
The Role of Water Relations and Oxidative Stress in the Vase Life Response to Prolonged Storage: A Case Study in Chrysanthemum

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Visual Quality Characteristics, Mass Partitioning and Stem Strength
2.3. Effect of Storage Period on Leaf and Flower Reflectance
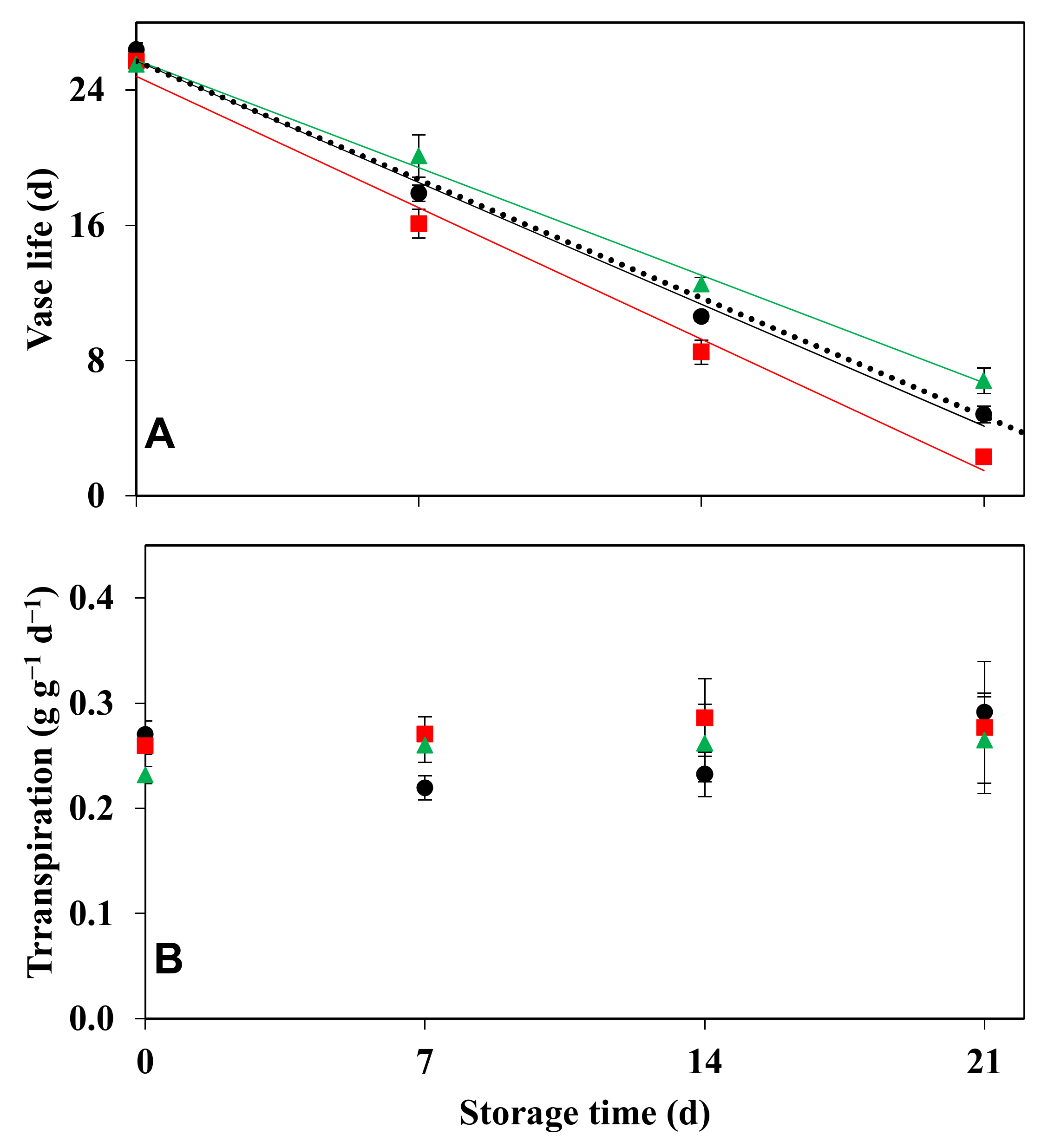
2.4. Effect of Storage Period on Vase Life
2.5. Investigation of Cut Flower Traits Potentially Related to Vase Life Response to Storage Period
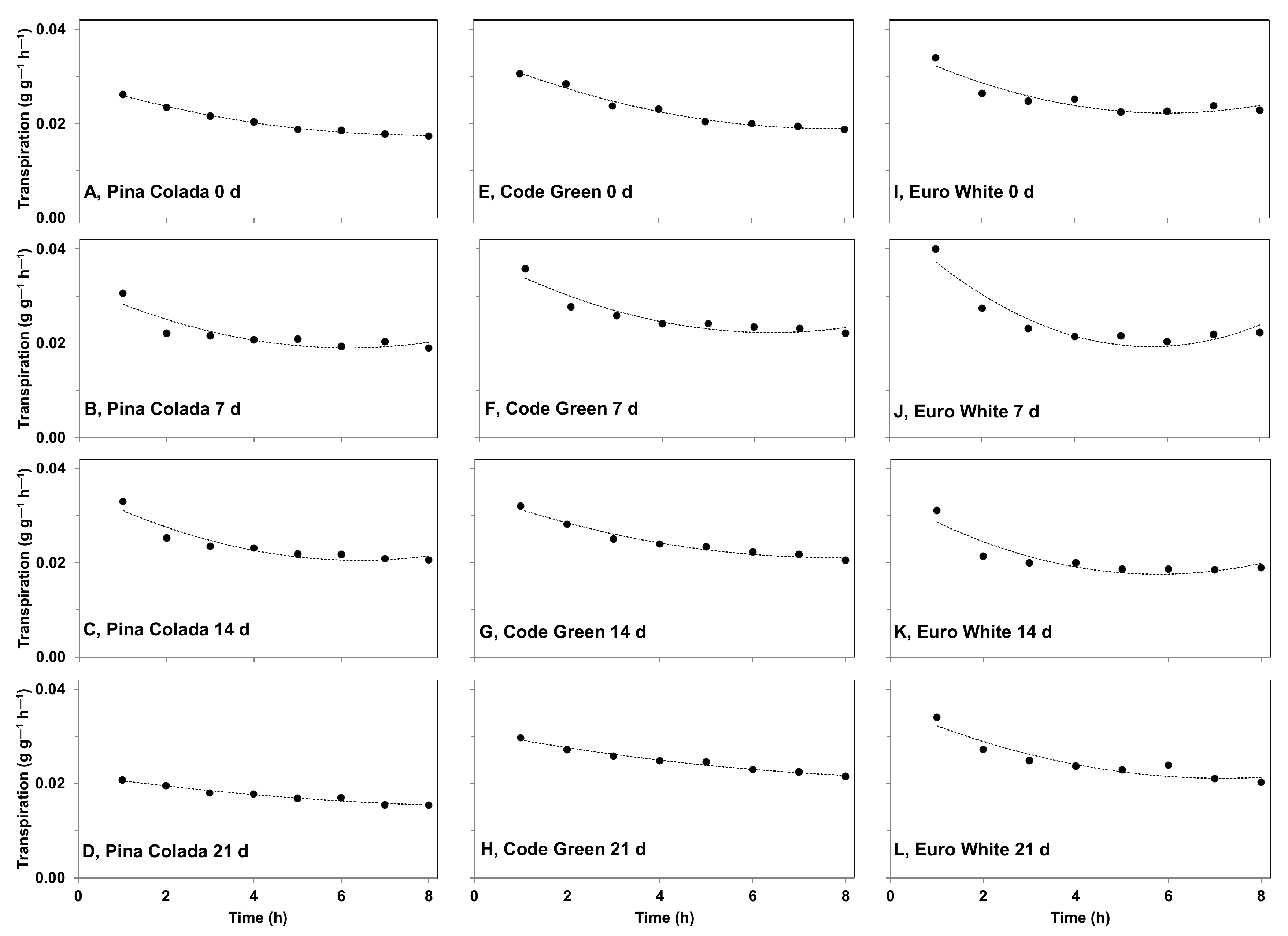
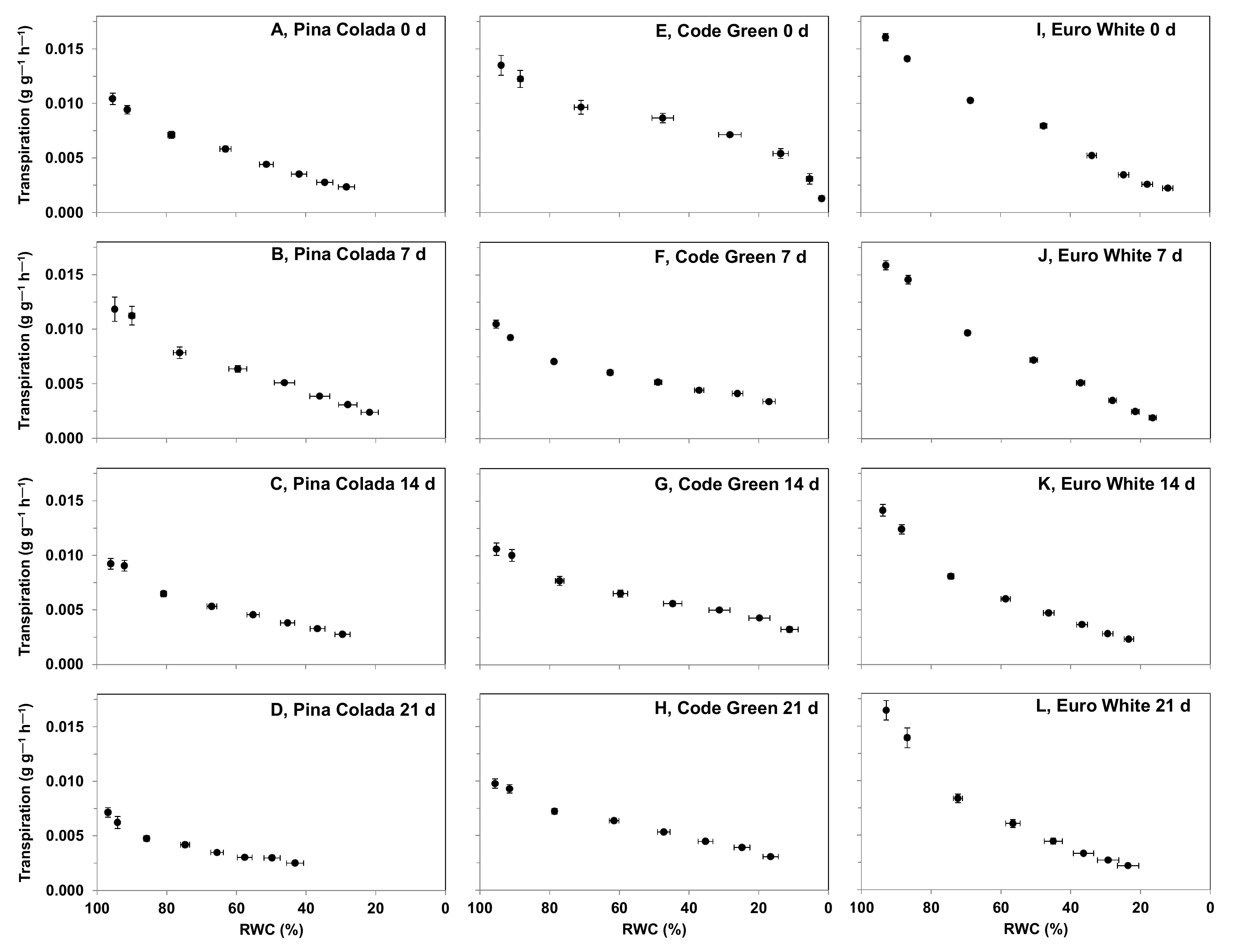
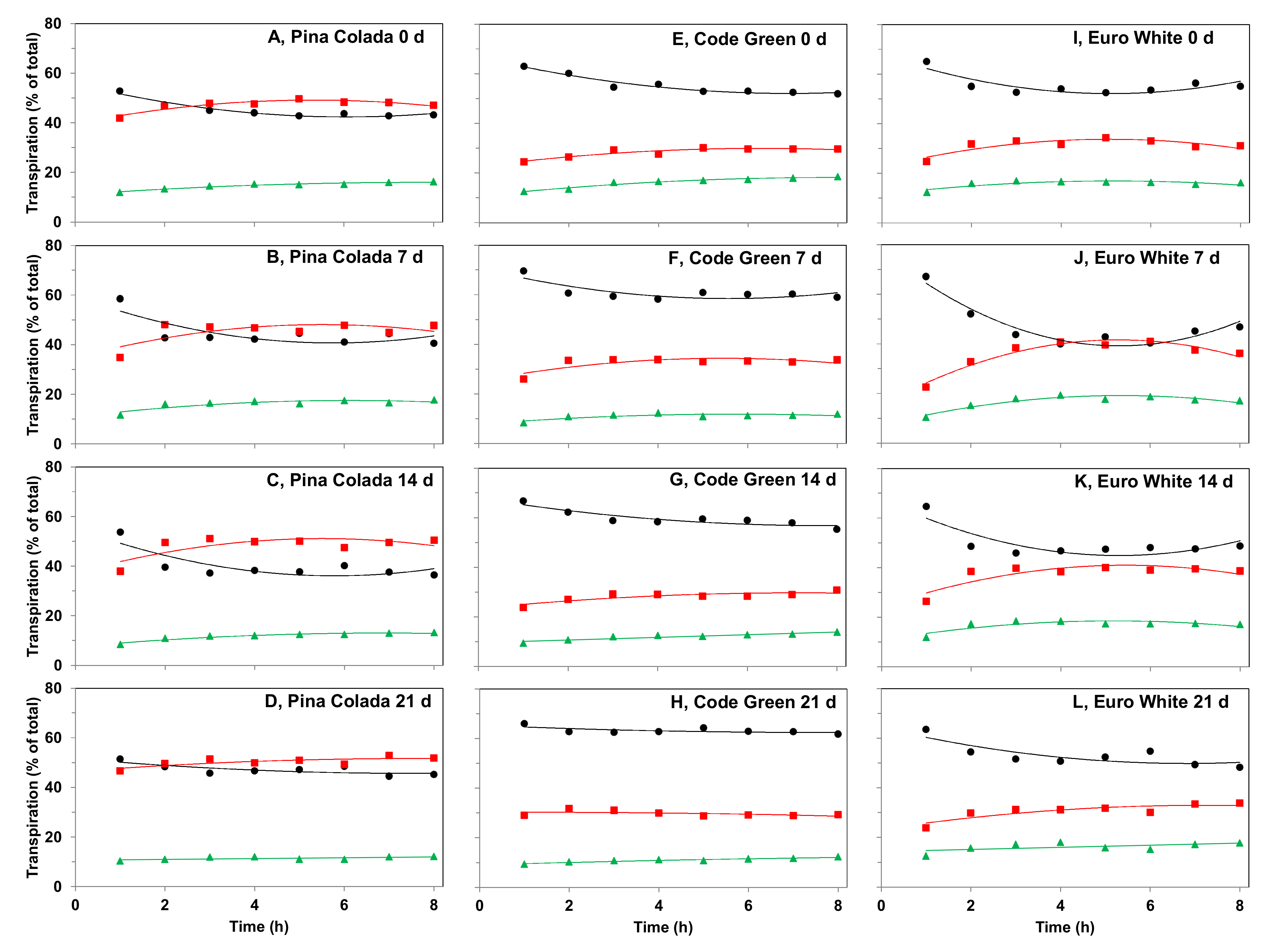
2.5.1. Effect of Storage Period on Cut Flower Transpiration Partitioning between Organs during Water Deficit
2.5.2. Stomatal Density and Dimensions on Cut Flower Organs
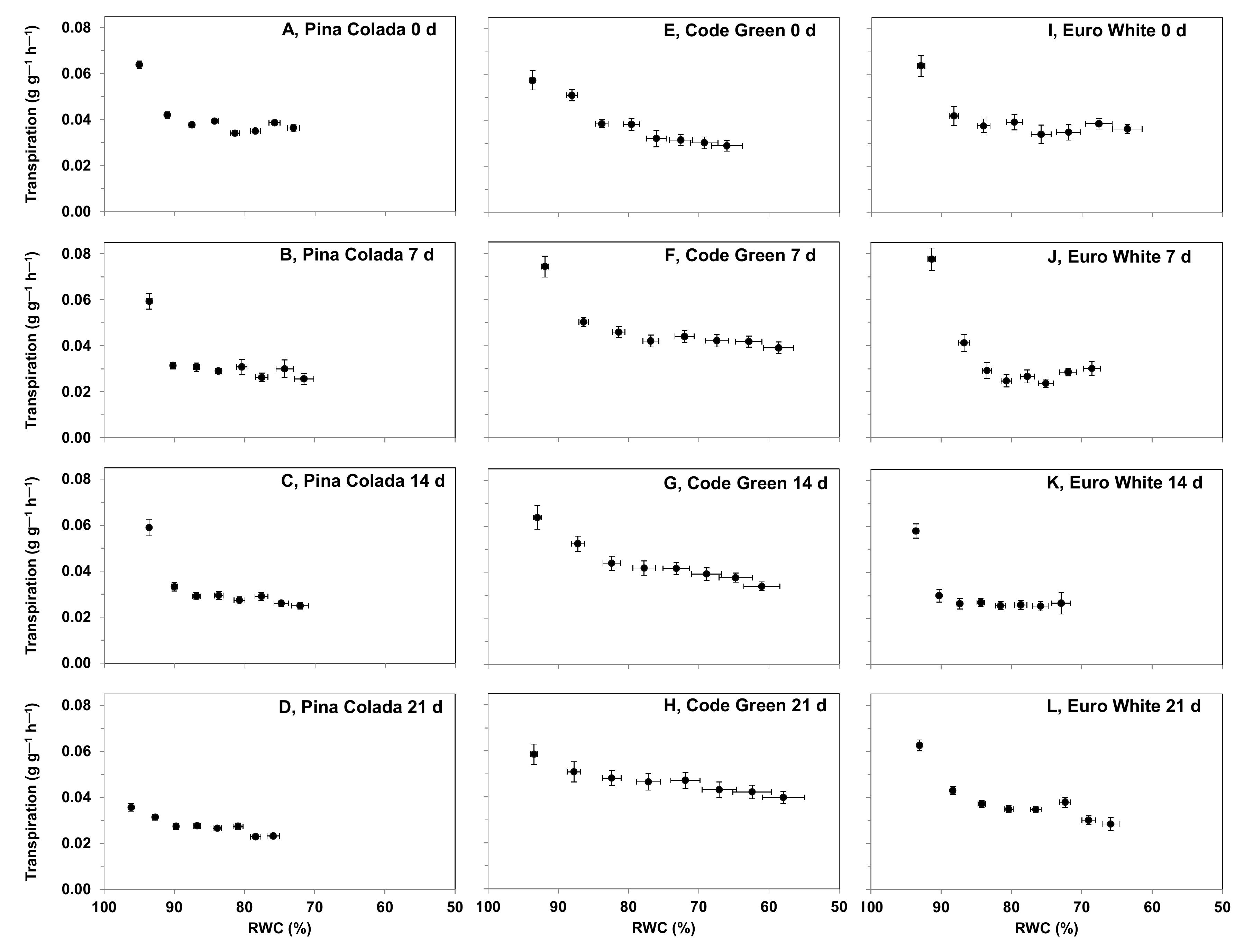
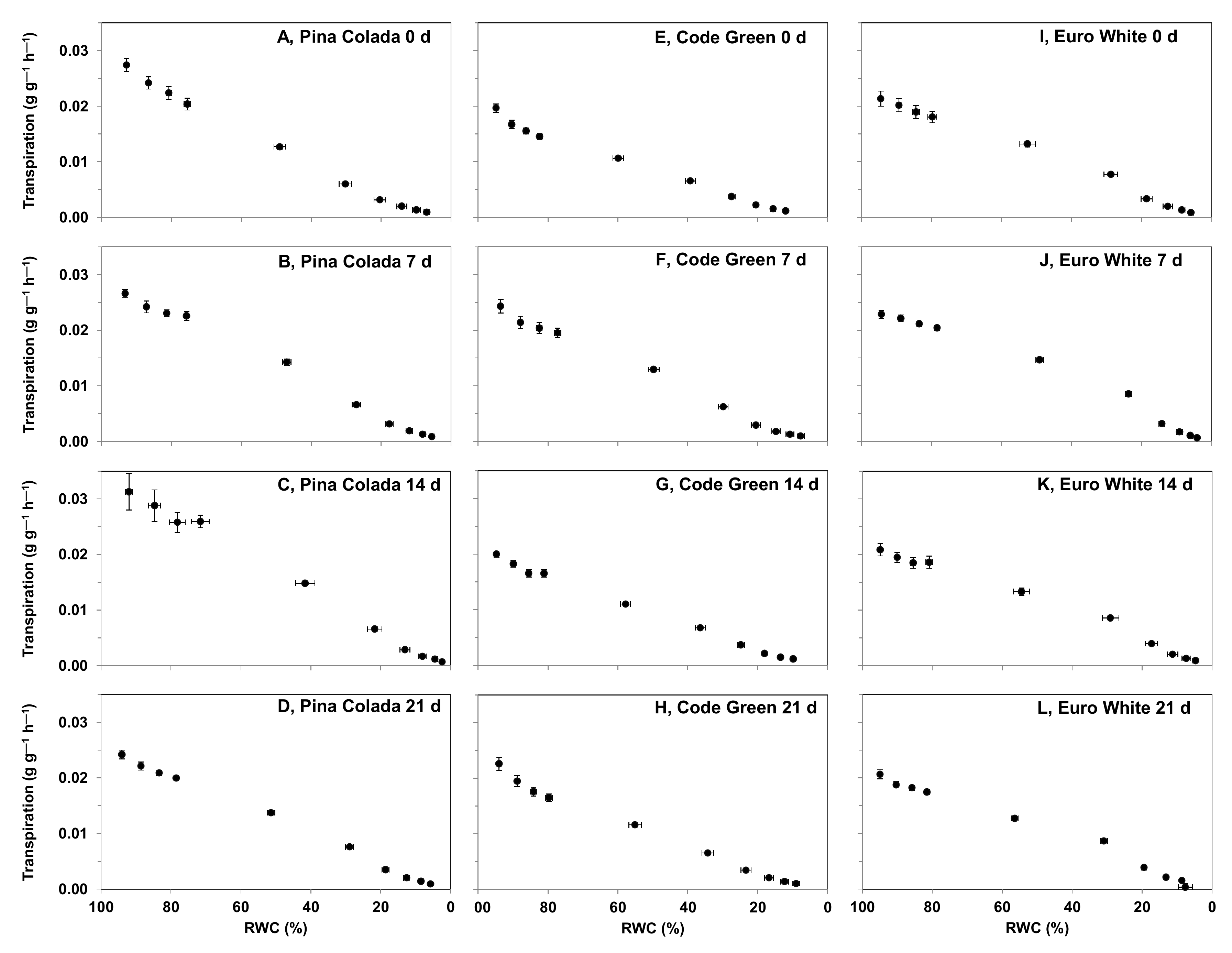
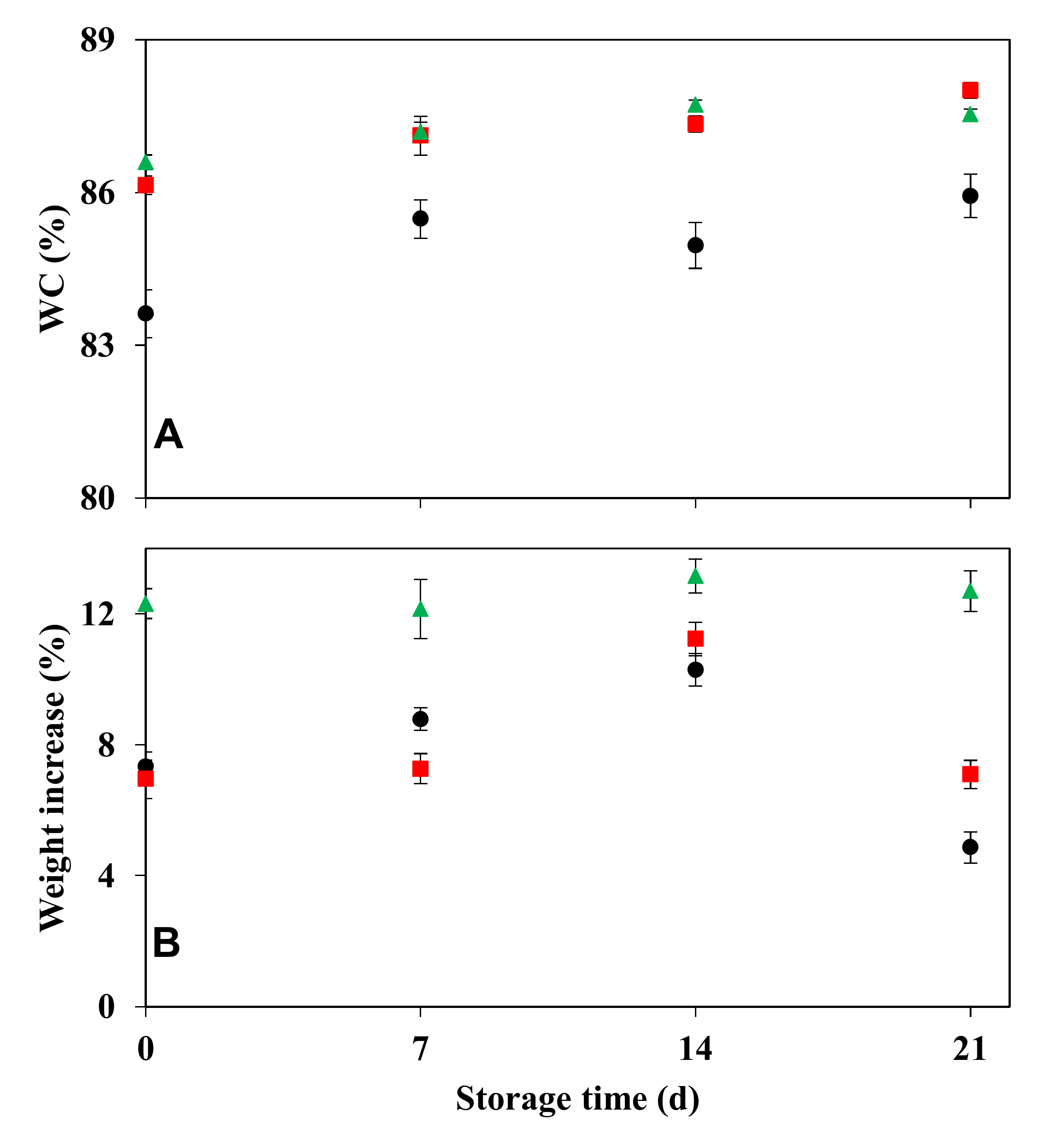
2.5.3. Effect of Storage Period on Hydration Status and Rehydration Ability following a Desiccation Event
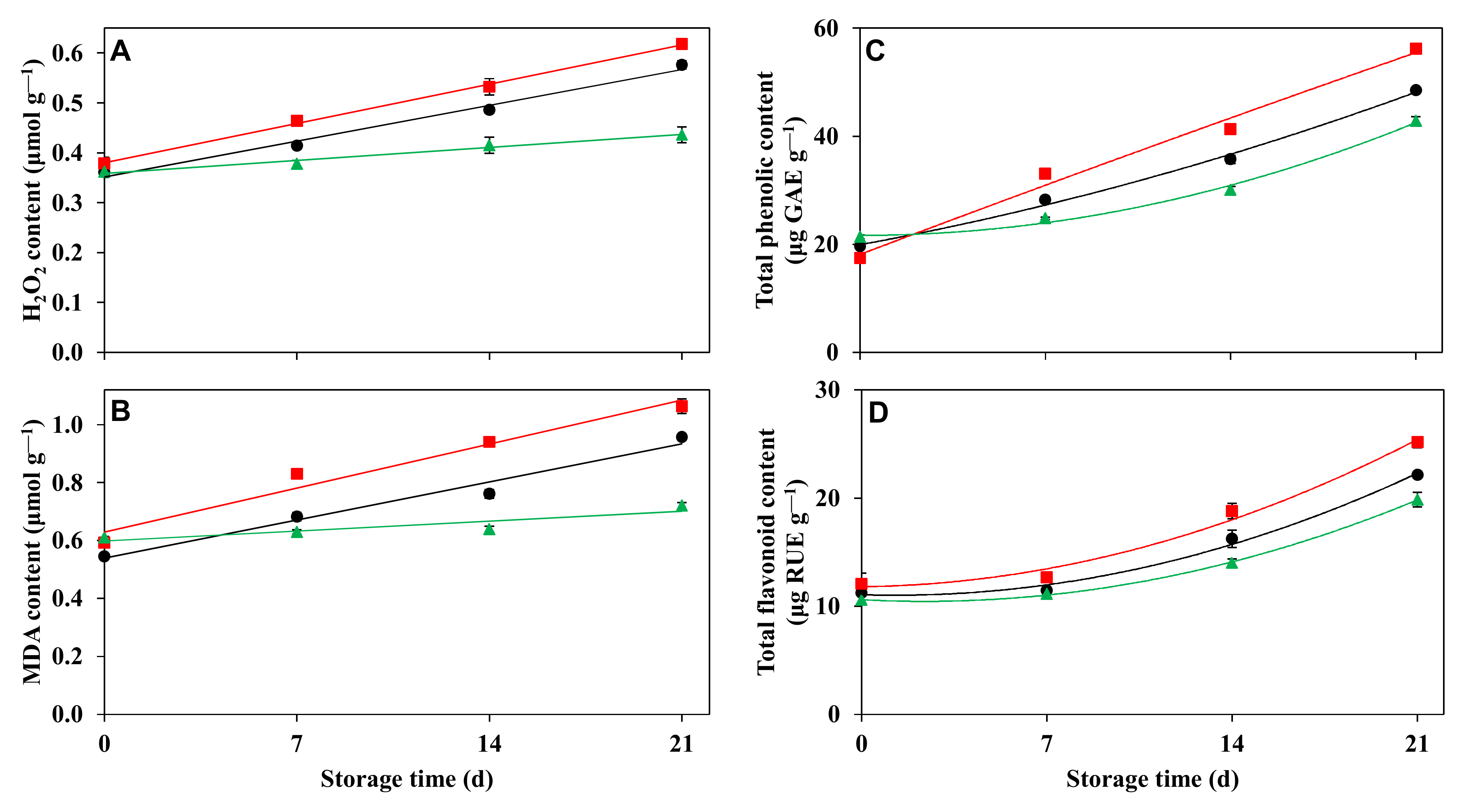
2.5.4. Effect of Storage Period on Leaf Hydrogen Peroxide Content
2.5.5. Effect of Storage Period on Lipid Peroxidation
2.5.6. Effect of Storage Period on Leaf Total Phenolic and Total Flavonoid Contents
2.5.7. Leaf and Floral Mineral Analysis
2.6. Statistical Design and Analysis
3. Results
3.1. Visual Quality Characteristics, Mass Partitioning and Stem Strength
3.2. Effect of Storage Period on Leaf and Flower Reflectance
3.3. Effect of Storage Period on Vase Life
3.4. Investigation of Cut Flower Traits Potentially Related to Vase Life Response to Storage Period
3.4.1. Effect of Storage Period on Cut Flower Transpiration Partitioning between Organs during Water Deficit
3.4.2. Stomatal Density and Dimensions on Cut Flower Organs
3.4.3. Effect of Storage Period on Hydration Status and Rehydration Ability following a Desiccation Event
3.4.4. Effect of Storage Period on Leaf Hydrogen Peroxide Content and Lipid Peroxidation
3.4.5. Effect of Storage Period on Leaf Total Phenolic and Total Flavonoid Contents
3.4.6. Leaf and Floral Mineral Analysis
4. Discussion
4.1. The Contribution of Distinct Organs to the Whole-Cut Flower Transpiration in Relation to the Presence and Functionality of Stomata
4.2. Vase Life Response to Long-Term Storage Is Related to Oxidative Stress
4.3. Leaf and Flower Reflectance Were Not Indicative of Storage Period
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Woltering, E.J. Why can’t we store flowers for longer? Opinion paper on physiological, (bio)physical and biochemical determinants of premature flower failure after long-term cold storage. Acta Hortic. 2019, 1263, 325–334. [Google Scholar] [CrossRef]
- Darras, A.I. The chilling injury effect in cut flowers: A brief review. J. Hortic. Sci. Biotechnol. 2019, 101, 1–8. [Google Scholar] [CrossRef]
- Skutnik, E.; Rabiza-Swider, J.; Jedrzejuk, A.; Łukaszewska, A. The effect of the long-term cold storage and preservatives on senescence of cut herbaceous peony flowers. Agronomy 2020, 10, 1631. [Google Scholar] [CrossRef]
- Mohammadi, M.; Aelaei, M.; Saidi, M. Pre-harvest spray of GABA and spermine delays postharvest senescence and alleviates chilling injury of gerbera cut flowers during cold storage. Sci Rep. 2021, 11, 14166. [Google Scholar] [CrossRef] [PubMed]
- Woltering, E.J.; Paillart, M.J.M. Effect of cold storage on stomatal functionality, water relations and flower performance in cut roses. Postharvest Biol. Technol. 2018, 136, 66–73. [Google Scholar] [CrossRef]
- Ahmadi-Majd, M.; Rezaei Nejad, A.; Mousavi-Fard, S.; Fanourakis, D. Deionized water as vase solution prolongs flower bud opening and vase life in cut carnation and rose through sustaining an improved water balance. Eur. J. Hortic. Sci. 2021, 86, 682–693. [Google Scholar] [CrossRef]
- Fanourakis, D.; Giday, H.; Li, T.; Kambourakis, E.; Ligoxigakis, E.K.; Papadimitriou, M.; Strataridaki, A.; Bouranis, D.; Fiorani, F.; Heuvelink, E.; et al. Antitranspirant compounds alleviate the mild-desiccation-induced reduction of vase life in cut roses. Postharvest Biol. Technol. 2016, 117, 110–117. [Google Scholar] [CrossRef]
- Fanourakis, D.; Papadopoulou, E.; Valla, A.; Tzanakakis, V.A.; Nektarios, P.A. Partitioning of transpiration to cut flower organs and its mediating role on vase life response to dry handling: A case study in chrysanthemum. Postharvest Biol. Technol. 2021, 181, 111636. [Google Scholar] [CrossRef]
- Fanourakis, D.; Hyldgaard, B.; Giday, H.; Bouranis, D.; Körner, O.; Nielsen, K.L.; Ottosen, C.O. Differential effects of elevated air humidity on stomatal closing ability of Kalanchoë blossfeldiana among the C3 and CAM states. Environ. Exp. Bot. 2017, 143, 115–124. [Google Scholar] [CrossRef]
- Fanourakis, D.; Aliniaeifard, S.; Sellin, A.; Giday, H.; Körner, O.; Rezaei Nejad, A.; Delis, C.; Bouranis, D.; Koubouris, G.; Kambourakis, E.; et al. Stomatal behavior following mid- or long term exposure to high relative air humidity: A review. Plant Physiol. Biochem. 2020, 153, 92–105. [Google Scholar] [CrossRef]
- Lin, X.; Li, H.; He, S.; Pang, Z.; Lin, S.; Li, H. Investigation of stomata in cut ‘Master’ carnations: Organographic distribution, morphology, and contribution to water loss. HortScience 2020, 55, 1144–1147. [Google Scholar] [CrossRef]
- Fanourakis, D.; Bouranis, D.; Tsaniklidis, G.; Rezaei Nejad, A.; Ottosen, C.O.; Woltering, E.J. Genotypic and phenotypic differences in fresh weight partitioning of cut rose stems: Implications for water loss. Acta Physiol. Plant 2020, 42, 48. [Google Scholar] [CrossRef]
- Xue, J.; Tang, Y.; Wang, S.; Xue, Y.; Liu, X.; Zhang, X. Evaluation of dry and wet storage on vase quality of cut peony based on the regulation of starch and sucrose metabolism. Postharvest Biol. Technol. 2019, 155, 11–19. [Google Scholar] [CrossRef]
- Aalifar, M.; Aliniaeifard, S.; Arab, M.; Mehrjerdi, M.Z.; Serek, M. Blue light postpones senescence of carnation flowers through regulation of ethylene and abscisic acid pathway-related genes. Plant Physiol. Biochem. 2020, 151, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi-Majd, M.; Rezaei Nejad, A.; Mousavi-Fard, S.; Fanourakis, D. Postharvest application of single, multi-walled carbon nanotubes and graphene oxide stimulates rose keeping quality. J. Hortic. Sci. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Horibe, T. Use of Light Stimuli as a Postharvest Technology for Cut Flowers. Front. Plant Sci. 2020, 11, 573490. [Google Scholar] [CrossRef]
- Kafkaletou, M.; Christopoulos, M.V.; Tsaniklidis, G.; Papadakis, I.; Ioannou, D.; Tzoutzoukou, C.; Tsantili, E. Nutritional value and consumer-perceived quality of fresh goji berries (Lycium barbarum L. and L. chinense L.) from plants cultivated in Southern Europe. Fruits 2018, 73, 5–12. [Google Scholar] [CrossRef]
- Pappi, P.; Nikoloudakis, N.; Fanourakis, D.; Zambounis, A.; Delis, C.; Tsaniklidis, G. Differential triggering of the phenylpropanoid biosynthetic pathway key gene expression via cold stress and viral infection in tomato leaves. Horticulturae 2021, 7, 448. [Google Scholar] [CrossRef]
- Yang, L.; Fanourakis, D.; Tsaniklidis, G.; Li, K.; Li, T. Contrary to red, blue monochromatic light improves the bioactive compound content in broccoli sprouts. Agronomy 2021, 11, 2139. [Google Scholar] [CrossRef]
- Fanourakis, D.; Velez-Ramirez, A.I.; In, B.C.; Barendse, H.; van Meeteren, U.; Woltering, E.J. A survey of preharvest conditions affecting the regulation of water loss during vase life. Acta Hortic. 2015, 1064, 195–204. [Google Scholar] [CrossRef]
- Fanourakis, D.; Carvalho, D.R.A.; Gitonga, V.W.; van Heusden, A.W.; Almeida, D.P.F.; Heuvelink, E.; Carvalho, S.M.P. Breeding cut roses for better keeping quality: First steps. Acta Hortic. 2012, 937, 875–882. [Google Scholar] [CrossRef] [Green Version]
- Caturegli, L.; Matteoli, S.; Gaetani, M.; Grossi, N.; Magni, S.; Minelli, A.; Corsini, G.; Remorini, D.; Volterrani, M. Effects of water stress on spectral reflectance of bermudagrass. Sci. Rep. 2020, 10, 15055. [Google Scholar] [CrossRef]
- Cundill, S.L.; Van der Werff, H.M.A.; Van der Meijde, M. Adjusting Spectral Indices for Spectral Response Function Differences of Very High Spatial Resolution Sensors Simulated from Field Spectra. Sensors 2015, 15, 6221–6240. [Google Scholar] [CrossRef] [PubMed]
- El-Hendawy, S.; Al-Suhaibani, N.; Hassan, W.; Tahir, M.; Schmidhalter, U. Hyperspectral reflectance sensing to assess the growth and photosynthetic properties of wheat cultivars exposed to different irrigation rates in an irrigated arid region. PLoS ONE 2017, 12, e0183262. [Google Scholar] [CrossRef]
- Zhu, H.; Chu, B.; Fan, Y.; Tao, X.; Yin, W.; He, Y. Hyperspectral Imaging for Predicting the Internal Quality of Kiwifruits Based on Variable Selection Algorithms and Chemometric Models. Sci. Rep. 2017, 7, 7845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhry, M.M.A.; Amodio, M.L.; Babellahi, F.; de Chiara, M.L.V.; Amigo Rubio, J.M.; Colelli, G. Hyperspectral imaging and multivariate accelerated shelf life testing (MASLT) approach for determining shelf life of rocket leaves. J. Food Eng. 2018, 238, 122–133. [Google Scholar] [CrossRef]
- Van Meeteren, U.; van Gelder, H.; van Ieperen, W. Reconsideration of the use of deionized water as vase water in postharvest experiments on cut flowers. Postharvest Biol. Technol. 1999, 18, 169–181. [Google Scholar] [CrossRef]
- Fanourakis, D.; Kazakos, P.; Nektarios, P.A. Allometric individual leaf area estimation in chrysanthemum. Agronomy 2021, 11, 795. [Google Scholar] [CrossRef]
- Körner, O.; Fanourakis, D.; Hwang, M.C.R.; Hyldgaard, B.; Tsaniklidis, G.; Nikoloudakis, N.; Larsen, D.H.; Ottosen, C.O.; Rosenqvist, E. Incorporating cultivar-specific stomatal traits into stomatal conductance models improves the estimation of evapotranspiration enhancing greenhouse climate management. Biosyst. Eng. 2021, 208, 131–151. [Google Scholar] [CrossRef]
- Koubouris, G.; Bouranis, D.; Vogiatzis, E.; Rezaei Nejad, A.; Giday, H.; Tsaniklidis, G.; Ligoxigakis, E.K.; Blazakis, K.; Kalaitzis, P.; Fanourakis, D. Leaf area estimation by considering leaf dimensions in olive tree. Sci. Hortic. 2018, 240, 440–445. [Google Scholar] [CrossRef]
- Gupta, S.; Rosenthal, D.M.; Stinchcombe, J.R.; Baucom, R.S. The remarkable morphological diversity of leaf shape in sweet potato (Ipomoea batatas): The influence of genetics, environment, and G9E. New Phytol. 2020, 225, 2183–2195. [Google Scholar] [CrossRef]
- Papadakis, V.M.; Orphanos, Y.; Kogou, S.; Melessanaki, K.; Pouli, P.; Fotakis, C. IRIS: A novel spectral imaging system for the analysis of cultural heritage objects. In O3A: Optics for Arts, Architecture, and Archaeology III; International Society for Optics and Photonics: Bellingham, WA, USA, 2011. [Google Scholar]
- Zacharopoulos, K.; Hatzigiannakis, P.; Karamaoynas, V.; Papadakis, M.; Andrianakis, M.; Melessanaki, K.; Zabulis, X. A method for the registration of spectral images of paintings and its evaluation. J. Cult. Herit. 2017, 29, 10–19. [Google Scholar] [CrossRef]
- VBN. Evaluation Cards for Cut Flowers; VBN (Association of Flower Auctions in The Netherlands): Leiden, The Netherlands, 2005. [Google Scholar]
- Fanourakis, D.; Giday, H.; Hyldgaard, B.; Bouranis, D.; Körner, O.; Ottosen, C.O. Low air humidity during growth promotes stomatal closure ability in roses. Eur. J. Hortic. Sci. 2019, 84, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Fanourakis, D.; Hyldgaard, B.; Giday, H.; Aulik, I.; Bouranis, D.; Körner, O.; Ottosen, C.O. Stomatal anatomy and closing ability is affected by supplementary light intensity in rose (Rosa hybrida L.). Hort Sci. 2019, 46, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Seif, M.; Aliniaeifard, S.; Arab, M.; Mehrjerdi, M.Z.; Shomali, A.; Fanourakis, D.; Li, T.; Woltering, E. Monochromatic red light during plant growth decreases the size and improves the functionality of stomata in chrysanthemum. Funct. Plant Biol. 2021, 48, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.R.A.; Fanourakis, D.; Correia, M.J.; Monteiro, J.A.; Araújo-Alves, J.P.L.; Vasconcelos, M.W.; Almeida, D.P.F.; Heuvelink, E.; Carvalho, S.M.P. Root-to-shoot ABA signaling does not contribute to genotypic variation in stomatal functioning induced by high relative air humidity. Environ. Exp. Bot. 2016, 123, 13–21. [Google Scholar] [CrossRef]
- Taheri-Garavand, A.; Rezaei Nejad, A.; Fanourakis, D.; Fatahi, S.; Ahmadi-Majd, M. Employment of artificial neural networks for non-invasive estimation of leaf water status using color features: A case study in Spathiphyllum wallisii. Acta Physiol. Plant. 2021, 43, 78. [Google Scholar] [CrossRef]
- Fanourakis, D.; Bouranis, D.; Giday, H.; Carvalho, D.R.A.; Rezaei Nejad, A.; Ottosen, C.O. Improving stomatal functioning at elevated growth air humidity: A review. J. Plant Phys. 2016, 207, 51–60. [Google Scholar] [CrossRef]
- Fanourakis, D.; Heuvelink, E.; Carvalho, S.M.P. Spatial heterogeneity in stomatal features during leaf elongation: An analysis using Rosa hybrida. Funct. Plant Biol. 2015, 42, 737–745. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, H.K.; Fanourakis, D.; Tsaniklidis, G.; Bouranis, D.; Rezaei Nejad, A.; Ottosen, C.O. Using artificial lighting based on electricity price without a negative impact on growth, visual quality or stomatal closing response in Passiflora. Sci. Hortic. 2020, 267, 109354. [Google Scholar] [CrossRef]
- Tsaniklidis, G.; Pappi, P.; Tsafouros, A.; Charova, S.N.; Nikoloudakis, N.; Roussos, P.A.; Paschalidis, K.A.; Delis, C. Polyamine homeostasis in tomato biotic/abiotic stress cross-tolerance. Gene 2020, 727, 144230. [Google Scholar] [CrossRef] [PubMed]
- Gerasopoulos, D.; Chebli, B. Effects of pre- and postharvest calcium applications on the vase-life of cut gerberas. J. Hortic. Sci. Biotechnol. 1999, 74, 78–81. [Google Scholar] [CrossRef]
- Torre, S.; Borochov, A.; Halevy, A.H. Calcium regulation of senescence in rose petals. Physiol. Plant. 1999, 107, 214–219. [Google Scholar] [CrossRef]
- Fanourakis, D.; Pieruschka, R.; Savvides, A.; Macnish, A.J.; Sarlikioti, V.; Woltering, E.J. Sources of vase life variation in cut roses: A review. Postharvest Biol. Technol. 2013, 78, 1–15. [Google Scholar] [CrossRef]
- Chatzistathis, T.; Fanourakis, D.; Aliniaeifard, S.; Kotsiras, A.; Delis, C.; Tsaniklidis, G. Leaf age-dependent effects of boron toxicity in two Cucumis melo varieties. Agronomy 2021, 11, 759. [Google Scholar] [CrossRef]
- Jones, J. Laboratory Guide for Conducting Soil Tests and Plant Analysis; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Kuo, S. Phosphorus. In Methods of Soil Analysis. Part 3: Chemical Methods; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Horneck, D.; Hanson, D. Determination of Potassium and Sodium by Flame Emission Spectrophotometry. In Handbook of Reference Methods for Plant Analysis; CRC Press: Boca Raton, FL, USA, 1998; pp. 153–155. [Google Scholar]
- Hanlon, E.A. Elemental Determination by Atomic Absorption Spectrometry. In Handbook of Reference Methods for Plant Analysis; CRC Press: Boca Raton, FL, USA, 1997; pp. 157–164. [Google Scholar]
- Van Doorn, W.G. Water relations of cut flowers. Hortic. Rev. 1997, 18, 1–85. [Google Scholar]
- Carvalho, D.R.A.; Koning-Boucoiran, C.F.S.; Fanourakis, D.; Vasconcelos, M.W.; Carvalho, S.M.P.; Heuvelink, E.; Krens, F.A.; Maliepaard, C. QTL analysis for stomatal functioning in tetraploid Rosa x hybrida grown at high relative air humidity and its implications on postharvest longevity. Mol. Breed. 2015, 35, 172. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index Name (Abbreviation) | Equation | Reference |
|---|---|---|
| Photochemical reflectance index (PRI) | [24] | |
| Renormalized difference vegetation index (RDVI) | [23] | |
| Water index (WI) | [22] | |
| Normalized difference vegetation index (NDVI) | [22] | |
| Ratio of WI with NDVI (WI:NDVI) | [22] | |
| Normalized water index (NWI) | [24] | |
| Optimized soil-adjusted vegetation index (OSAVI) | [23] | |
| Structure independent pigment index (SIPI) | [23] | |
| Blue/green index 2 (BGI2) | [23] | |
| Triangular vegetation index (TVI) | )] | [23] |
| Modified chlorophyll absorption in reflectance index (MCARI) | ) | [23] |
| Cultivar | Leaf | Flower | Leaf Weight (%) | Flower Weight (%) | Stem Weight (%) | Stem Strength (g m−1) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Aspect Ratio | Circularity | Roundness | Solidity | Number | Distance (cm) | Diameter (cm) | |||||
| ‘Pina Colada’ | 1.85 ± 0.03 b | 0.15 ± 0.00 b | 0.54 ± 0.01 b | 0.67 ± 0.00 b | 35.9 ± 0.8 a | 21.8 ± 0.9 a | 8.4 ± 0.2 a | 30.1 ± 0.7 b | 29.9 ± 0.5 a | 40.0 ± 0.5 | 20 ± 0 b |
| ‘Code Green’ | 1.63 ± 0.04 c | 0.19 ± 0.00 a | 0.62 ± 0.02 a | 0.72 ± 0.00 a | 16.9 ± 0.8 b | 16.4 ± 1.1 b | 3.6 ± 0.1 c | 33.5 ± 1.4 ab | 28.4 ± 1.0 ab | 38.2 ± 0.8 | 27 ± 1 a |
| ‘Euro White’ | 2.21 ± 0.03 a | 0.12 ± 0.00 c | 0.46 ± 0.01 c | 0.62 ± 0.01 c | 12.6 ± 0.5 c | 10.9 ± 0.7 c | 7.6 ± 0.1 b | 34.6 ± 0.9 a | 26.1 ± 1.0 b | 39.4 ± 0.9 | 30 ± 1 a |
| n | 60 | 60 | 60 | 60 | 40 | 12 | 12 | 12 | 12 | 12 | 12 |
| p | <0.00001 | <0.00001 | <0.00001 | <0.00001 | <0.00001 | <0.00001 | <0.00001 | 0.011487 | 0.013213 | 0.220553 | <0.00001 |
| Cultivar | Fresh Weight (g) | Number of Leaves | Leaf Area (cm2) | Stem Area (cm2) | Flower Area (cm2) |
|---|---|---|---|---|---|
| ‘Pina Colada’ | 98.2 ± 1.7 a | 38.4 ± 1.2 a | 637 ± 24 a | 199 ± 5 a | 387 ± 17 a |
| ‘Code Green’ | 84.8 ± 1.9 b | 15.0 ± 0.5 b | 543 ± 20 b | 149 ± 6 b | 111 ± 5 c |
| ‘Euro White’ | 94.6 ± 1.6 a | 10.4 ± 0.4 c | 610 ± 20 ab | 149 ± 4 b | 162 ± 7 b |
| p | <0.00001 | <0.00001 | <0.00001 | <0.00001 | <0.00001 |
| Cultivar | Storage Period (d) | Cumulative Transpiration (g g−1) | Decrease in Transpiration (%) | RWC (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cut Flower | Leaf | Stem | Flower | Cut Flower | Leaf | Stem | Flower | Leaf | ||
| ‘Pina Colada’ | 0 | 0.164 | 0.248 ± 0.008 de | 0.189 ± 0.009 ab | 0.079 ± 0.004 d | 33.7 | 45.7 ± 3.2 ab | 25.8 ± 1.3 | 9.1 ± 1.5 ab | 75.3 ± 0.9 |
| ‘Code Green’ | 0.184 | 0.309 ± 0.019 acd | 0.133 ± 0.005 abc | 0.103 ± 0.007 abc | 38.7 | 49.4 ± 1.7 ab | 25.6 ± 1.9 | 8.7 ± 1.5 ab | 69.6 ± 1.9 | |
| ‘Euro White’ | 0.202 | 0.327 ± 0.018 ab | 0.157 ± 0.009 bcd | 0.121 ± 0.002 a | 32.8 | 40.8 ± 4.2 ab | 15.0 ± 1.7 | 12.2 ± 1.0 ab | 70.0 ± 3.1 | |
| ‘Pina Colada’ | 7 | 0.174 | 0.263 ± 0.013 cd | 0.193 ± 0.005 ab | 0.092 ± 0.008 bc | 38.0 | 53.8 ± 6.3 bc | 14.7 ± 3.0 | 3.8 ± 1.6 ab | 76.9 ± 1.2 |
| ‘Code Green’ | 0.206 | 0.378 ± 0.019 a | 0.171 ± 0.008 a | 0.079 ± 0.002 d | 38.3 | 46.0 ± 4.2 ab | 19.3 ± 1.5 | 10.8 ± 3.1 ab | 68.9 ± 3.2 | |
| ‘Euro White’ | 0.198 | 0.282 ± 0.011 bcd | 0.173 ± 0.004 bcd | 0.122 ± 0.003 a | 44.4 | 57.0 ± 7.3 c | 10.1 ± 2.1 | 8.2 ± 1.1 ab | 76.3 ± 1.8 | |
| ‘Pina Colada’ | 14 | 0.190 | 0.258 ± 0.011 cd | 0.224 ± 0.018 a | 0.073 ± 0.004 de | 37.6 | 56.2 ± 3.0 c | 13.4 ± 4.0 | 2.0 ± 1.2 a | 77.2 ± 1.5 |
| ‘Code Green’ | 0.197 | 0.354 ± 0.023 ab | 0.143 ± 0.005 ab | 0.083 ± 0.004 cd | 36.0 | 45.0 ± 2.8 ab | 17.3 ± 2.3 | 5.4 ± 1.5 ab | 66.0 ± 3.7 | |
| ‘Euro White’ | 0.167 | 0.242 ± 0.011 de | 0.155 ± 0.008 bcd | 0.106 ± 0.004 ab | 39.0 | 57.0 ± 5.5 c | 10.8 ± 1.8 | 12.1 ± 1.0 ab | 77.5 ± 1.3 | |
| ‘Pina Colada’ | 21 | 0.141 | 0.222 ± 0.008 e | 0.174 ± 0.005 bcd | 0.054 ± 0.004 e | 25.8 | 33.9 ± 3.5 ab | 17.1 ± 1.5 | 13.8 ± 5.4 bc | 79.4 ± 0.8 |
| ‘Code Green’ | 0.199 | 0.376 ± 0.028 a | 0.152 ± 0.007 a | 0.076 ± 0.003 d | 27.6 | 31.2 ± 2.2 a | 26.4 ± 1.5 | 4.8 ± 1.0 ab | 63.1 ± 3.4 | |
| ‘Euro White’ | 0.198 | 0.308 ± 0.010 abc | 0.150 ± 0.004 de | 0.121 ± 0.007 a | 40.4 | 55.2 ± 4.3 c | 15.0 ± 1.9 | 15.7 ± 2.0 c | 72.7 ± 1.5 | |
| p | ||||||||||
| Cultivar | <0.001 | <0.001 | <0.001 | 0.009 | <0.001 | 0.003 | <0.001 | |||
| Storage period | 0.341 | 0.004 | <0.001 | 0.001 | <0.001 | 0.029 | 0.444 | |||
| Cultivar × Storage period | <0.001 | 0.002 | <0.001 | 0.007 | 0.272 | 0.005 | 0.093 | |||
| Cultivar | Adaxial Stomatal | Abaxial Stomatal | Leaf Stomatal Density (mm−2) | Stomata on the Abaxial Side (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Density (mm−2) | Length (μm) | Width (μm) | Size (μm2) | Length to Width | Density (mm−2) | Length (μm) | Width (μm) | Size (μm2) | Length to Width | |||
| ‘Pina Colada’ | 10.0 ± 0.6 a | 58.8 ± 2.1 b | 32.2 ± 1.0 b | 1903 ± 125 b | 1.83 ± 0.04 a | 32.9 ± 1.1 b | 56.9 ± 2.8 b | 38.4 ± 1.2 b | 2192 ± 166 b | 1.49 ± 0.04 | 42.9 ± 1.2 | 76.8 ± 1.3 b |
| ‘Code Green’ | 10.4 ± 0.5 a | 63.6 ± 0.8 a | 37.4 ± 0.8 a | 2388 ± 79 a | 1.72 ± 0.03 ab | 28.8 ± 0.4 b | 57.1 ± 1.1 b | 37.9 ± 0.8 b | 2167 ± 55 b | 1.51 ± 0.05 | 39.3 ± 0.7 | 73.4 ± 1.5 b |
| ‘Euro White’ | 5.9 ± 0.4 b | 57.3 ± 1.2 b | 34.7 ± 0.8 ab | 1990 ± 77 b | 1.66 ± 0.03 b | 37.1 ± 0.8 a | 63.8 ± 1.1 a | 42.7 ± 1.2 a | 2731 ± 91 a | 1.50 ± 0.05 | 43.0 ± 0.8 | 86.0 ± 0.9 a |
| n | 8 | 4 | 4 | 4 | 4 | 8 | 4 | 4 | 4 | 4 | 8 | 8 |
| p | <0.00001 | 0.003543 | 0.000482 | 0.000579 | 0.020969 | 0.000071 | 0.001509 | 0.000626 | 0.000035 | 0.886066 | 0.086436 | <0.00001 |
| Cultivar | Stomatal | ||||
|---|---|---|---|---|---|
| Density (mm−2) | Length (μm) | Width (μm) | Size (μm2) | Length to Width | |
| ‘Pina Colada’ | 2.2 ± 0.7 | 49.6 ± 2.6 | 24.0 ± 1.1 | 1194 ± 101 | 2.08 ± 0.12 a |
| ‘Code Green’ | 3.7 ± 0.8 | 47.0 ± 3.0 | 30.8 ± 2.5 | 1476 ± 220 | 1.55 ± 0.08 b |
| ‘Euro White’ | 1.6 ± 0.7 | 50.4 ± 3.9 | 28.9 ± 2.2 | 1484 ± 205 | 1.75 ± 0.10 ab |
| n | 8 | 4 | 4 | 4 | 4 |
| p | 0.123582 | 0.742849 | 0.081331 | 0.80645 | 0.007452 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanourakis, D.; Papadakis, V.M.; Psyllakis, E.; Tzanakakis, V.A.; Nektarios, P.A. The Role of Water Relations and Oxidative Stress in the Vase Life Response to Prolonged Storage: A Case Study in Chrysanthemum. Agriculture 2022, 12, 185. https://doi.org/10.3390/agriculture12020185
Fanourakis D, Papadakis VM, Psyllakis E, Tzanakakis VA, Nektarios PA. The Role of Water Relations and Oxidative Stress in the Vase Life Response to Prolonged Storage: A Case Study in Chrysanthemum. Agriculture. 2022; 12(2):185. https://doi.org/10.3390/agriculture12020185
Chicago/Turabian StyleFanourakis, Dimitrios, Vassilis M. Papadakis, Evangelos Psyllakis, Vasileios A. Tzanakakis, and Panayiotis A. Nektarios. 2022. "The Role of Water Relations and Oxidative Stress in the Vase Life Response to Prolonged Storage: A Case Study in Chrysanthemum" Agriculture 12, no. 2: 185. https://doi.org/10.3390/agriculture12020185