
Segregation Distortion for Male Parents in High Density Genetic Maps from Reciprocal Crosses between Two Self-Incompatible Cultivars Confirms a Gametophytic System for Self-Incompatibility in Citrus

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Plant Genotyping
2.3. Linkage Analysis and Genetic Mapping
2.4. Analysis of Segregation Distortion
2.5. Haplotype Analysis
3. Results
3.1. SNP Calling
3.2. Genetic Linkage Maps of ‘Fortune’ Mandarin and ‘Ellendale’ Tangor
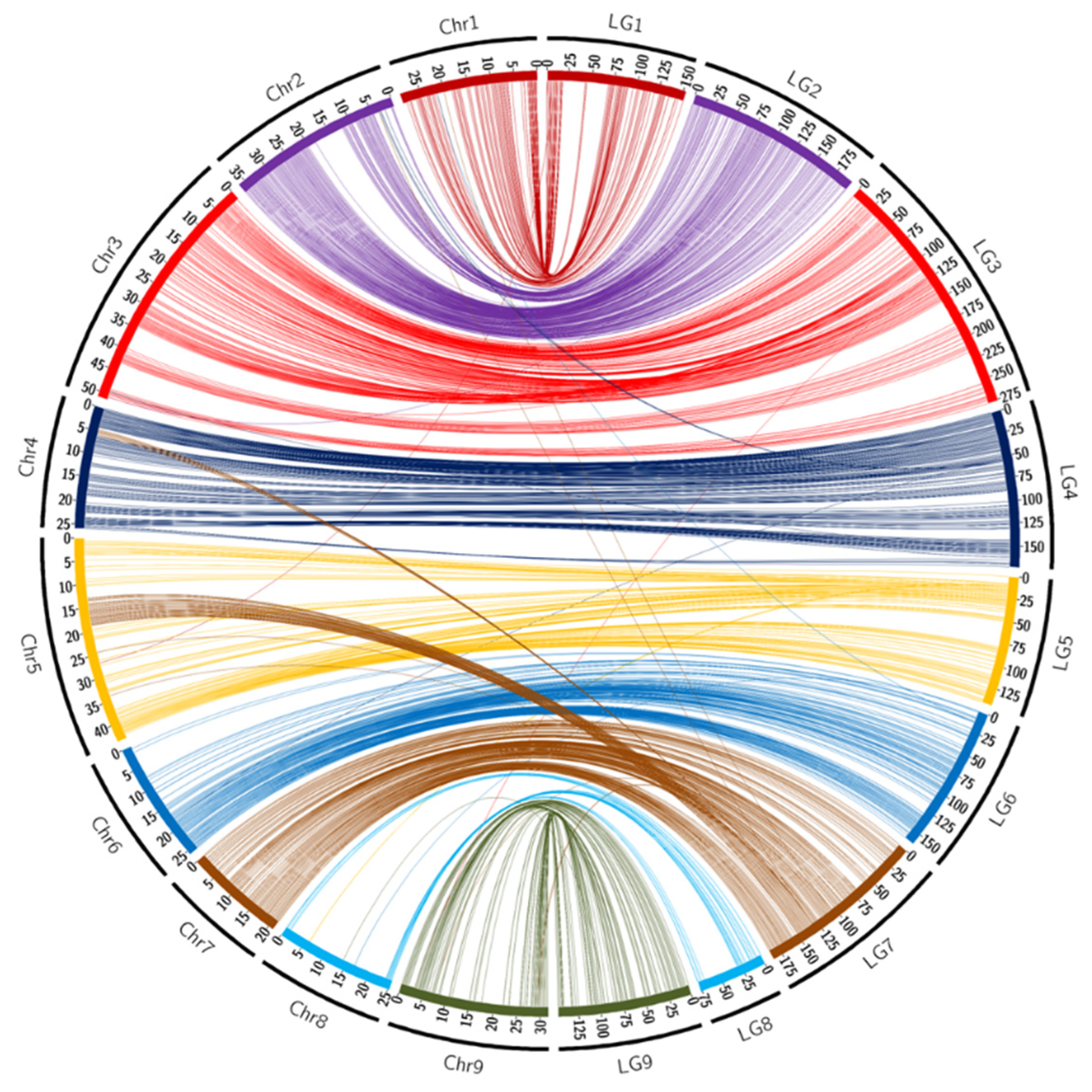
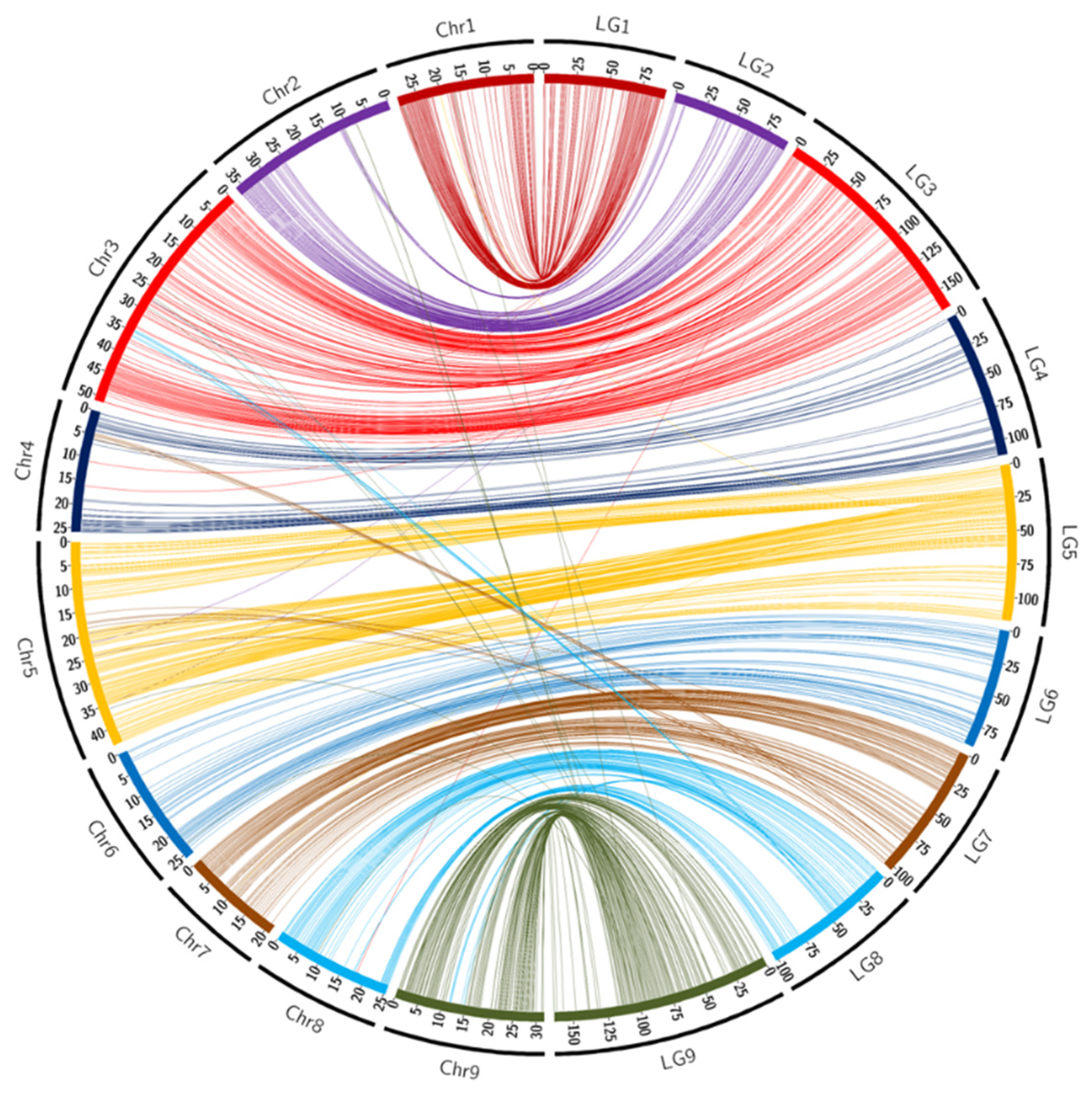
3.3. Synteny and Collinearity with the Reference Genome of Clementine
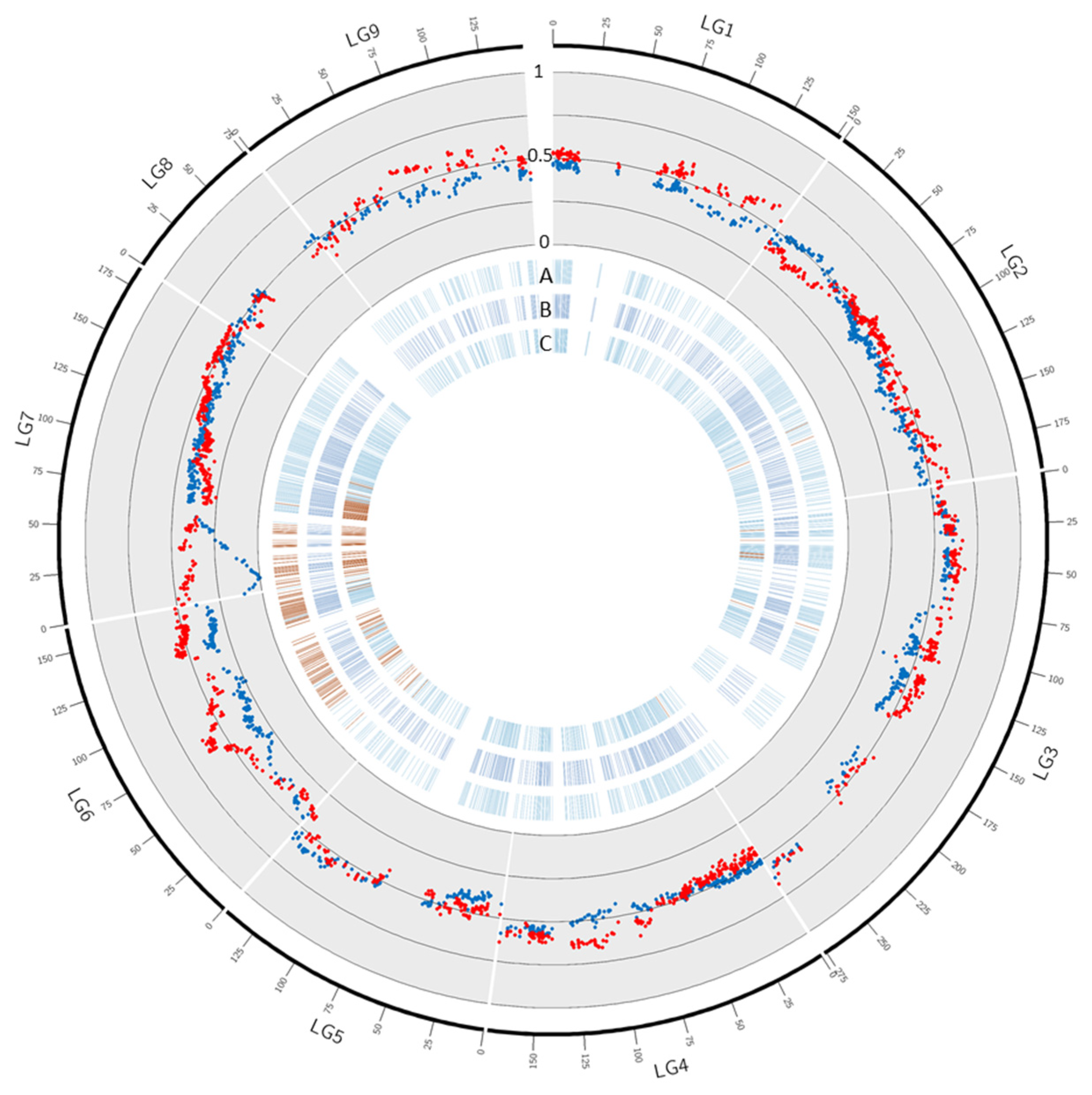
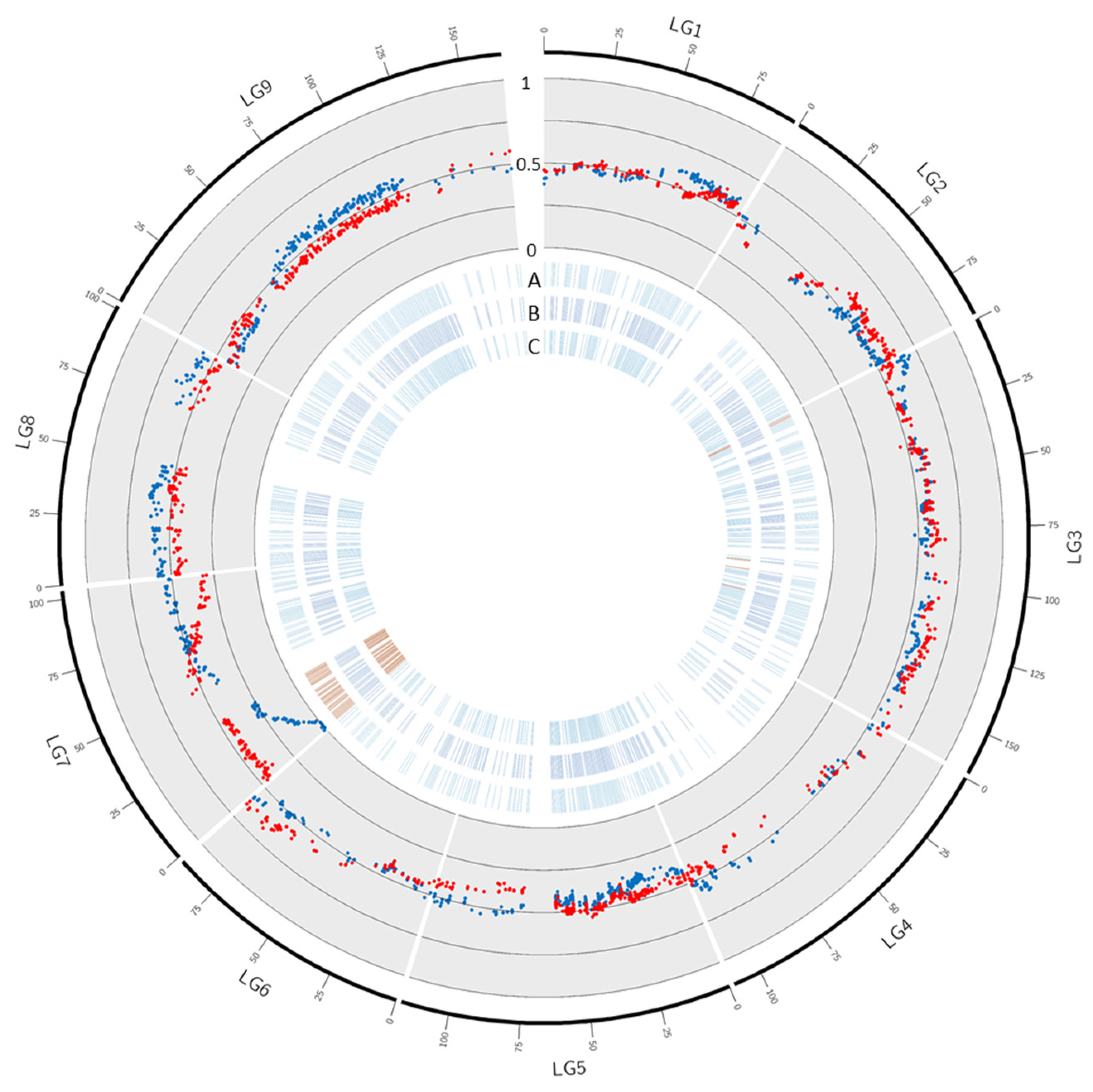
3.4. Segregation Distortion
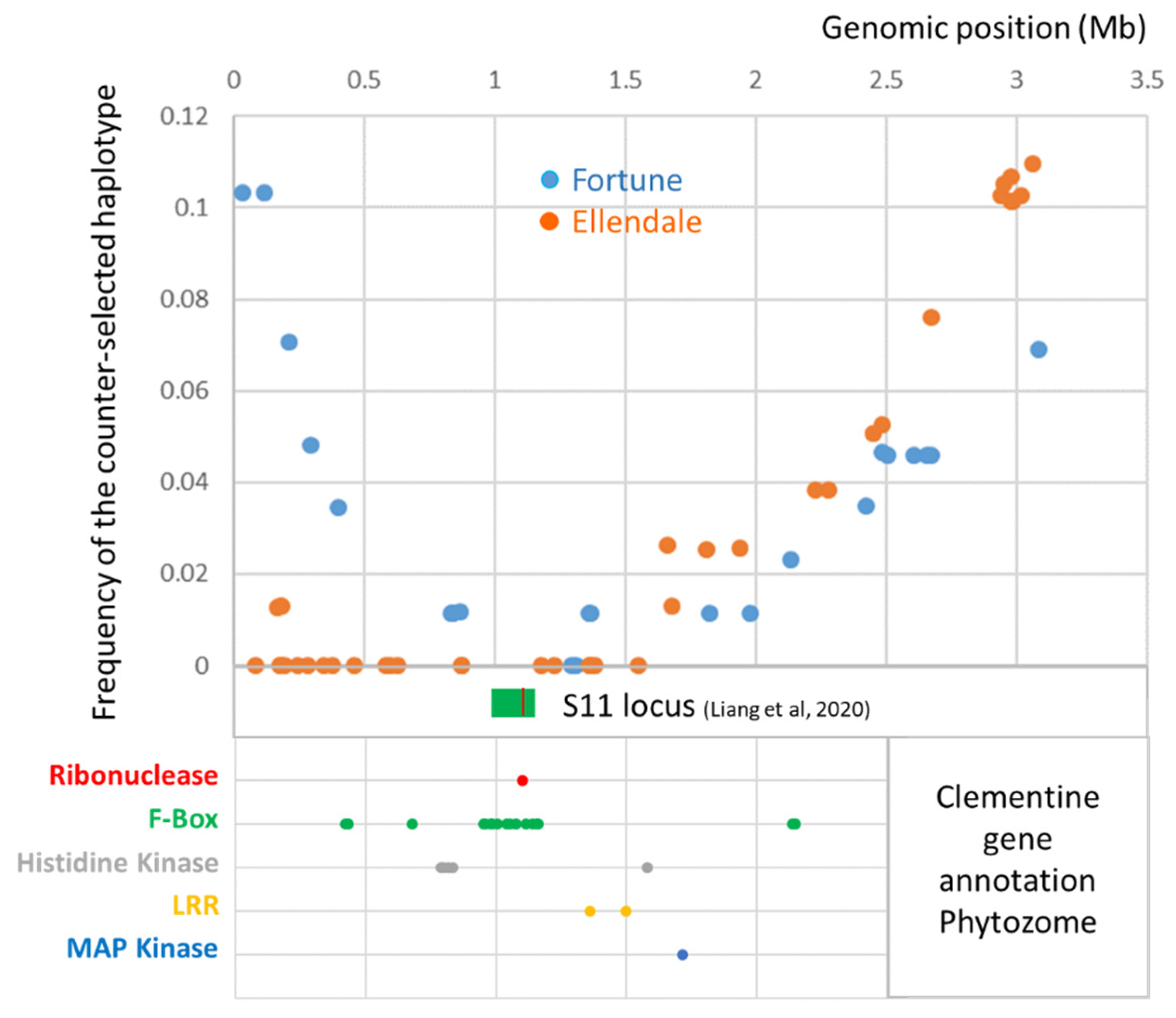
3.5. Gene Annotation in the Fully Skewed Region of Chromosome 7 in Male Parents
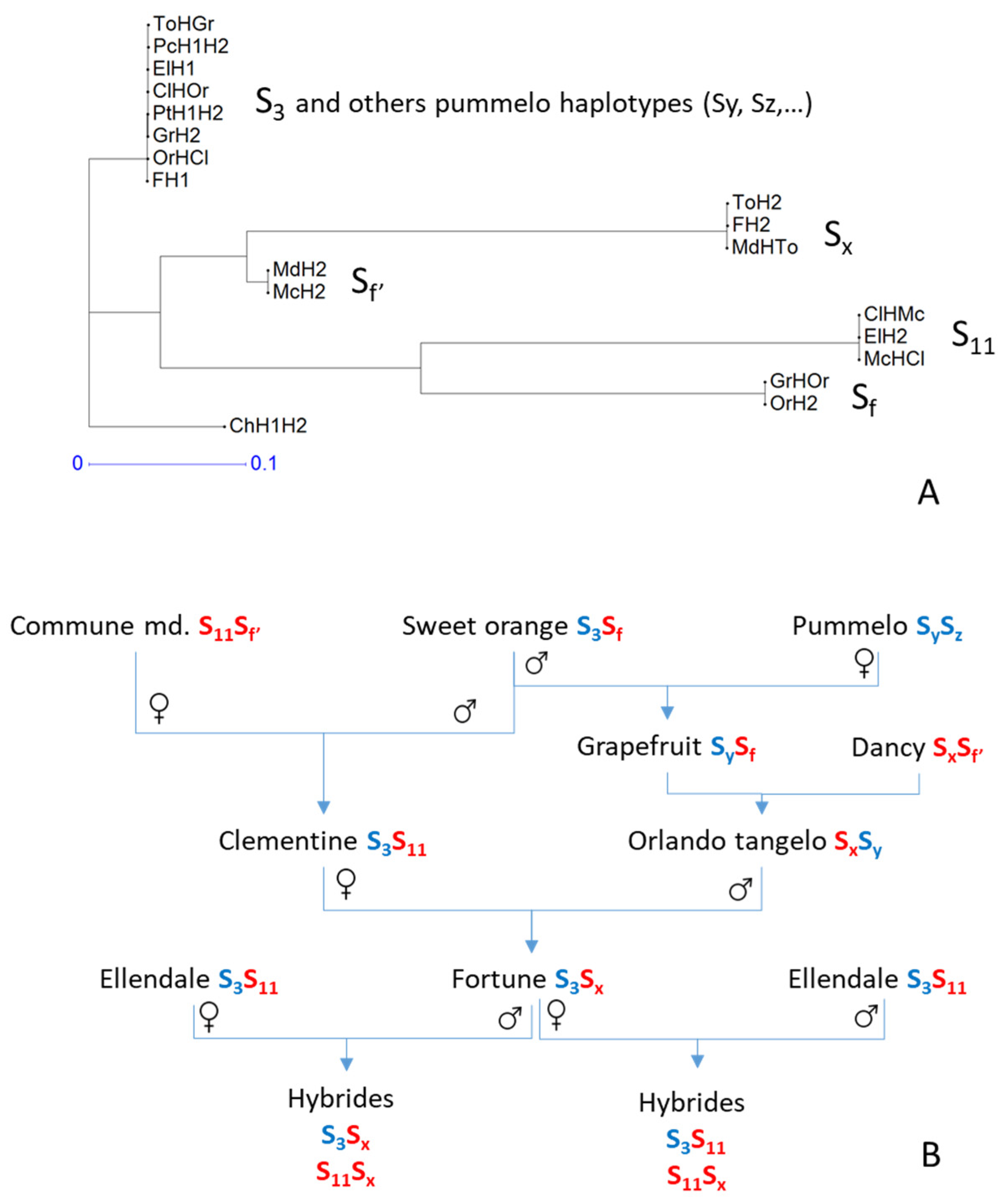
3.6. Haplotype Structure and Origin of ‘Fortune’ Mandarin and ‘Ellendale’ Tangor
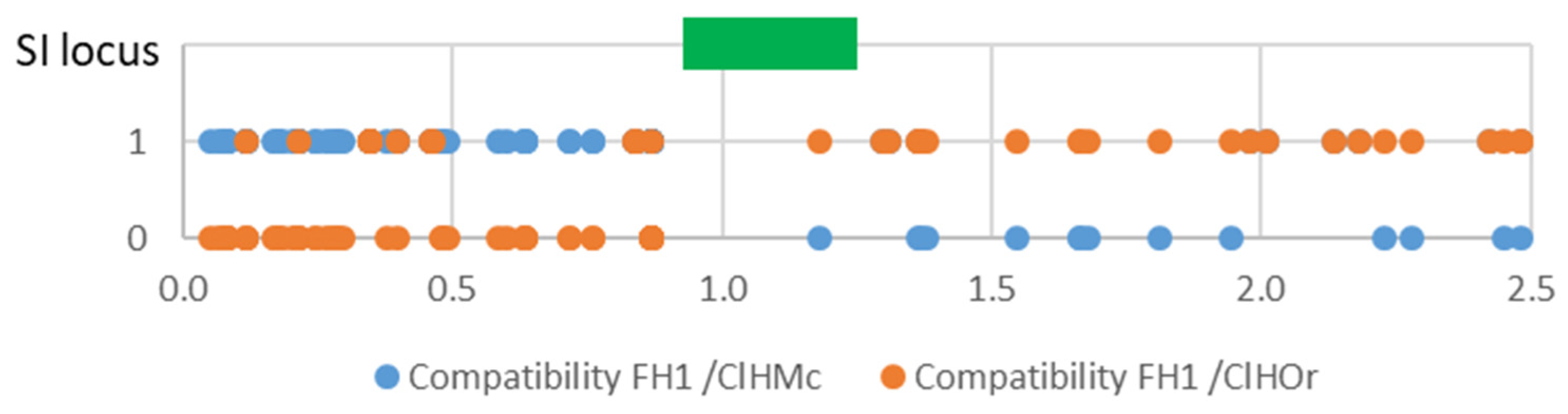
3.7. Haplotypic Structure around the SI Locus
4. Discussion
4.1. High Density Genetic Maps of ‘Fortune’ Mandarin and Ellendale ‘Tangor’ Were Produced
4.2. The Origins of ‘Fortune’ Mandarin and ‘Ellendale’ Tangor Were Assessed through Analysis of Chromosome Haplotypes
4.3. Segregation Distortion in the Male Parent Revealed a Genomic Region Involved in Self-Incompatibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goto, S.; Yoshioka, T.; Ohta, S.; Kita, M.; Hamada, H.; Shimizu, T. QTL Mapping of Male Sterility and Transmission Pattern in Progeny of Satsuma Mandarin. PLoS ONE 2018, 13, e0200844. [Google Scholar] [CrossRef] [PubMed]
- Aleza, P.; Juarez, J.; Cuenca, J.; Ollitrault, P.; Navarro, L. Recovery of Citrus Triploid Hybrids by Embryo Rescue and Flow Cytometry from 2x x 2x Sexual Hybridisation and Its Application to Extensive Breeding Programs. Plant Cell Rep. 2010, 29, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.; Smith, M.W.; Froelicher, Y.; Russo, G.; Gmitter, F.G. Chapter 7—Traditional breeding. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F.G., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 129–148. ISBN 978-0-12-812163-4. [Google Scholar]
- Soost, R.K. The Incompatibility Gene System in Citrus. Proc. First Int. Citrus Symp. 1968, 1, 189–190. [Google Scholar]
- Vardi, A.; Neumann, H.; Frydman-Shani, A.; Yaniv, Y.; Spiegel-Roy, P. Tentative Model on the Inheritance of Juvenility, Self-Incompatibility and Parthenocarpy. Acta Hortic. 2000, 199–206. [Google Scholar] [CrossRef]
- Raveh, E.; Goldenberg, L.; Porat, R.; Carmi, N.; Gentile, A.; La Malfa, S. Conventional Breeding of Cultivated Citrus Varieties. In The Citrus Genome; Gentile, A., La Malfa, S., Deng, Z., Eds.; Compendium of Plant Genomes; Springer International Publishing: Cham, Switzerland, 2020; pp. 33–48. ISBN 978-3-030-15308-3. [Google Scholar]
- Distefano, G.; Casas, G.L.; Malfa, S.L.; Gentile, A.; Tribulato, E.; Herrero, M. Pollen Tube Behavior in Different Mandarin Hybrids. J. Am. Soc. Hortic. Sci. 2009, 134, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Gambetta, G.; Gravina, A.; Fasiolo, C.; Fornero, C.; Galiger, S.; Inzaurralde, C.; Rey, F. Self-Incompatibility, Parthenocarpy and Reduction of Seed Presence in ‘Afourer’ Mandarin. Sci. Hortic. 2013, 164, 183–188. [Google Scholar] [CrossRef]
- Ferrer, M.M.; Good, S.V. Self-Sterility in Flowering Plants: Preventing Self-Fertilization Increases Family Diversification Rates. Ann. Bot. 2012, 110, 535–553. [Google Scholar] [CrossRef] [Green Version]
- McClure, B. S-RNase and SLF Determine S-Haplotype–Specific Pollen Recognition and Rejection. Plant Cell 2004, 16, 2840–2847. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Nishio, T. Commonalities and Differences between Brassica and Arabidopsis Self-Incompatibility. Hortic. Res. 2014, 1, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, N.; Singh, S. Progress on Deciphering the Molecular Aspects of Cell-to-Cell Communication in Brassica Self-Incompatibility Response. 3 Biotech 2018, 8, 347. [Google Scholar] [CrossRef]
- Maliepaard, C.; Alston, F.H.; van Arkel, G.; Brown, L.M.; Chevreau, E.; Dunemann, F.; Evans, K.M.; Gardiner, S.; Guilford, P.; van Heusden, A.W.; et al. Aligning Male and Female Linkage Maps of Apple (Malus Pumila mill.) Using Multi-Allelic Markers. Theor. Appl. Genet. 1998, 97, 60–73. [Google Scholar] [CrossRef]
- Claessen, H.; Keulemans, W.; Van de Poel, B.; De Storme, N. Finding a Compatible Partner: Self-Incompatibility in European Pear (Pyrus Communis); Molecular Control, Genetic Determination, and Impact on Fertilization and Fruit Set. Front. Plant Sci. 2019, 10, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, D.; Baraket, G.; Perez, V.; Ben Mustapha, S.; Salhi-Hannachi, A.; Hormaza, J.I. Analysis of Self-Incompatibility and Genetic Diversity in Diploid and Hexaploid Plum Genotypes. Front. Plant Sci. 2019, 10, 896. [Google Scholar] [CrossRef] [Green Version]
- Brancher, T.L.; Hawerroth, M.C.; Kvitschal, M.V.; Manenti, D.C.; Guidolin, A.F.; Brancher, T.L.; Hawerroth, M.C.; Kvitschal, M.V.; Manenti, D.C.; Guidolin, A.F. Self-Incompatibility Alleles in Important Genotypes for Apple Breeding in Brazil. Crop Breed. Appl. Biotechnol. 2020, 20. [Google Scholar] [CrossRef]
- Knight, R.; Rogers, H.H. Sterility in Theobroma cacao L. Nature 1953, 172, 164. [Google Scholar] [CrossRef] [PubMed]
- Alagna, F.; Caceres, M.E.; Pandolfi, S.; Collani, S.; Mousavi, S.; Mariotti, R.; Cultrera, N.G.M.; Baldoni, L.; Barcaccia, G. The Paradox of Self-Fertile Varieties in the Context of Self-Incompatible Genotypes in Olive. Front. Plant Sci. 2019, 10, 725. [Google Scholar] [CrossRef]
- Zhang, S.; Liang, M.; Wang, N.; Xu, Q.; Deng, X.; Chai, L. Reproduction in Woody Perennial Citrus: An Update on Nucellar Embryony and Self-Incompatibility. Plant Reprod. 2018, 31, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Denna, D.W. The Potential Use of Self-Incompatibility for Breeding F1 Hybrids of Naturally Self-Pollinated Vegetable Crops. Euphytica 1971, 20, 542–548. [Google Scholar] [CrossRef]
- Brewbaker, J.L.; Gorrez, D.D. Genetics of Self-Incompatibility in the Monocot Genera, Ananas (Pineapple) and Gasteria. Am. J. Bot. 1967, 54, 611–616. [Google Scholar] [CrossRef]
- Goldway, M.; Stern, R.; Zisovich, A.; Raz, A.; Sapir, G.; Schnieder, D.; Nyska, R. The Self-Incompatibility Fertilization System in Rosaceae: Agricultural and Genetic Aspects. Acta Hortic. 2012, 77–82. [Google Scholar] [CrossRef]
- Durand, E.; Chantreau, M.; Veve, A.L.; Stetsenko, R.; Dubin, M.; Genete, M.; Llaurens, V.; Poux, C.; Roux, C.; Billiard, S.; et al. Evolution of Self-Incompatibility in the Brassicaceae: Lessons from a Textbook Example of Natural Selection. Evol. Appl. 2020, 13, 1279–1297. [Google Scholar] [CrossRef] [Green Version]
- Higashiyama, T. Peptide Signaling in Pollen–Pistil Interactions. Plant Cell Physiol. 2010, 51, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClure, B.; Cruz-García, F.; Romero, C. Compatibility and Incompatibility in S-RNase-Based Systems. Ann. Bot. 2011, 108, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Lanaud, C.; Fouet, O.; Legavre, T.; Lopes, U.; Sounigo, O.; Eyango, M.C.; Mermaz, B.; Da Silva, M.R.; Loor Solorzano, R.G.; Argout, X.; et al. Deciphering the Theobroma Cacao Self-Incompatibility System: From Genomics to Diagnostic Markers for Self-Compatibility. J. Exp. Bot. 2017, 68, 4775–4790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.G.; Huang, S.; Li, S.; Staiger, C.J.; Franklin-Tong, V.E. Actin Depolymerization Is Sufficient to Induce Programmed Cell Death in Self-Incompatible Pollen. J. Cell Biol. 2006, 174, 221–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, K.A.; Poulter, N.S.; Franklin-Tong, V.E. Taking One for the Team: Self-Recognition and Cell Suicide in Pollen. J. Exp. Bot. 2014, 65, 1331–1342. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-L.; Wu, J.; Xu, G.-H.; Gao, Y.-B.; Chen, G.; Wu, J.-Y.; Wu, H.-Q.; Zhang, S.-L. S-RNase Disrupts Tip-Localized Reactive Oxygen Species and Induces Nuclear DNA Degradation in Incompatible Pollen Tubes of Pyrus Pyrifolia. J. Cell Sci. 2010, 123, 4301–4309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Meng, D.; Gu, Z.; Yang, Q.; Yuan, H.; Li, Y.; Chen, Q.; Yu, J.; Liu, C.; Li, T. Apple S-RNase Triggers Inhibition of TRNA Aminoacylation by Interacting with a Soluble Inorganic Pyrophosphatase in Growing Self-Pollen Tubes in Vitro. New Phytol. 2018, 218, 579–593. [Google Scholar] [CrossRef]
- Soost, R.K. Incompatibility Alleles in the Genus Citrus. Proc. Amer. Soc. Hort. Sci. 1965, 87, 176–180. [Google Scholar]
- JungHee, K.; Mori, T.; Wakana, A.; BinhXuan, N.; Masuda, J.; Sakai, K.; Kajiwara, K. Production of Homozygous S1 Seedlings for S Gene in “Hirado Buntan” Pummelo (Citrus Grandis osbeck) and Determination of the S Alleles (S9 and S10) by Pollination with the S1 Seedlings to Citrus Cultivars. J. Fac. Agric. Kyushu Univ. 2010, 55, 239–245. [Google Scholar]
- Yamamoto, M.; Kubo, T.; Tominaga, S. Self- and Cross-Incompatibility of Various Citrus Accessions. J. Jpn. Soc. Hortic. Sci. 2006, 75, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Mori, T.; Wakana, A.; Ngo, B.X.; Sakai, K.; Kajiwara, K. Determination of Self-Incompatible Citrus Cultivars with S1 and/or S2 Alleles by Pollination with Homozygous S1 Seedlings (S1S1 or S2S2) of ‘Banpeiyu’ Pummelo. J. Jpn. Soc. Hortic. Sci. 2011, 80, 404–413. [Google Scholar] [CrossRef] [Green Version]
- Ngo, B.X.; Kim, J.-H.; Wakana, A.; Isshiki, S.; Mori, T. Estimation of Self-Incompatibility Genotypes of Citrus Cultivars with Got-3 Allozyme Markers. J. Jpn. Soc. Hortic. Sci. 2011, 80, 284–294. [Google Scholar] [CrossRef]
- Wakana, A.; Ngo, B.; Fukudome, I.; Kajiwara, K. Estimation of the Degree of Self-Incompatibility Reaction during Flower Bud Development and Production of Self-Fertilized Seeds by Bud Pollination in Self-Incompatible Citrus Cultivars. J. Fac. Agric. Kyushu Univ. 2004, 49, 307–320. [Google Scholar] [CrossRef]
- Zhou, X.-H.; Kim, J.-H.; Wakana, A.; Sakai, K.; Kajiwara, K.; Mizunoe, Y. Distribution and Evolution of Citrus with S4and/or S5gene Alleles for Self-Incompatibility with Special Focus on the Origin of Satsuma Mandarin (Citrus Unshiu marc.; SfS4). Genet. Resour. Crop Evol. 2018, 65, 1013–1033. [Google Scholar] [CrossRef]
- Kim, J.H.; Handayani, E.; Wakana, A.; Sato, M.; Miyamoto, M.; Miyazaki, R.; Zhou, X.; Sakai, K.; Mizunoe, Y.; Shigyo, M.; et al. Distribution and Evolution of Citrus Accessions with S3 and/or S11 Alleles for Self-Incompatibility with an Emphasis on Sweet Orange [Citrus Sinensis (L.) Osbeck; Sf S3 or Sf S3sm]. Genet. Resour. Crop Evol. 2020, 67, 2101–2117. [Google Scholar] [CrossRef]
- Gentile, A.; Antognoni, F.; Iorio, R.A.; Distefano, G.; Las Casas, G.; La Malfa, S.; Serafini-Fracassini, D.; Del Duca, S. Polyamines and Transglutaminase Activity Are Involved in Compatible and Self-Incompatible Pollination of Citrus Grandis. Amino Acids 2012, 42, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Uchida, A.; Abe, A.; Yamamoto, A.; Hirano, T.; Kunitake, H. Effects of Polyamines on Self-Incompatibility-like Responses in Pollen Tubes of Citrus Cultivars, Banpeiyu and Hyuganatsu. J. Am. Soc. Hortic. Sci. 2015, 140, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Aloisi, I.; Distefano, G.; Antognoni, F.; Potente, G.; Parrotta, L.; Faleri, C.; Gentile, A.; Bennici, S.; Mareri, L.; Cai, G.; et al. Temperature-Dependent Compatible and Incompatible Pollen-Style Interactions in Citrus Clementina hort. Ex Tan. Show Different Transglutaminase Features and Polyamine Pattern. Front. Plant Sci. 2020, 11, 1018. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.; Merelo, P.; Distefano, G.; La Malfa, S.; Lo Piero, A.R.; Tadeo, F.R.; Talon, M.; Gentile, A. Comparative Transcriptome Analysis of Stylar Canal Cells Identifies Novel Candidate Genes Implicated in the Self-Incompatibility Response of Citrus Clementina. BMC Plant Biol. 2012, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.; Yang, W.; Su, S.; Fu, L.; Yi, H.; Chen, C.; Deng, X.; Chai, L. Genome-Wide Identification and Functional Analysis of S-RNase Involved in the Self-Incompatibility of Citrus. Mol. Genet. Genom. MGG 2017, 292, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Ye, Z.; da Silva, J.A.T.; Qin, Y.; Hu, G. Identifying Differentially Expressed Genes in Pollen from Self-Incompatible “Wuzishatangju” and Self-Compatible “Shatangju” Mandarins. Int. J. Mol. Sci. 2013, 14, 8538–8555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Ding, F.; He, X.; Luo, C.; Huang, G.; Hu, Y. Characterization of the ‘Xiangshui’ Lemon Transcriptome by de Novo Assembly to Discover Genes Associated with Self-Incompatibility. Mol. Genet. Genom. 2015, 290, 365–375. [Google Scholar] [CrossRef]
- Liang, M.; Cao, Z.; Zhu, A.; Liu, Y.; Tao, M.; Yang, H.; Xu, Q.; Wang, S.; Liu, J.; Li, Y.; et al. Evolution of Self-Compatibility by a Mutant S m -RNase in Citrus. Nat. Plants 2020, 6, 131–142. [Google Scholar] [CrossRef]
- Honsho, C.; Ushijima, K.; Anraku, M.; Ishimura, S.; Yu, Q.; Gmitter, F.G.J.; Tetsumura, T. Association of T2/S-RNase With Self-Incompatibility of Japanese Citrus Accessions Examined by Transcriptomic, Phylogenetic, and Genetic Approaches. Front. Plant Sci. 2021, 12, 121. [Google Scholar] [CrossRef]
- Barry, G.H.; Gmitter, F.G., Jr.; Chen, C.; Roose, M.L.; Federici, C.T.; McCollum, G.T. Investigating the Parentage of “Orri” and “Fortune” Mandarin Hybrids. Acta Hortic. 2015, 449–456. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Furr, J.R. New Tangerines for the Desert. Calif. Citrogr. 1964, 49, 266–276. [Google Scholar]
- Hodgson, R.W. Horticultural varieties of citrus. In The Citrus Industry; UC Press: Berkeley, CA, USA; Los Angeles, CA, USA, 1967; Volume 1, pp. 431–591. [Google Scholar]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Légaré, G.; Boyle, B.; Normandeau, É.; Laroche, J.; Larose, S.; Jean, M.; et al. An Improved Genotyping by Sequencing (GBS) Approach Offering Increased Versatility and Efficiency of SNP Discovery and Genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef] [Green Version]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q.; Buckler, E.S. TASSEL-GBS: A High Capacity Genotyping by Sequencing Analysis Pipeline. PLoS ONE 2014, 9, e90346. [Google Scholar] [CrossRef]
- Ollitrault, P.; Terol, J.; Chen, C.; Federici, C.T.; Lotfy, S.; Hippolyte, I.; Ollitrault, F.; Berard, A.; Chauveau, A.; Cuenca, J.; et al. A Reference Genetic Map of C. Clementina Hort. Ex Tan.; Citrus Evolution Inferences from Comparative Mapping. BMC Genom. 2012, 13, 1–20. [Google Scholar] [CrossRef]
- Guo, F.; Yu, H.; Tang, Z.; Jiang, X.; Wang, L.; Wang, X.; Xu, Q.; Deng, X. Construction of a SNP-Based High-Density Genetic Map for Pummelo Using RAD Sequencing. Tree Genet. Genomes 2015, 11, 2. [Google Scholar] [CrossRef]
- Curtolo, M.; Cristofani-Yaly, M.; Gazaffi, R.; Takita, M.A.; Figueira, A.; Machado, M.A. QTL Mapping for Fruit Quality in Citrus Using DArTseq Markers. BMC Genom. 2017, 18, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Roose, M.L.; Yu, Q.; Du, D.; Yu, Y.; Zhang, Y.; Deng, Z.; Stover, E.; Gmitter, F.G.J. Construction of High-Density Genetic Maps and Detection of QTLs Associated With Huanglongbing Tolerance in Citrus. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- van Os, H.; Stam, P.; Visser, R.G.; van Eck, H.J. SMOOTH: A Statistical Method for Successful Removal of Genotyping Errors from High-Density Genetic Linkage Data. TAG Theoretical. Appl. Genet. Angew. Genet. 2005, 112, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An Information Aesthetic for Comparative Genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasche, H.; Hiltemann, S. Galactic Circos: User-Friendly Circos Plots within the Galaxy Platform. GigaScience 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Storey, J.D. A Direct Approach to False Discovery Rates. J. R. Stat. Soc. Ser. B Stat. Methodol. 2002, 64, 479–498. [Google Scholar] [CrossRef] [Green Version]
- Oueslati, A.; Salhi-Hannachi, A.; Luro, F.; Vignes, H.; Mournet, P.; Ollitrault, P. Genotyping by Sequencing Reveals the Interspecific C. Maxima/C. Reticulata Admixture along the Genomes of Modern Citrus Varieties of Mandarins, Tangors, Tangelos, Orangelos and Grapefruits. PLoS ONE 2017, 12, e0185618. [Google Scholar] [CrossRef] [Green Version]
- do Amaral, M.; Barbosa de Paula, M.F.; Ollitrault, F.; Rivallan, R.; de Andrade Silva, E.M.; da Silva Gesteira, A.; Luro, F.; Garcia, D.; Ollitrault, P.; Micheli, F. Phylogenetic Origin of Primary and Secondary Metabolic Pathway Genes Revealed by C. Maxima and C. Reticulata Diagnostic SNPs. Front. Plant Sci. 2019, 10, 1128. [Google Scholar] [CrossRef]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J.; et al. Sequencing of Diverse Mandarin, Pummelo and Orange Genomes Reveals Complex History of Admixture during Citrus Domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Ollitrault, P.; Terol, J.; Garcia-Lor, A.; Bérard, A.; Chauveau, A.; Froelicher, Y.; Belzile, C.; Morillon, R.; Navarro, L.; Brunel, D.; et al. SNP Mining in C. Clementina BAC End Sequences; Transferability in the Citrus Genus (Rutaceae), Phylogenetic Inferences and Perspectives for Genetic Mapping. BMC Genom. 2012, 13, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernet, G.; Fernandez-Ribacoba, J.; Carbonell, E.; Asins, M. Comparative Genome-Wide Segregation Analysis and Map Construction Using a Reciprocal Cross Design to Facilitate Citrus Germplasm Utilization. Mol. Breed. 2010, 659–673. [Google Scholar] [CrossRef]
- Yu, Y.; Chen, C.; Gmitter, F.G. QTL Mapping of Mandarin (Citrus Reticulata) Fruit Characters Using High-Throughput SNP Markers. Tree Genet. Genomes 2016, 12, 77. [Google Scholar] [CrossRef]
- Chen, C.X.; Bowman, K.D.; Choi, Y.A.; Dang, P.M.; Rao, M.N.; Huang, S.; Soneji, J.R.; McCollum, T.G.; Gmitter, F.G., Jr. EST-SSR Genetic Maps for Citrus Sinensis and Poncirus Trifoliata. Tree Genet. Genomes 2008, 4, 1–10. [Google Scholar] [CrossRef]
- Calvez, L.; Dereeper, A.; Mournet, P.; Froelicher, Y.; Bruyère, S.; Morillon, R.; Ollitrault, P. Intermediate Inheritance with Disomic Tendency in Tetraploid Intergeneric Citrus × Poncirus Hybrids Enhances the Efficiency of Citrus Rootstock Breeding. Agronomy 2020, 10, 1961. [Google Scholar] [CrossRef]
- Bowman, F.T. Citrus Growing in Australia; Halstead Press: Sydney, New South Wales, Australia, 1956; p. 311. [Google Scholar]
- Carlos de Oliveira, A.; Bastianel, M.; Cristofani-Yaly, M.; Morais do Amaral, A.; Machado, M.A. Development of Genetic Maps of the Citrus Varieties “Murcott” Tangor and “Pera” Sweet Orange by Using Fluorescent AFLP Markers. J. Appl. Genet. 2007, 48, 219–231. [Google Scholar] [CrossRef]
- Distefano, G.; Hedhly, A.; Las Casas, G.; La Malfa, S.; Herrero, M.; Gentile, A. Male–Female Interaction and Temperature Variation Affect Pollen Performance in Citrus. Sci. Hortic. 2012, 140, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Distefano, G.; Gentile, A.; Hedhly, A.; La Malfa, S. Temperatures during Flower Bud Development Affect Pollen Germination, Self-Incompatibility Reaction and Early Fruit Development of Clementine (Citrus Clementina hort. Ex Tan.). Plant Biol. Stuttg. Ger. 2018, 20, 191–198. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sc\LG | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | Mks/Sc |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 150 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 152 |
| 2 | 0 | 349 | 0 | 4 | 0 | 0 | 1 | 0 | 0 | 354 |
| 3 | 0 | 1 | 351 | 1 | 0 | 0 | 0 | 0 | 0 | 353 |
| 4 | 0 | 0 | 0 | 328 | 0 | 0 | 12 | 0 | 0 | 340 |
| 5 | 1 | 0 | 0 | 0 | 132 | 0 | 63 | 0 | 0 | 196 |
| 6 | 0 | 0 | 0 | 1 | 0 | 303 | 0 | 0 | 0 | 304 |
| 7 | 0 | 0 | 0 | 0 | 0 | 0 | 285 | 0 | 0 | 285 |
| 8 | 0 | 0 | 0 | 0 | 3 | 1 | 0 | 54 | 1 | 59 |
| 9 | 0 | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 109 | 113 |
| N | 0 | 0 | 0 | 1 | 0 | 1 | 5 | 1 | 0 | 8 |
| Mks/LG | 151 | 350 | 352 | 335 | 135 | 306 | 370 | 55 | 110 | 2164 |
| LG size cM | 152.4 | 194.5 | 276.4 | 172.6 | 142.3 | 162.1 | 185.2 | 75.9 | 146.9 | 1508.4 |
| Genes | 97 | 242 | 263 | 246 | 88 | 233 | 233 | 43 | 78 | 1523 |
| SC\LG | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | Mks/Sc |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 182 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 185 |
| 2 | 0 | 128 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 131 |
| 3 | 2 | 0 | 227 | 0 | 0 | 0 | 0 | 8 | 3 | 240 |
| 4 | 0 | 0 | 2 | 66 | 0 | 0 | 6 | 0 | 0 | 74 |
| 5 | 0 | 2 | 0 | 0 | 266 | 0 | 6 | 0 | 1 | 275 |
| 6 | 0 | 0 | 0 | 0 | 0 | 59 | 0 | 0 | 1 | 60 |
| 7 | 0 | 0 | 0 | 0 | 1 | 0 | 146 | 0 | 0 | 147 |
| 8 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 105 | 1 | 107 |
| 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 240 | 247 |
| N | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Mks/LG | 185 | 130 | 230 | 66 | 268 | 59 | 158 | 120 | 251 | 1467 |
| LG size cM | 92.3 | 91.2 | 164.9 | 110.3 | 117.0 | 90.2 | 102.6 | 101.2 | 164.6 | 1034.3 |
| Genes | 136 | 98 | 156 | 55 | 171 | 39 | 109 | 70 | 166 | 1000 |
| ‘Fortune’ Mandarin | ‘Ellendale’ Tangor | |||||||
|---|---|---|---|---|---|---|---|---|
| LG | Total Markers | All Gametes | Male Gametes | Female Gametes | Total Markers | All Gametes | Male Gametes | Female Gamete |
| 1 | 151 | 0 | 0 | 0 | 185 | 0 | 0 | 0 |
| 2 | 350 | 1 | 2 | 0 | 130 | 0 | 0 | 0 |
| 3 | 352 | 10 | 1 | 0 | 230 | 8 | 3 | 0 |
| 4 | 335 | 0 | 0 | 0 | 66 | 0 | 0 | 0 |
| 5 | 135 | 0 | 0 | 0 | 268 | 0 | 0 | 0 |
| 6 | 306 | 69 | 255 | 0 | 59 | 2 | 0 | 0 |
| 7 | 370 | 121 | 54 | 0 | 158 | 93 | 93 | 0 |
| 8 | 55 | 0 | 0 | 0 | 120 | 0 | 0 | 0 |
| 9 | 110 | 1 | 1 | 0 | 251 | 0 | 0 | 0 |
| Total | 2164 | 202 | 313 | 0 | 1467 | 103 | 96 | 0 |
| LG | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| Comp. FH1/Hclem | 99.3% | 100.0% | 99.3% | 99.7% | 99.6% | 98.6% | 99.6% | 100.0% | 99.0% |
| Comp FH2/Hclem | 67.7% | 58.6% | 67.7% | 70.8% | 72.7% | 77.2% | 37.6% | 86.4% | 80.6% |
| Rec FH1 | 2 | 0 | 1 | 0 | 0 | 3 | 2 | 0 | 1 |
| Comp EH1/HOr | 77.1% | 54.8% | 78.9% | 77.0% | 85.5% | 79.8% | 97.5% | 99.5% | 84.8% |
| CompEH2/HOr | 98.3% | 99.7% | 97.5% | 98.9% | 97.2% | 97.9% | 82.8% | 60.9% | 98.2% |
| LG | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| Orlando | 98.9% | 99.0% | 97.9% | 99.0% | 98.9% | 99.0% | 97.1% | 97.8% | 98.7% | 98.5% |
| Grapefruit | 95.0% | 97.4% | 90.5% | 85.8% | 96.6% | 83.7% | 89.4% | 57.8% | 55.6% | 85.1% |
| Dancy | 82.3% | 66.3% | 66.1% | 81.1% | 67.0% | 96.1% | 49.4% | 98.0% | 97.3% | 75.7% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ollitrault, P.; Ahmed, D.; Costantino, G.; Evrard, J.-C.; Cardi, C.; Mournet, P.; Perdereau, A.; Froelicher, Y. Segregation Distortion for Male Parents in High Density Genetic Maps from Reciprocal Crosses between Two Self-Incompatible Cultivars Confirms a Gametophytic System for Self-Incompatibility in Citrus. Agriculture 2021, 11, 379. https://doi.org/10.3390/agriculture11050379
Ollitrault P, Ahmed D, Costantino G, Evrard J-C, Cardi C, Mournet P, Perdereau A, Froelicher Y. Segregation Distortion for Male Parents in High Density Genetic Maps from Reciprocal Crosses between Two Self-Incompatible Cultivars Confirms a Gametophytic System for Self-Incompatibility in Citrus. Agriculture. 2021; 11(5):379. https://doi.org/10.3390/agriculture11050379
Chicago/Turabian StyleOllitrault, Patrick, Dalel Ahmed, Gilles Costantino, Jean-Charles Evrard, Celine Cardi, Pierre Mournet, Aude Perdereau, and Yann Froelicher. 2021. "Segregation Distortion for Male Parents in High Density Genetic Maps from Reciprocal Crosses between Two Self-Incompatible Cultivars Confirms a Gametophytic System for Self-Incompatibility in Citrus" Agriculture 11, no. 5: 379. https://doi.org/10.3390/agriculture11050379