
An Eleven-Year Survey on Field Disease Susceptibility of Citrus Accessions to Colletotrichum and Alternaria Species

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Germplasm, Timing, and Location of Phytopatological Survey
2.2. Description of Disease Symptoms
2.3. Isolation and Fungal Identification
2.4. Susceptibility Evaluation and Risk of Disease Outbreaks
3. Results and Discussion
3.1. Disease Symptoms
3.2. Isolation and Fungal Identification
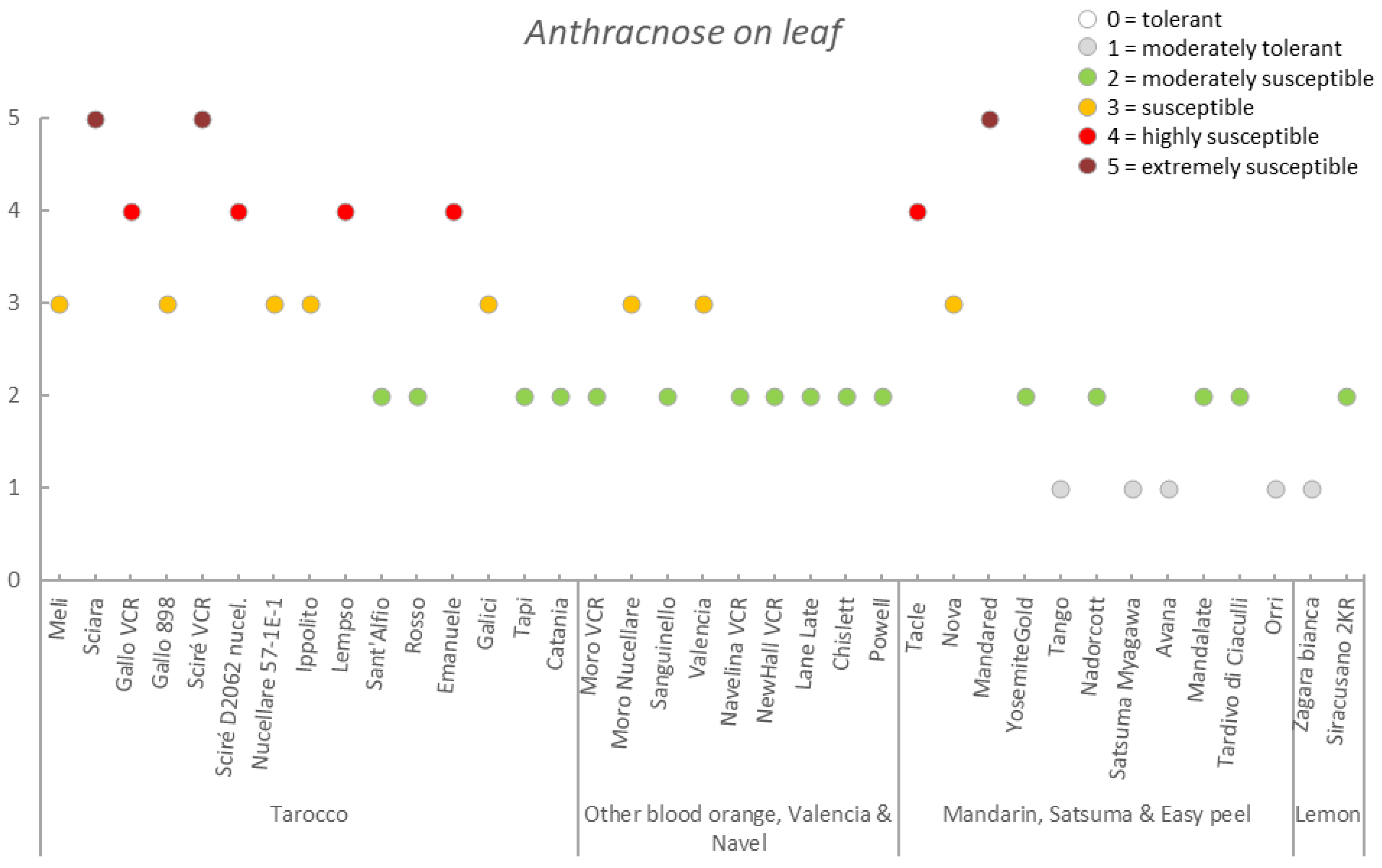
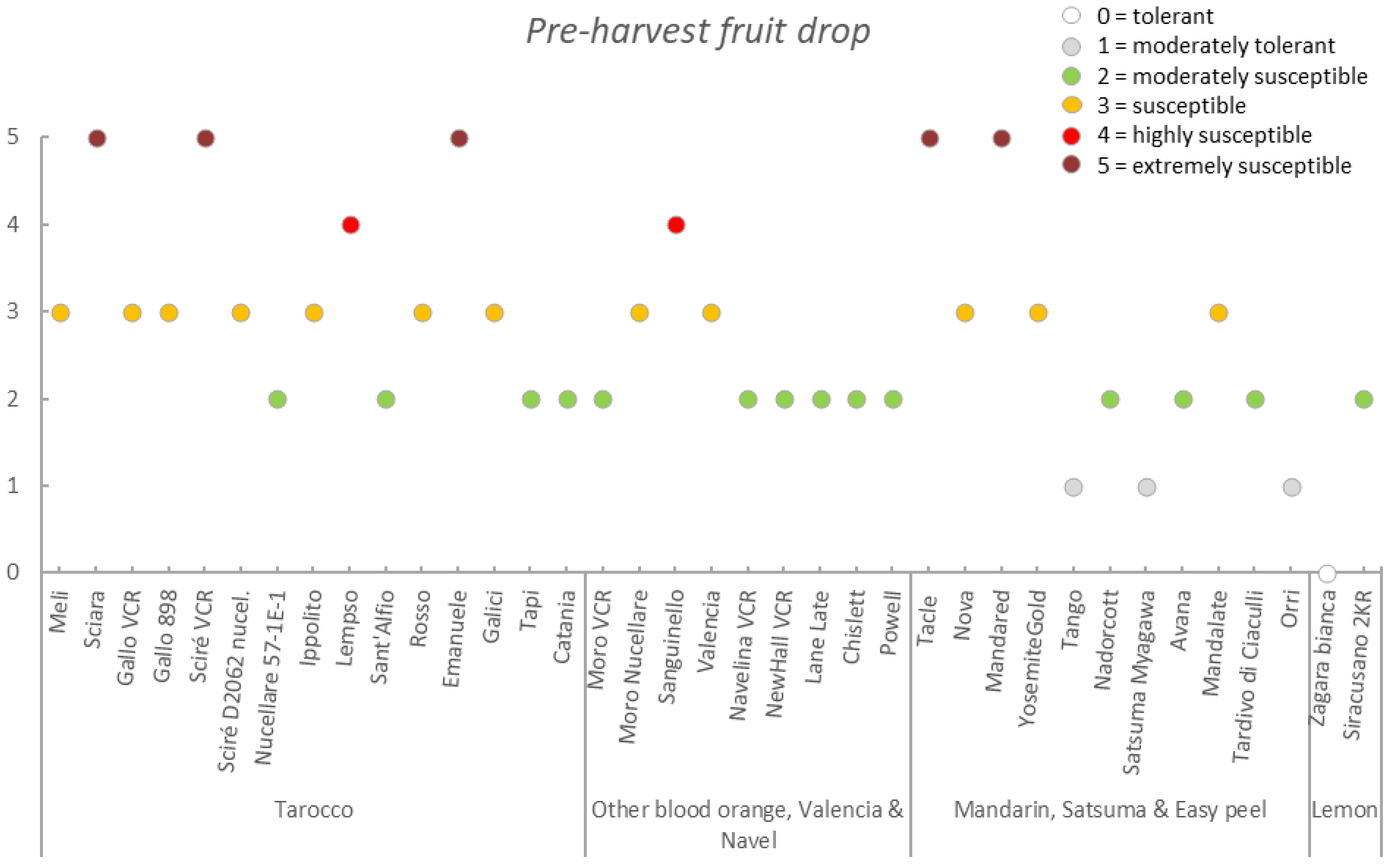
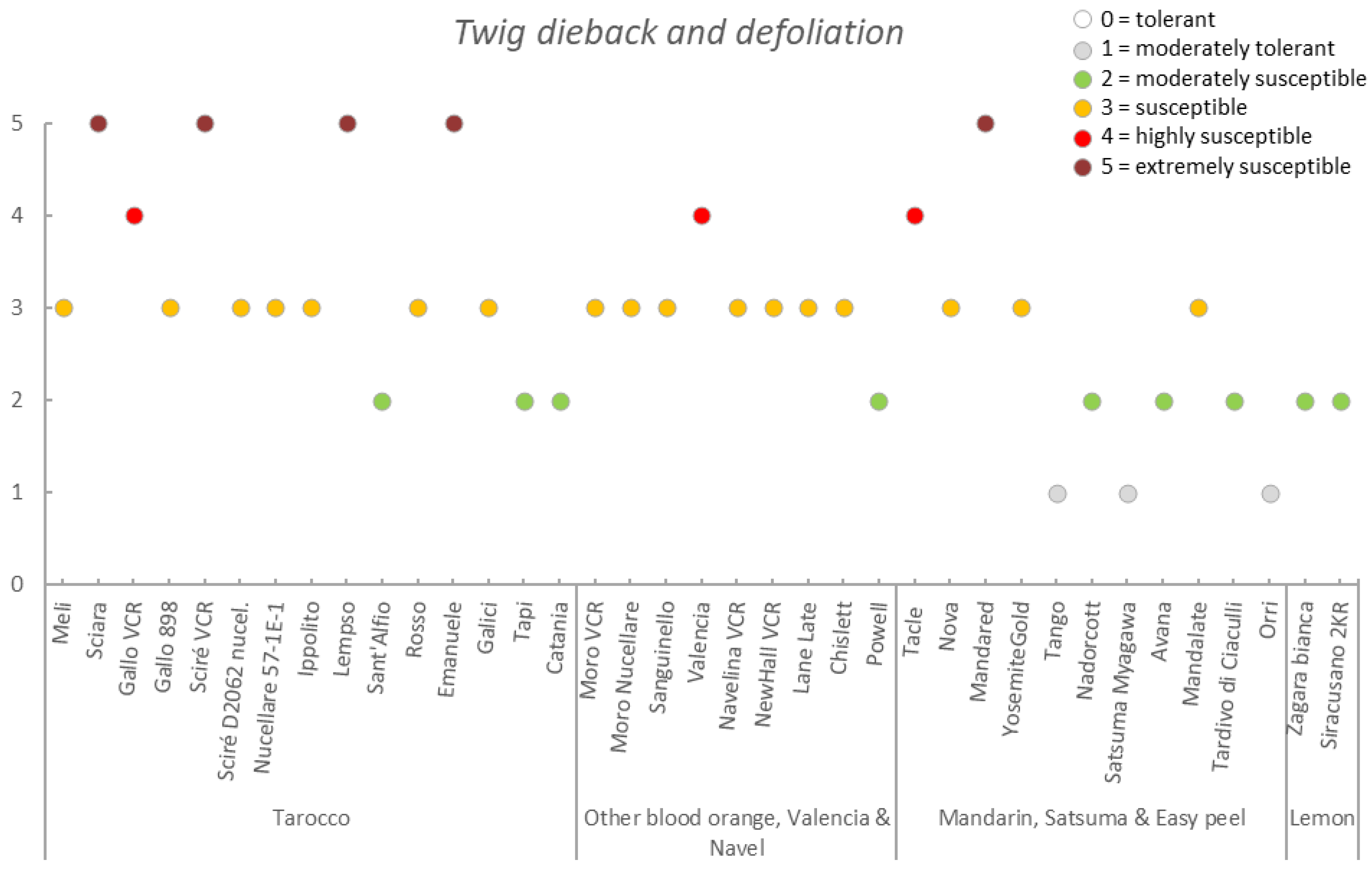
3.3. Germplasm Susceptibility Evaluation
3.4. Risk Assessment of Epidemic Outbreaks
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. 2021. Available online: http://www.fao.org/faostat/en/#home (accessed on 24 November 2020).
- Timmer, L.W.; Solel, Z.; Orozco-Santos, M. Alternaria brown spot of mandarins. In Compendium of Citrus Diseases; Timmer, L.W., Garnsey, S.M., Graham, J.H., Eds.; APS Press, Inc.: St. Paul, MN, USA, 2000; pp. 19–21. [Google Scholar]
- Lima, W.G.; Spósito, M.B.; Amorim, L.; Gonçalves, F.P.; Melo de Filho, P.A. Colletotrichum gloeosporioides, a new causal agent of citrus post-bloom fruit drop. Eur. J. Plant Pathol. 2011, 131, 157–165. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Crous, P.W. Species of Diaporthe on Camellia and Citrus in the Azores Islands. Phytopathol. Mediterr. 2018, 57, 307–319. [Google Scholar]
- Aiello, D.; Carrieri, R.; Guarnaccia, V.; Vitale, A.; Lahoz, E.; Polizzi, G. Characterization and pathogenicity of Colletotrichum gloeosporioides and C. karstii causing pre-harvest disease on Citrus sinensis in Italy. J. Phytopathol. 2015, 163, 168–177. [Google Scholar] [CrossRef]
- Aiello, D.; Guarnaccia, V.; Azzaro, A.; Polizzi, G. Alternaria brown spot on new clones of sweet orange and lemon in Italy. Phytopathol. Mediterr. 2020, 59, 131–145. [Google Scholar]
- Guarnaccia, V.; Van Niekerk, J.; Crous, P.W.; Sandoval-Denis, M. Neocosmospora spp. associated with dry root rot of citrus in South Africa. Phytopathol. Mediterr. 2021, 60, 81–102. [Google Scholar] [CrossRef]
- Wang, W.; de Silva, D.D.; Moslemi, A.; Edwards, J.; Ades, P.K.; Crous, P.W.; Taylor, P.W.J. Colletotrichum Species Causing Anthracnose of Citrus in Australia. J. Fungi 2021, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Guarnaccia, V.; Groenewald, J.Z.; Polizzi, G.; Crous, P.W. High Species Diversity in Colletotrichum Associated with Citrus Diseases in Europe. Pers. Mol. Phylogeny Evol. Fungi 2017, 39, 32–50. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Gehrmann, T.; Silva-Junior, G.J.; Fourie, P.H.; Haridas, S.; Vu, D.; Spatafora, J.; Martin, F.M.; Robert, V.; Grigoriev, I.V.; et al. Phyllosticta citricarpa and sister species of global importance to Citrus. Mol. Plant Pathol. 2019, 20, 1619–1635. [Google Scholar] [CrossRef] [Green Version]
- Juroszek, P.; Racca, P.; Link, S.; Farhumand, J.; Kleinhenz, B. Overview on the review articles published during the past 30 years relating to the potential climate change effects on plant pathogens and crop disease risks. Plant Pathol. 2020, 69, 179–193. [Google Scholar] [CrossRef]
- Möller, M.; Stukenbrock, E.H. Evolution and genome architecture in fungal plant pathogens. Nat. Rev. Microbiol. 2017, 15, 756–771. [Google Scholar] [CrossRef]
- Bezerra, J.D.P.; Crous, P.W.; Aiello, D.; Gullino, M.L.; Polizzi, G.; Guarnaccia, V. Genetic Diversity and Pathogenicity of Botryosphaeriaceae Species Associated with Symptomatic Citrus Plants in Europe. Plants 2021, 10, 492. [Google Scholar] [CrossRef]
- Rhaiem, A.; Taylor, P.W.J. Colletotrichum gloeosporioides associated with anthracnose symptoms on citrus, a new report for Tunisia. Eur. J. Plant Pathol. 2016, 146, 219–224. [Google Scholar] [CrossRef]
- Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Eskalen, A. Identification, distribution, and pathogenicity of Diatrypaceae and Botryosphaeriaceae associated with citrus branch canker in the Southern California desert. Plant Dis. 2016, 100, 2402–2413. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Aragón, D.; Silva-Rojas, H.V.; Guarnaccia, V.; Mora-Aguilera, J.A.; Aranda-Ocampo, S.; Bautista-Martínez, N.; Téliz-Ortíz, D. Colletotrichum species causing anthracnose on avocado fruit in Mexico: Current status. Plant Pathol. 2020, 69, 1513–1528. [Google Scholar] [CrossRef]
- Khodadadi, F.; González, J.B.; Martin, P.L.; Giroux, E.; Bilodeau, G.J.; Peter, K.A.; Doyle, V.P.; Aćimović, S.G. Identification and characterization of Colletotrichum species causing apple bitter rot in New York and description of C. noveboracense sp. nov. Sci. Rep. 2020, 10, 11043. [Google Scholar] [CrossRef]
- Vitale, A.; Alfenas, A.C.; Siqueira, D.L.D.; Magistà, D.; Perrone, G.; Polizzi, G. Cultivar resistance against Colletotrichum asianum in the world collection of mango germplasm in southeastern Brazil. Plants 2020, 9, 182. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Yang, Y.; Hyde, K.D.; Bahkali, A.H.; Liu, Z. Colletotrichum Species on Citrus Leaves in Guizhou and Yunnan Provinces, China. Cryptogam. Mycol. 2012, 33, 267–283. [Google Scholar]
- Cheng, B.P.; Huang, Y.H.; Song, X.B.; Peng, A.T.; Ling, J.F.; Chen, X. First Report of Colletotrichum siamense Causing Leaf Drop and Fruit Spot of Citrus reticulata Blanco cv. Shiyue Ju in China. Plant Dis. 2013, 97, 1508. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Chen, G.Q.; Hou, X.; Fu, Y.S.; Cai, L.; Hyde, K.D.; Li, H.Y. Colletotrichum Species Associated with Cultivated Citrus in China. Fungal Divers. 2013, 61, 61–74. [Google Scholar] [CrossRef]
- Douanla-Meli, C.; Unger, J. Phylogenetic Study of The Colletotrichum Species on Imported Citrus Fruits Uncovers a Low Diversity and A New Species in The Colletotrichum gigasporum Complex. Fungal Biol. 2017, 121, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Kunta, M.; Park, J.W.; Vedasharan, P.; da Graça, J.V.; Terry, M.D. First Report of Colletotrichum queenslandicum on Persian Lime Causing Leaf Anthracnose in The United States. Plant Dis. 2018, 102, 677. [Google Scholar] [CrossRef]
- Hu, W.L.; Ma, Y.Z.; Chen, J.Z. First Report of Citrus sinensis Anthracnose Caused by Colletotrichum fructicola in China. Plant Dis. 2019, 103, 1018. [Google Scholar] [CrossRef]
- Silva, A.O.; Savi, D.C.; Gomes, F.B.; Gos, F.M.V.R.; Silva, G.J.; Glienke, C. Identification of Colletotrichum species associated with postbloom fruit drop in Brazil through GAPDH sequencing analysis and multiplex PCR. Eur. J. Plant Pathol. 2016, 147, 731–748. [Google Scholar] [CrossRef]
- McGovern, R.J.; Seijo, T.E.; Hendricks, K.; Roberts, P.D. New report of Colletotrichum gloeosporioides causing postbloom fruit drop on citrus in Bermuda. Can. J. Plant Pathol. 2012, 34, 187–194. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [Green Version]
- Perrone, G.; Magistà, D.; Ismail, A.M. First report of Colletotrichum kahawae subsp. ciggaro on mandarin in Italy. J. Plant Pathol. 2016, 98, 682. [Google Scholar]
- Uysal, A.; Kurt, S. First Report of Colletotrichum karstii Causing Anthracnose on Citrus in the Mediterranean Region of Turkey. J. Plant Pathol. 2019, 101, 753. [Google Scholar] [CrossRef] [Green Version]
- Daoud, H.B.H.; Baraldi, E.; Iotti, M.; Leonardi, P.; Boughalleb-M’hamdi, N. Characterization and Pathogenicity of Colletotrichum spp. Causing Citrus Anthracnose in Tunisia. Phytopathol. Mediterr. 2019, 58, 175–185. [Google Scholar]
- Ramos, A.P.; Talhinhas, P.; Sreenivasaprasad, S.; Oliveira, H. Characterization of Colletotrichum gloeosporioides, As the Main Causal Agent of Citrus Anthracnose, and C. karstii As Species Preferentially Associated with Lemon Twig Dieback in Portugal. Phytoparasitica 2016, 44, 549–561. [Google Scholar] [CrossRef]
- Benyahia, H.; Jrifia, A.; Smaili, C.; Afellah, M.; Lamsetef, Y.; Timmer, L.W. First Report of Colletotrichum gloeosporioides Causing Wither-tip on Twigs and Tear Stain on Fruit of Citrus in Morocco. Plant Pathol. 2003, 52, 798. [Google Scholar] [CrossRef]
- Pérez-Mora, J.L.; Mora-Romero, G.A.; Beltrán-Peña, H.; García-León, E.; Lima, N.B.; Camacho-Tapia, M.; Tovar-Pedraza, J.M. First Report of Colletotrichum siamense and C. gloeosporioides Causing Anthracnose of Citrus Spp. In Mexico. Plant Dis. 2020, 105, 496. [Google Scholar] [CrossRef]
- Cruz-Lagunas, B.; Ortega-Acosta, S.A.; Reyes-García, G.; Toribio-Jiménez, J.; Juárez-López, P.; Guillén-Sánchez, D.; Damián-Nava, A.; Romero-Ramírez, Y.; Palemón-Alberto, F. Colletotrichum gloeosporioides Causes Anthracnose on Grapefruit (Citrus paradisi) In Mexico. Australas. Plant Dis. Notes 2020, 15, 31. [Google Scholar] [CrossRef]
- Fayyaz, A.; Sahi, S.T.; Nawaz-ul-Rehman, M.S.; Amrao, L. Molecular Characterization of Fungal Pathogens Associated with Citrus Wither-tip/Dieback from Major Citrus Growing Areas of Punjab, Pakistan. Int. J. Agric. Biol. 2020, 23, 1165–1170. [Google Scholar]
- Mahiout, D.; Bendahmane, B.S.; Benkada, M.Y.; Mekouar, H.; Berrahal, N.; Rickauer, M. First Report of Colletotrichum gloeosporioides on Citrus in Algeria. Phytopathol. Mediterr. 2018, 57, 355–359. [Google Scholar]
- Timmer, L.W.; Peever, T.L.; Solel, Z.; Akimitsu, K. Alternaria diseases of citrus-novel pathosystems. Phytopathol. Mediterr. 2003, 42, 99–112. [Google Scholar]
- Peever, T.L.; Su, G.; Carpenter-Boggs, L.; Timmer, L.W. Molecular systematics of citrus-associated Alternaria species. Mycologia 2004, 96, 119–134. [Google Scholar] [CrossRef]
- Dewdney, M.M.; Timmer, L.W. Florida Citrus Pest Management Guide: Alternaria Brown Spot; Institute of Food and Agricultural Sciences University of Florida: Gainesville, FL, USA, 2010. [Google Scholar]
- Gama, A.B.; Baggio, J.S.; Rebello, C.S.; Lourenco, S.D.A.; Gasparoto, M.C.D.G.; da Silva Junior, G.J.; Peres, N.A.; Amorim, L. Sensitivity of Colletotrichum acutatum Isolates from Citrus to Carbendazim, Difenoconazole, Tebuconazole, and Trifloxystrobin. Plant Dis. 2020, 104, 1621–1628. [Google Scholar] [CrossRef]
- Chitolina, G.M.; Silva-Junior, G.J.; Feichtenberger, E.; Pereira, R.G.; Amorim, L. Distribution of Alternaria alternata isolates with resistance to quinone outside inhibitor (QoI) fungicides in Brazilian orchards of tangerines and their hybrids. Crop Prot. 2021, 141, 105493. [Google Scholar] [CrossRef]
- Piccirillo, G.; Carrieri, R.; Polizzi, G.; Azzaro, A.; Lahoz, E.; Fernández-Ortuño, D.; Vitale, A. In vitro and in vivo activity of QoI fungicides against Colletotrichum gloeosporioides causing fruit anthracnose in Citrus sinensis. Sci. Hortic. 2018, 236, 90–95. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Woundernberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; DeVries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Chung, W.H.; Chung, W.C.; Peng, M.T.; Yang, H.R.; Huang, J.W. Specific detection of benzimidazole resistance in Colletotrichum gloeosporioides from fruit crops by PCR-RFLP. New Biotechnol. 2010, 27, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Templeton, M.D.; Rikkerink, E.H.A.; Solon, S.L.; Crowhurst, R.N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria themes and variations (236-243). Classification of citrus pathogens. Mycotaxon 1999, 70, 263–323. [Google Scholar]
- Peres, N.A.; Mackenzie, S.J.; Peever, T.L.; Timmer, L.W. Postbloom fruit drop of citrus and key lime anthracnose are caused by distinct phylogenetic lineages of Colletotrichum acutatum. Phytopathology 2008, 98, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Peever, T.L.; Ibanez, A.; Akimitsu, K.; Timmer, L.W. Worldwide phylogeography of the citrus brown spot pathogen, Alternaria alternata. Phytopathology 2002, 9, 794–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akimitsu, K.; Peever, T.L.; Timmer, L.W. Molecular, ecological and evolutionary approaches to understanding Alternaria diseases of citrus. Mol. Plant Pathol. 2003, 4, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.F.; Du, S.F.; Cai, M.L.; Luo, C.X. Identification of Anthracnose of Wild Citrus. Southwest China J. Agric. Sci. 2017, 30, 590–594. [Google Scholar]
- Vicent, A.; Armengol, J.; Sales, R.; García-Jiménez, J.; Alfaro-Lassala, F. First Report of Alternaria Brown Spot of Citrus in Spain. Plant Dis. 2000, 84, 1044. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Rewal, H.S.; Sethi, A. Pre-harvest stem-end rot in citrus cultivars due to Colletotrichum gloeosporioides. Eur. J. Hort Sci. 2007, 72, 20–25. [Google Scholar]
- Brown, G.E.; Eckert, J.W. Alternaria rot. In Compendium of Citrus Diseases; Timmer, L.W., Garnsey, S.M., Graham, J.H., Eds.; APS Press, Inc.: St. Paul, MN, USA, 2000; p. 37. [Google Scholar]
- Peever, T.L.; Carpenter-Boggs, L.W.; Timmer, L.W.; Carris, L.M.; Bhatia, A. Citrus Black rot is caused by phylogenetically distinct lineages of Alternaria alternata. Phytopathology 2005, 95, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Simmons, E.G. Alternaria: An Identification Manual; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007. [Google Scholar]
- Sutton, B.C. The genus Glomerella and its anamorph Colletotrichum. In Colletotrichum Biology, Pathology and Control; Bailey, J.A., Jeger, M.J., Eds.; CAB International: Wallingford, UK, 1992. [Google Scholar]
- Cannon, P.F.; Buddie, A.G.; Bridge, P.D. The typification of Colletotrichum gloeosporioides. Mycotaxon 2008, 104, 189–204. [Google Scholar]
- Frare, G.F.; Couto, H.T.Z.; Ciampi-Guillardi, M.; Amorim, L. The causal agent of citrus postbloom fruit drop, Colletotrichum acutatum, can survive on weeds. Australas. Plant Pathol. 2016, 45, 339–346. [Google Scholar] [CrossRef]
- Runkle, J.; Flocks, J.; Economos, J.; Dunlop, A.L. A systematic review of Mancozeb as a reproductive and developmental hazard. Environ. Int. 2017, 99, 29–42. [Google Scholar] [CrossRef]
- Wang, Z.; Kottawatta, K.S.A.; Kodithuwakku, S.P.; Fernando, T.S.; Lee, Y.L.; Ng, E.H.Y.; Yeung, W.S.B.; Lee, K.F. The fungicide Mancozeb reduces spheroid attachment onto endometrial epithelial cells through downregulation of estrogen receptor β and integrin β3 in Ishikawa cells. Ecotoxicol. Environ. Saf. 2021, 208, 111606. [Google Scholar] [CrossRef] [PubMed]
- Triantafyllidis, V.; Zotos, A.; Kosma, C.; Kokkotos, E. Environmental implications from long-term Citrus cultivation and wide use of Cu fungicides in Mediterranean soils. Water Air Soil Pollut. 2020, 231, 218. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Köhl, J.; Jones, J.B.; Aubertot, J.N. Thirteen decades of antimicrobial copper compounds applied in agriculture. A review. Agron. Sustain. Dev. 2018, 38, 28. [Google Scholar] [CrossRef] [Green Version]
- Cuenca, J.; Aleza, P.; Garcia-lor, A.; Ollitrault, P.; Navarro, L. Fine mapping for identification of Citrus Alternaria brown spot candidate resistance genes and development of new SNP markers for marker-assisted selection. Front. Plant. Sci. 2016, 7, 1948. [Google Scholar] [CrossRef] [Green Version]
- Arlotta, C.; Ciacciulli, A.; Strano, M.C.; Cafaro, V.; Salonia, F.; Caruso, P.; Licciardello, C.; Russo, G.; Malcolm Smith, M.W.; Cuenca, J.; et al. Disease resistant Citrus breeding using newly developed high resolution melting and CAPS protocols for Alternaria brown spot marker assisted selection. Agronomy 2020, 10, 1368. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
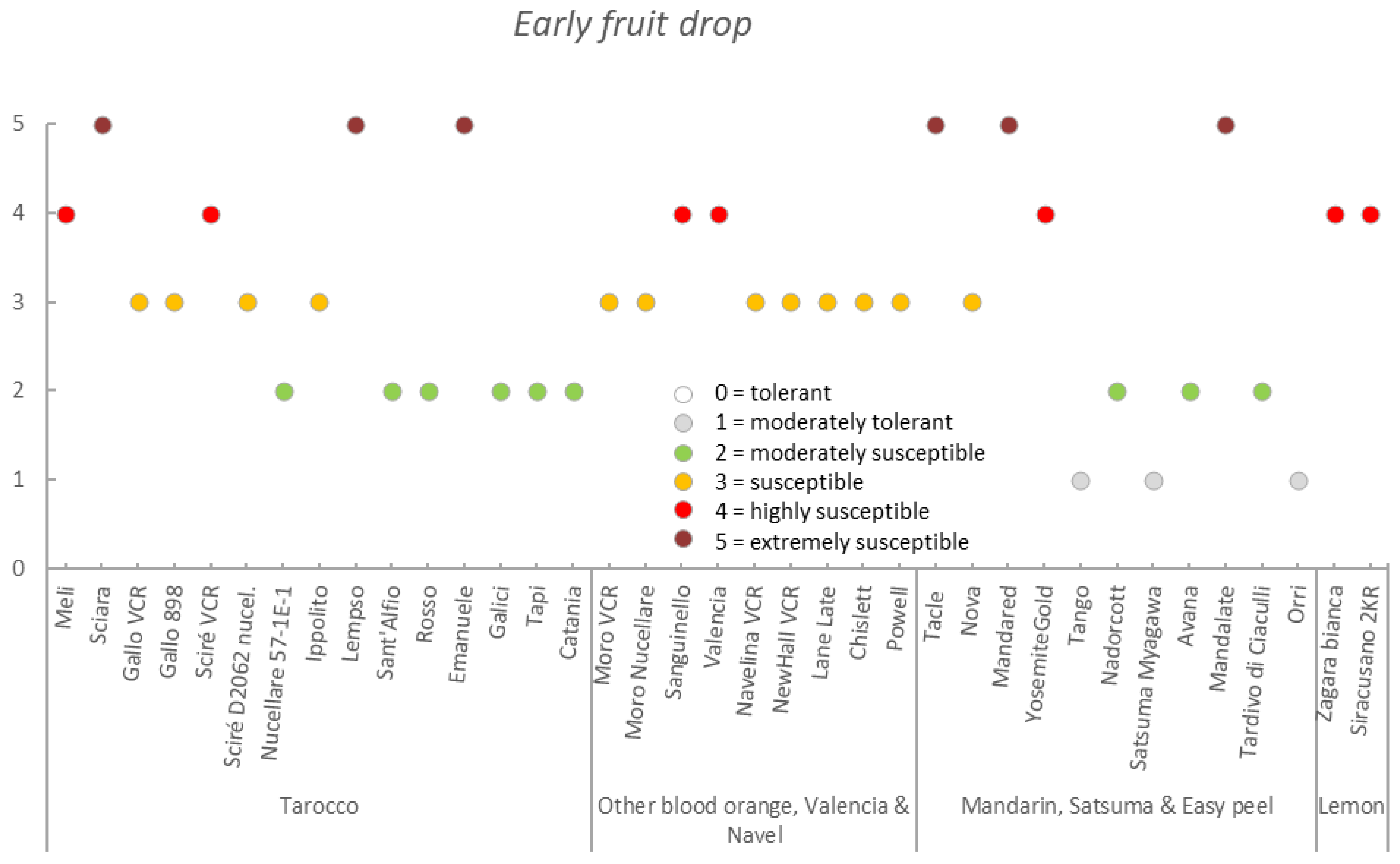
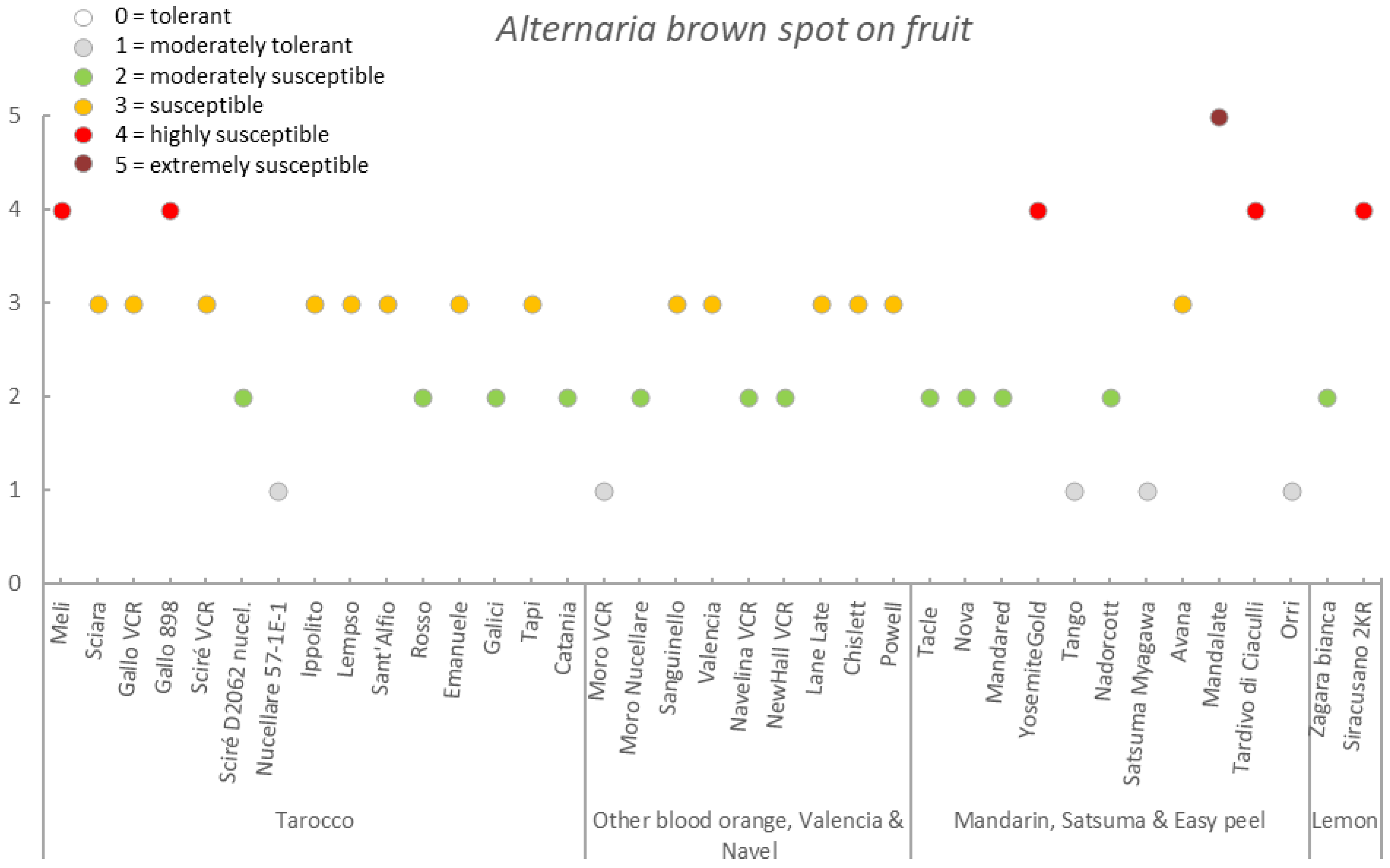
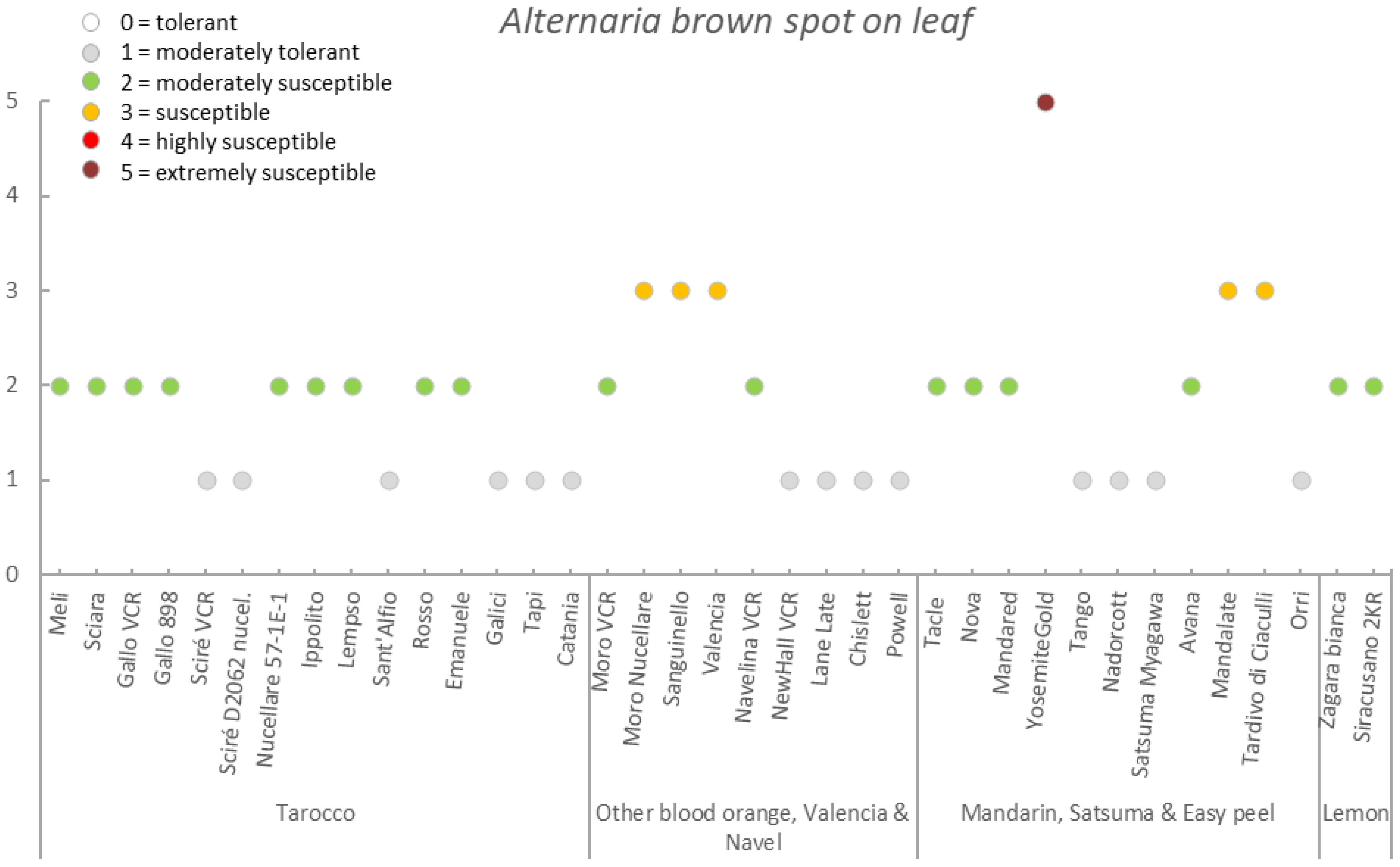
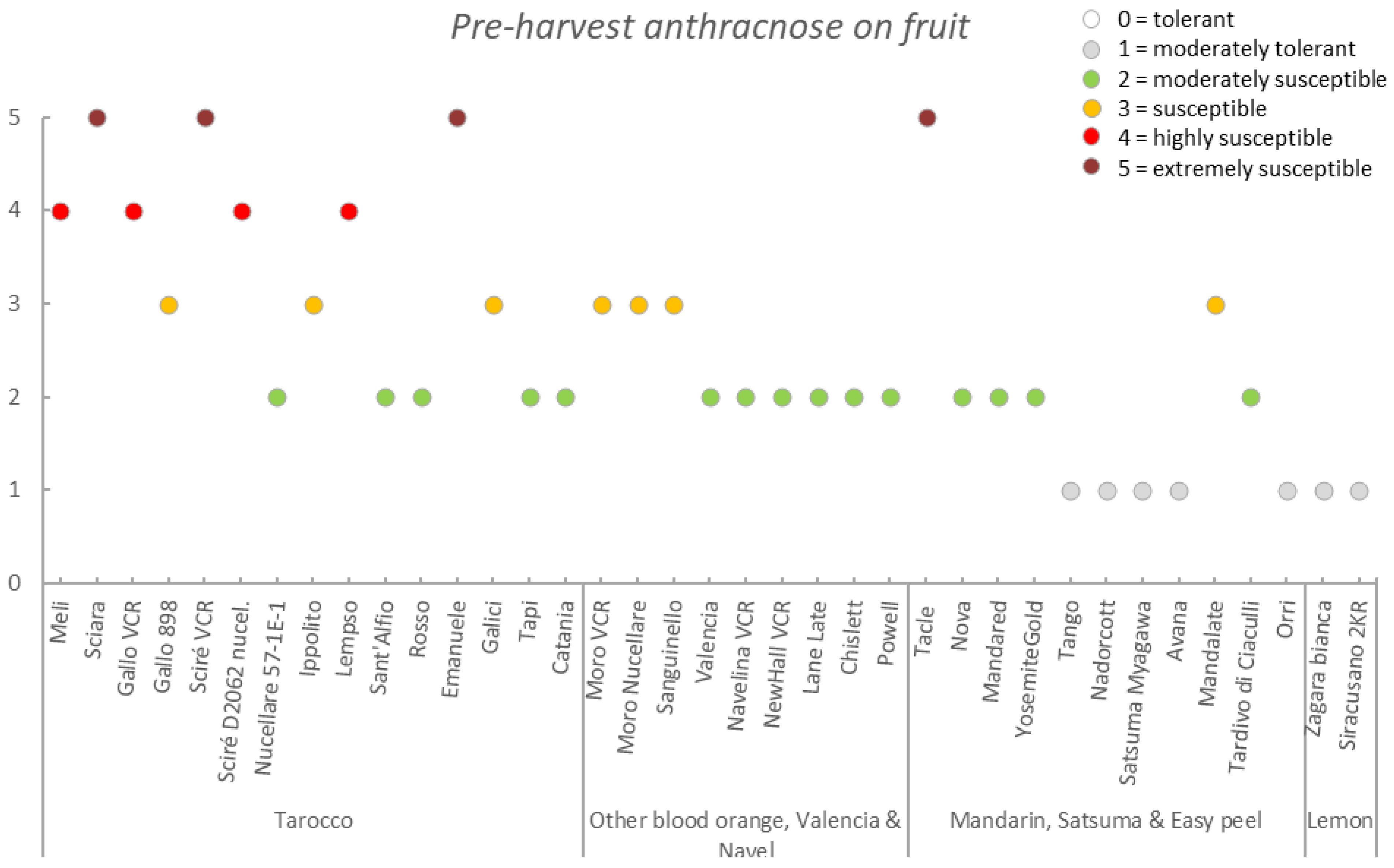
| Germplasm | Symptoms | |||||||
|---|---|---|---|---|---|---|---|---|
| Group | Accession | Early Fruit Drop x,y,z | Alternaria Brown Spot on Fruit x,y,z | Alternaria Brown Spot on Leaf x,y,z | Anthracnose on Leaf x,y,z | Pre-Harvest x,y,z Anthracnose on Fruit | Pre-Harvest Fruit Drop x,y,z | Twig Dieback and Defoliation x,y,z |
| Tarocco | Meli | 3.4 ± 0.6 (2–4) | 2.7 ± 0.5 (2–4) | 1.3 ± 0.4 (1–2) | 2.1 ± 0.5 (1–3) | 2.6 ± 0.7 (2–4) | 2.7 ± 0.4 (2–3) | 2.4 ± 0.6 (1–3) |
| Sciara | 3.2 ± 0.7 (2–5) | 2.7 ± 0.5 (2–3) | 1.4 ± 0.5 (1–2) | 3.8 ± 0.8 (2–5) | 4.0 ± 0.6 (2–5) | 4.0 ± 0.8 (2–5) | 3.5 ± 0.8 (2–5) | |
| Gallo VCR | 2.2 ± 0.6 (1–3) | 2.0 ± 0.8 (1–3) | 1.1 ± 0.2 (1–2) | 2.6 ± 0.7 (1–4) | 2.5 ± 0.8 (1–4) | 2.4 ± 0.6 (1–3) | 2.8 ± 0.6 (1–4) | |
| Gallo 898 | 2.7 ± 0.5 (2–3) | 2.7 ± 0.6 (2–4) | 1.2 ± 0.4 (1–2) | 2.6 ± 0.5 (2–3) | 2.5 ± 0.5 (2–3) | 2.2 ± 0.7 (1–3) | 2.8 ± 0.4 (2–3) | |
| Sciré VCR | 3.1 ± 0.6 (2–4) | 1.4 ± 0.5 (1–3) | 1.0 ± 0.0 (1–1) | 3.7 ± 0.7 (3–5) | 4.0 ± 0.7 (3–5) | 3.6 ± 0.6 (2–5) | 3.9 ± 0.6 (3–5) | |
| Sciré D2062 nucel. | 3.0 ± 0.0 (3–3) | 1.9 ± 0.3 (1–2) | 1.0 ± 0.0 (1–1) | 3.1 ± 0.8 (2–4) | 3.0 ± 0.7 (2–4) | 2.8 ± 0.4 (2–3) | 3.0 ± 0.0 (3–3) | |
| Nucellare 57-1E-1 | 1.8 ± 0.4 (1–2) | 1.0 ± 0.0 (1–1) | 1.0 ± 0.2 (1–2) | 1.6 ± 0.6 (1–3) | 1.2 ± 0.4 (1–2) | 1.6 ± 0.5 (1–2) | 1.7 ± 0.5 (1–3) | |
| Ippolito | 2.2 ± 0.4 (2–3) | 2.1 ± 0.4 (2–3) | 1.0 ± 0.2 (1–2) | 2.6 ± 0.5 (2–3) | 2.1 ± 0.4 (1–3) | 2.1 ± 0.3 (2–3) | 2.1 ± 0.4 (2–3) | |
| Lempso | 3.4 ± 0.7 (2–5) | 1.6 ± 0.6 (1–3) | 1.1 ± 0.2 (1–2) | 3.4 ± 0.7 (2–4) | 3.0 ± 0.8 (2–4) | 2.8 ± 0.6 (2–4) | 3.6 ± 0.7 (3–5) | |
| Sant’Alfio | 1.8 ± 0.4 (1–2) | 2.3 ± 0.7 (1–3) | 1.0 ± 0.0 (1–1) | 2.0 ± 0.0 (2–2) | 1.9 ± 0.3 (1–2) | 2.0 ± 0.0 (2–2) | 1.9 ± 0.3 (1–2) | |
| Rosso | 2.0 ± 0.0 (2–2) | 1.8 ± 0.4 (1–2) | 1.2 ± 0.4 (1–2) | 2.0 ± 0.0 (2–2) | 1.8 ± 0.4 (1–2) | 2.2 ± 0.4 (2–3) | 2.0 ± 0.7 (1–3) | |
| Emanuele | 4.2 ± 0.4 (4–5) | 2.6 ± 0.5 (2–3) | 2.0 ± 0.0 (2–2) | 4.0 ± 0.0 (4–4) | 5.0 ± 0.0 (5–5) | 5.0 ± 0.0 (5–5) | 4.0 ± 0.7 (3–5) | |
| Galici | 1.9 ± 0.3 (1–2) | 1.2 ± 0.4 (1–2) | 1.0 ± 0.0 (1–1) | 2.2 ± 0.4 (2–3) | 2.0 ± 0.5 (1–3) | 2.2 ± 0.4 (2–3) | 2.2 ± 0.4 (2–3) | |
| Tapi | 1.5 ± 0.5 (1–2) | 1.6 ± 0.9 (1–3) | 1.0 ± 0.0 (1–1) | 1.7 ± 0.5 (1–2) | 1.5 ± 0.5 (1–2) | 1.7 ± 0.5 (1–2) | 1.8 ± 0.4 (1–2) | |
| Catania | 2.0 ± 0.0 (2–2) | 1.3 ± 0.6 (1–2) | 1.0 ± 0.0 (1–1) | 2.0 ± 0.0 (2–2) | 1.8 ± 0.4 (1–2) | 2.0 ± 0.0 (2–2) | 2.0 ± 0.0 (2–2) | |
| Mean | 2.6 ± 0.8 | 1.9 ± 0.6 | 1.2 ± 0.3 | 2.6 ± 0.8 | 2.6 ± 1.1 | 2.6 ± 0.9 | 2.7 ± 0.8 | |
| Germplasm | Symptoms | |||||||
|---|---|---|---|---|---|---|---|---|
| Group | Accession | Early Fruit Drop x,y,z | Alternaria Brown Spot on Fruit x,y,z | Alternaria Brown Spot on Leaf x,y,z | Anthracnose on Leaf x,y,z | Pre-Harvest x,y,z Anthracnose on Fruit | Pre-Harvest Fruit Drop x,y,z | Twig Dieback and Defoliation x,y,z |
| Other blood orange, Valencia & Navel | Moro VCR | 2.3 ± 0.7 (1–3) z | 1.0 ± 0.0 (1–1) z | 1.7 ± 0.5 (1–2) z | 1.9 ± 0.3 (1–2) z | 1.9 ± 0.6 (1–3) z | 1.7 ± 0.5 (1–2) z | 2.3 ± 0.5 (2–3) z |
| Moro Nucellare | 2.3 ± 0.5 (2–3) | 1.2 ± 0.4 (1–2) | 1.5 ± 0.7 (1–3) | 2.0 ± 0.3 (1–3) | 1.9 ± 0.4 (1–3) | 1.9 ± 0.5 (1–3) | 2.3 ± 0.5 (2–3) | |
| Sanguinello | 2.9 ± 1.3 (1–4) | 2.0 ± 0.9 (1–3) | 1.9 ± 0.8 (1–3) | 1.7 ± 0.5 (1–2) | 1.8 ± 0.7 (1–3) | 2.2 ± 1.0 (1–4) | 2.7 ± 0.5 (2–3) | |
| Valencia | 3.2 ± 0.6 (2–4) | 2.4 ± 0.5 (2–3) | 2.0 ± 0.7 (1–3) | 2.3 ± 0.6 (1–3) | 1.8 ± 0.4 (1–2) | 2.4 ± 0.5 (2–3) | 2.7 ± 0.7 (2–4) | |
| Navelina VCR | 2.3 ± 0.5 (2–3) | 1.3 ± 0.5 (1–2) | 1.0 ± 0.2 (1–2) | 1.5 ± 0.5 (1–2) | 1.5 ± 0.5 (1–2) | 1.8 ± 0.4 (1–2) | 2.1 ± 0.5 (1–3) | |
| NewHall VCR | 2.2 ± 0.4 (2–3) | 1.3 ± 0.5 (1–2) | 1.0 ± 0.0 (–1) | 1.3 ± 0.5 (1–2) | 1.7 ± 0.5 (1–2) | 1.9 ± 0.3 (1–2) | 2.1 ± 0.5 (1–3) | |
| Lane Late | 2.4 ± 0.5 (2–3) | 2.6 ± 0.5 (2–3) | 1.0 ± 0.0 (1–1) | 1.5 ± 0.5 (1–2) | 1.5 ± 0.5 (1–2) | 1.8 ± 0.4 (1–2) | 1.9 ± 0.5 (1–3) | |
| Chislett | 2.3 ± 0.5 (2–3) | 2.5 ± 0.5 (2–3) | 1.0 ± 0.0 (1–1) | 2.0 ± 0.0 (2–2) | 2.0 ± 0.0 (2–2) | 2.0 ± 0.0 (2–2) | 2.2 ± 0.4 (2–3) | |
| Powell | 2.3 ± 0.5 (2–3) | 2.2 ± 0.4 (2–3) | 1.0 ± 0.0 (1–1) | 1.5 ± 0.5 (1–2) | 1.7 ± 0.5 (1–2) | 1.5 ± 0.5 (1–2) | 1.7 ± 0.5 (1–2) | |
| Mean | 2.5 ± 0.3 | 1.8 ± 0.6 | 1.3 ± 0.4 | 1.8 ± 0.3 | 1.8 ± 0.2 | 1.9 ± 0.3 | 2.2 ± 0.3 | |
| Germplasm | Symptoms | |||||||
|---|---|---|---|---|---|---|---|---|
| Group | Accession | Early Fruit Drop x,y,z | Alternaria Brown Spot on Fruit x,y,z | Alternaria Brown Spot on Leaf x,y,z | Anthracnose on Leaf x,y,z | Pre-Harvest x,y,z Anthracnose on Fruit | Pre-Harvest Fruit Drop x,y,z | Twig Dieback and Defoliation x,y,z |
| Mandarin, Satsuma & Easy peel | Tacle | 3.2 ± 1.0 (2–5) z | 1.5 ± 0.5 (1–2) z | 1.2 ± 0.4 (1–2) z | 2.3 ± 0.8 (1–4) z | 2.6 ± 1.2 (1–5) z | 4.0 ± 0.8 (2–5) z | 3.2 ± 0.7 (2–4) z |
| Nova | 2.1 ± 0.5 (1–3) | 1.6 ± 0.5 (1–2) | 1.1 ± 0.4 (1–2) | 1.6 ± 0.6 (1–3) | 1.4 ± 0.5 (1–2) | 1.8 ± 0.6 (1–3) | 2.2 ± 0.6 (1–3) | |
| Mandared | 3.5 ± 1.4 (2–5) | 1.6 ± 0.5 (1–2) | 1.5 ± 0.5 (1–2) | 4.2 ± 0.7 (3–5) | 1.9 ± 0.3 (1–2) | 4.4 ± 0.5 (4–5) | 4.6 ± 0.5 (4–5) | |
| Yosemite Gold | 2.8 ± 1.1 (2–4) | 3.6 ± 0.5 (3–4) | 2.4 ± 1.9 (1–5) | 1.2 ± 0.4 (1–2) | 1.8 ± 0.4 (1–2) | 2.4 ± 0.9 (1–3) | 2.0 ± 0.7 (1–3) | |
| Tango | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | |
| Nadorcott | 2.0 ± 0.0 (2–2) | 1.7 ± 0.6 (1–2) | 1.0 ± 0.0 (1–1) | 2.0 ± 0.0 (2–2) | 1.0 ± 0.0 (1–1) | 2.0 ± 0.0 (2–2) | 2.0 ± 0.0 (2–2) | |
| Satsuma Miagawa | 1.0 ± 0.0 (1–1) | 1.0 ± 0.0 (1–1) | 1.0 ± 0.0 (1–1) | 1.0 ± 0.0 (1–1) | 1.0 ± 0.0 (1–1) | 1.0 ± 0.0 (1–1) | 1.0 ± 0.0 (1–1) | |
| Avana | 1.6 ± 0.5 (1–2) | 2.4 ± 0.5 (2–3) | 2.0 ± 0.0 (2–2) | 1.0 ± 0.0 (1–1) | 1.0 ± 0.0 (1–1) | 1.6 ± 0.5 (1–2) | 1.2 ± 0.4 (1–2) | |
| Mandalate | 2.8 ± 1.2 (2–5) | 3.9 ± 0.9 (3–5) | 2.4 ± 0.5 (2–3) | 1.8 ± 0.4 (1–2) | 2.6 ± 0.5 (2–3) | 2.1 ± 0.3 (2–3) | 2.3 ± 0.5 (2–3) | |
| Tardivo di Ciaculli | 2.0 ± 0.0 (2–2) | 3.1 ± 0.6 (2–4) | 1.7 ± 0.7 (1–3) | 1.1 ± 0.3 (1–2) | 1.1 ± 0.3 (1–2) | 1.3 ± 0.5 (1–2) | 1.7 ± 0.5 (1–2) | |
| Orri | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | 1.0 ± 0.0 * | |
| Mean | 2.1 ± 0.9 | 2.0 ± 1.1 | 1.5 ± 0.6 | 1.7 ± 1.0 | 1.5 ± 0.6 | 2.0 ± 1.2 | 2.0 ± 1.1 | |
| Lemon | Femminello Zagara Bianca | 2.6 ± 0.8 (2–4) z | 1.8 ± 0.4 (1–2) z | 1.7 ± 0.5 (1–2) z | 1.0 ± 0.0 (1–1) z | 1.0 ± 0.0 (1–1) z | 0.0 ± 0.0 z | 1.7 ± 0.5 (1–2) z |
| Femminello Siracusano 2KR | 2.6 ± 0.8 (2–4) | 2.9 ± 0.7 (2–4) | 1.5 ± 0.5 (1–2) | 1.2 ± 0.4 (1–2) | 1.0 ± 0.0 (1–1) | 0.4 ± 0.8 (0–2) | 1.7 ± 0.5 (1–2) | |
| Mean | 2.6 ± 0.0 | 2.4 ± 0.8 | 1.6 ± 0.1 | 1.1 ± 0.1 | 1.0 ± 0.0 | 0.2 ± 0.3 | 1.7 ± 0.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitale, A.; Aiello, D.; Azzaro, A.; Guarnaccia, V.; Polizzi, G. An Eleven-Year Survey on Field Disease Susceptibility of Citrus Accessions to Colletotrichum and Alternaria Species. Agriculture 2021, 11, 536. https://doi.org/10.3390/agriculture11060536
Vitale A, Aiello D, Azzaro A, Guarnaccia V, Polizzi G. An Eleven-Year Survey on Field Disease Susceptibility of Citrus Accessions to Colletotrichum and Alternaria Species. Agriculture. 2021; 11(6):536. https://doi.org/10.3390/agriculture11060536
Chicago/Turabian StyleVitale, Alessandro, Dalia Aiello, Antonino Azzaro, Vladimiro Guarnaccia, and Giancarlo Polizzi. 2021. "An Eleven-Year Survey on Field Disease Susceptibility of Citrus Accessions to Colletotrichum and Alternaria Species" Agriculture 11, no. 6: 536. https://doi.org/10.3390/agriculture11060536