Properties. Agriculture. 2022; 12(10):1528. https://doi.org/10.3390/agriculture12101528
Chicago/Turabian StyleLi, Shengnan, Zedong Wu, Chunhong Liu, Lianxue Fan, Yongheng He, Ke Lu, Dajun Liu, and Guojun Feng. 2022. "Overexpression of CsGSH2 Alleviates Propamocarb Residues and Phytotoxicity in Cucumber by Enhancing Antioxidant and Glutathione Detoxification Properties" Agriculture 12, no. 10: 1528. https://doi.org/10.3390/agriculture12101528