Application of Zinc Oxide Nanoparticles and Plant Growth Promoting Bacteria Reduces Genetic Impairment under Salt Stress in Tomato (Solanum lycopersicum L. ‘Linda’)


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Phenotypic Assay
2.3. Genotypic Assay
2.3.1. Isolation of Genomic DNA
2.3.2. Random Amplification of Polymorphic DNA and Coupled Restriction Enzyme Digestion-Random Amplification assays
2.3.3. Electrophoresis
2.3.4. Statistical Analysis
Phonotypic Analysis
Genetic Analysis
3. Results
3.1. Phenotypic Analysis
3.2. Genetic Analysis
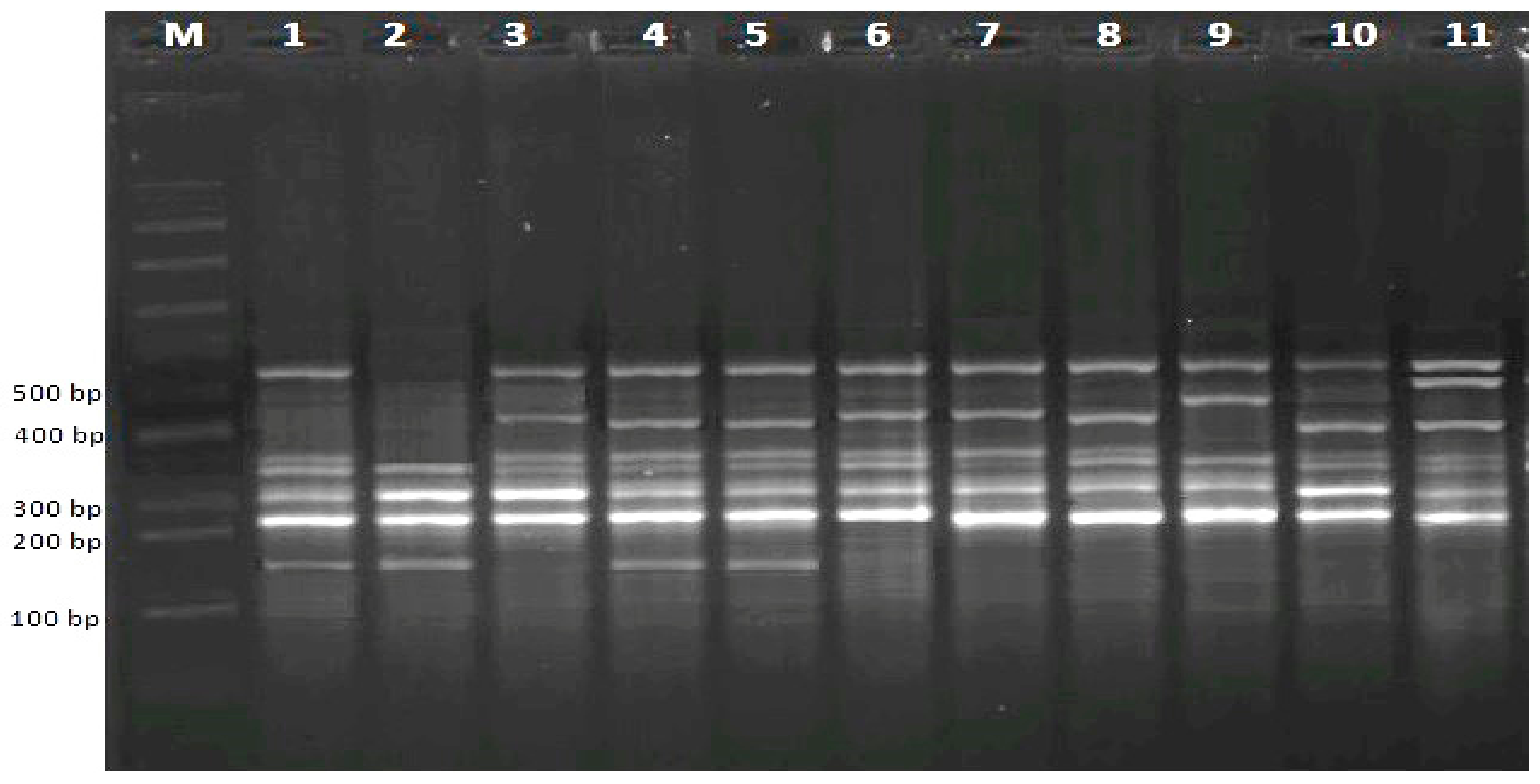
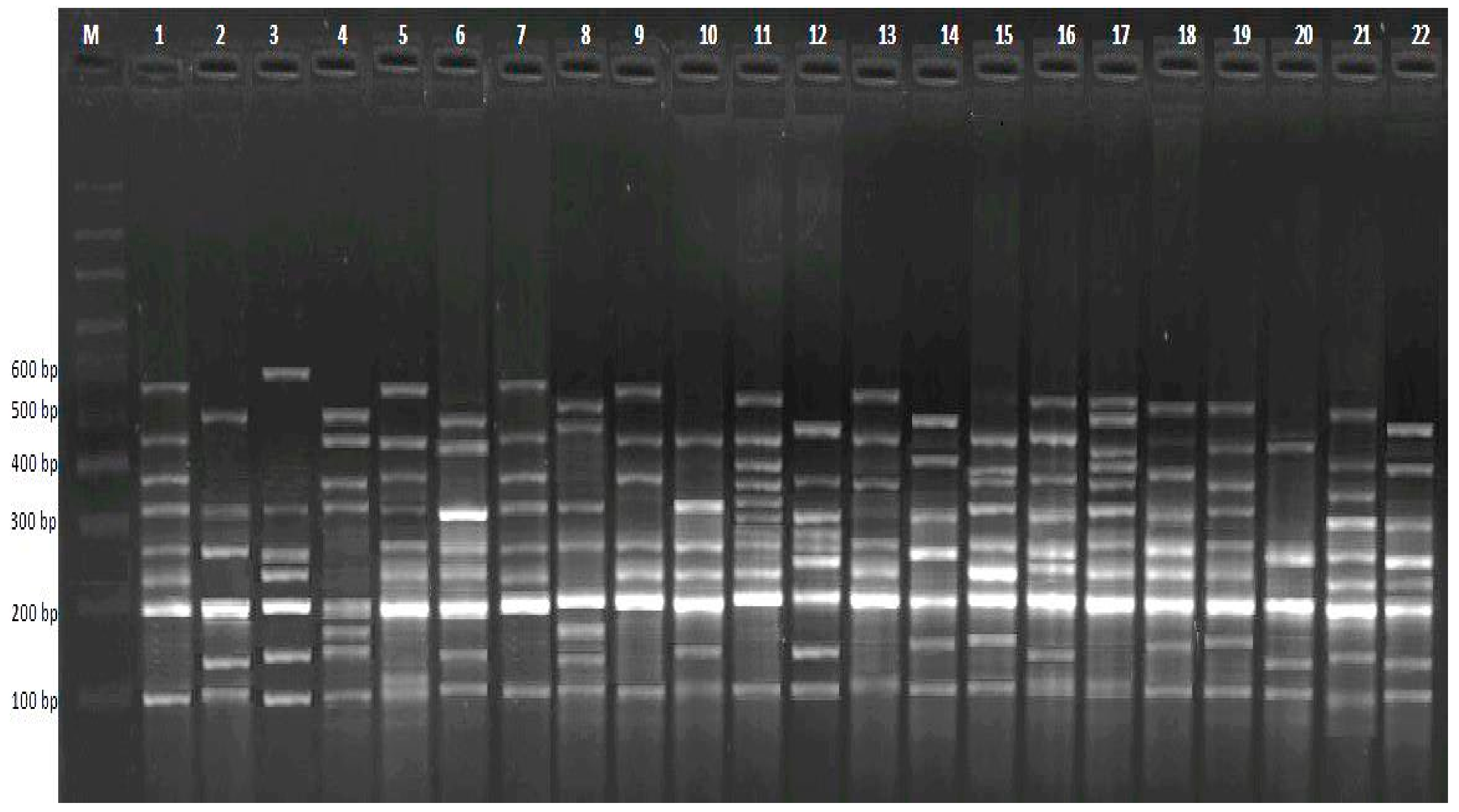
3.2.1. RAPD Analysis
3.2.2. CRED-RA Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben Mrad, M.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef]
- Chen, L.-T.; Luo, M.; Wang, Y.-Y.; Wu, K. Involvement of Arabidopsi s histone deacetylase HDA6 in ABA and salt stress response. J. Exp. Bot. 2010, 61, 3345–3353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigmaz, B.; Agar, G.; Arslan, E.; Aydin, M.; Taspinar, M.S. The role of putrescine against the long terminal repeat (LTR) retrotransposon polymorphisms induced by salinity stress in Triticum aestivum. Acta Physiol. Plant. 2015, 37, 251. [Google Scholar] [CrossRef]
- Kumar, S.; Beena, A.S.; Awana, M.; Singh, A. Salt-induced tissue-specific cytosine methylation downregulates expression of HKT genes in contrasting wheat (Triticum aestivum L.) genotypes. DNA Cell Biol. 2017, 36, 283–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldi, M.; Garofalo, P.; Rubino, P.; Steduto, P. Processing tomatoes under different irrigation regimes in Southern Italy: Agronomic and economic assessments in a simulation case study. J. Agrometeorol. 2011, 3, 39–56. [Google Scholar]
- Wang, Y.; Wisniewski, M.; Meilan, R.; Uratsu, S.L.; Cui, M.; Dandekar, A.; Fuchigami, L. Ectopic expression of Mn-SOD in Lycopersicon esculentum leads to enhanced tolerance to salt and oxidative stress. J. Appl. Hortic. 2007, 9, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Dimkpa, C.; Weinand, T.; Asch, F. Plant–rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 2009, 32, 1682–1694. [Google Scholar] [CrossRef]
- Shilev, S.; Sancho, E.D.; Benlloch-González, M. Rhizospheric bacteria alleviate salt-produced stress in sunflower. J. Environ. Manag. 2012, 95, S37–S41. [Google Scholar] [CrossRef]
- Nair, R.; Varghese, S.H.; Nair, B.G.; Maekawa, T.; Yoshida, Y.; Kumar, D.S. Nanoparticulate material delivery to plants. Plant Sci. 2010, 179, 154–163. [Google Scholar] [CrossRef]
- Sheykh, B.N.; Hasanzadeh, G.T.A.; Baghestani, M.M.; Zand, B. Study the effect of zinc foliar application on the quantitative and qualitative yield of grain corn under water stress. Electron. J. Crop Prod. 2009, 2, 59–73. [Google Scholar]
- Prasad, T.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Reddy, K.R.; Sreeprasad, T.; Sajanlal, P.; Pradeep, T. Effect of nanoscale zinc oxide particles on the germination, growth and yield of peanut. J. Plant Nutr. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Murali Achary, V.M.; Panda, B.B. Aluminium-induced DNA damage and adaptive response to genotoxic stress in plant cells are mediated through reactive oxygen intermediates. Mutagenesis 2009, 25, 201–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardemir, G.; Agar, G.; Arslan, E.; Erturk, F.A. Determination of genetic and epigenetic effects of glyphosate on Triticum aestivum with RAPD and CRED-RA techniques. Theor. Exp. Plant Physiol. 2015, 27, 131–139. [Google Scholar] [CrossRef]
- Saleh, B. DNA changes in cotton (Gossypium hirsutum L.) under salt stress as revealed by RAPD marker. Adv. Hortic. Sci. 2016, 30, 13–21. [Google Scholar]
- Zhang, Q.; Liu, H.; Hu, H.; Li, S.; Ying, Y.; Wu, J. Analysis of the DNA methylation on Camptotheca acuminata decne plants growing in vitro in response to sodium chloride stress. Propag. Ornam. Plants 2014, 14, 76–83. [Google Scholar]
- Erturk, F.A.; Agar, G.; Arslan, E.; Nardemir, G. Analysis of genetic and epigenetic effects of maize seeds in response to heavy metal (Zn) stress. Environ. Sci. Pollut. Res. 2015, 22, 10291–10297. [Google Scholar] [CrossRef]
- Hossein-Pour, A.; Ozkan, G.; Nalci, O.B.; Haliloglu, K. Estimation of genomic instability and DNA methylation due to aluminum (Al) stress in wheat (Triticum aestivum L.) using iPBS and CRED-iPBS analyses. Turk. J. Bot. 2019, 43, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Taspinar, M.S.; Aydin, M.; Arslan, E.; Sigmaz, B.; Agar, G. Salinity and putrescine effects on DNA methylation changes in Triticum aestivum L. J. Biotechnol. 2017, 256, S101. [Google Scholar] [CrossRef]
- Yildirim, N.; Agar, G.; Taspinar, M.S.; Turan, M.; Aydin, M.; Arslan, E. Protective role of humic acids against dicamba-induced genotoxicity and DNA methylation in Phaseolus vulgaris L. Acta Agric. Scand. Sect. B Soil Plant Sci. 2014, 64, 141–148. [Google Scholar]
- Zeinalzadehtabrizi, H.; Hosseinpour, A.; Aydin, M.; Haliloglu, K. A modified genomic DNA extraction method from leaves of sunflower for PCR based analyzes. J. Biodivers. Environ. Sci. 2015, 7, 222–225. [Google Scholar]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki-Ourang, S.; Mehrabi, A.-A.; Siddique, K.H. Screening wild progenitors of wheat for salinity stress at early stages of plant growth: Insight into potential sources of variability for salinity adaptation in wheat. Crop Pasture Sci. 2018, 69, 649–658. [Google Scholar] [CrossRef]
- Ahmadi, J.; Pour-Aboughadareh, A.; Ourang, S.F.; Mehrabi, A.A.; Siddique, K.H. Wild relatives of wheat: Aegilops–Triticum accessions disclose differential antioxidative and physiological responses to water stress. Acta Physiol. Plant. 2018, 40, 90. [Google Scholar] [CrossRef]
- Katsuhara, M.; Kawasaki, T. Salt stress induced nuclear and DNA degradation in meristematic cells of barley roots. Plant Cell Physiol. 1996, 37, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Nouck, A.E.; Taffouo, V.D.; Tsoata, E.; Dibong, D.S.; Nguemezi, S.T.; Gouado, I.; Youmbi, E. Growth, biochemical constituents, micronutrient uptake and yield response of six tomato (Lycopersicum esculentum L.) cultivars grown under salinity stress. Agron. J. 2016, 15, 58–67. [Google Scholar]
- Laware, S.; Raskar, S. Influence of zinc oxide nanoparticles on growth, flowering and seed productivity in onion. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 874–881. [Google Scholar]
- Fathi, A.; Zahedi, M.; Torabian, S.; Khoshgoftar, A. Response of wheat genotypes to foliar spray of ZnO and Fe2O3 nanoparticles under salt stress. J. Plant Nutr. 2017, 40, 1376–1385. [Google Scholar] [CrossRef]
- Ojeda-Barrios, D.L.; Perea-Portillo, E.; Hernández-Rodríguez, O.A.; Ávila-Quezada, G.; Abadía, J.; Lombardini, L. Foliar fertilization with zinc in pecan trees. Hort Sci. 2014, 49, 562–566. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification. Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Jiang, W.; Sun, X.; Xu, H.; Mantri, N.; Lu, H. Optimal concentration of zinc sulfate in foliar spray to alleviate salinity stress in Glycine soja. J. Agric. Sci. Technol. 2014, 16, 445–460. [Google Scholar]
- Marschner, P. Rhizosphere biology. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: London, UK, 2012; pp. 369–388. [Google Scholar]
- Ali, E.; Mahmoud, A.M. Effect of foliar spray by different salicylic acid and zinc concentrations on seed yield and yield components of mungbean in sandy soil. Asian J. Crop Sci. 2013, 5, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Behnajady, M.; Modirshahla, N.; Hamzavi, R. Kinetic study on photocatalytic degradation of CI Acid Yellow 23 by ZnO photocatalyst. J. Hazard. Mater. 2006, 133, 226–232. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Graham, P.; Vance, C. Nitrogen fixation in perspective: An overview of research and extension needs. Field Crop. Res. 2000, 65, 93–106. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R.; Gonzalez, T.; Bashan, Y. Genetics of phosphate solubilization and its potential applications for improving plant growth-promoting bacteria. Plant Soil 2006, 287, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Costacurta, A.; Vanderleyden, J. Synthesis of phytohormones by plant-associated bacteria. Crit. Rev. Microbiol. 1995, 21, 1–18. [Google Scholar] [CrossRef]
- Citterio, S.; Aina, R.; Labra, M.; Ghiani, A.; Fumagalli, P.; Sgorbati, S.; Santagostino, A. Soil genotoxicity assessment: A new strategy based on biomolecular tools and plant bioindicators. Environ. Sci. Technol. 2002, 36, 2748–2753. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.A. Application of DNA (RAPD) and ultrastructure to detect the effect of cadmium stress in Egyptian clover and Sudan grass plantlets. J. Stress Physiol. Biochem. 2012, 8, 241–257. [Google Scholar]
- Tanee, T.; Chadmuk, P.; Sudmoon, R.; Chaveerach, A.; Noikotr, K. Genetic analysis for identification, genomic template stability in hybrids and barcodes of the Vanda species (Orchidaceae) of Thailand. Afr. J. Biotechnol. 2012, 11, 11772–11781. [Google Scholar]
- Peng, H.; Zhang, J. Plant genomic DNA methylation in response to stresses: Potential applications and challenges in plant breeding. Prog. Nat. Sci. 2009, 19, 1037–1045. [Google Scholar] [CrossRef]
- Zhong, L.; Wang, J.-B. The Role of DNA Hypermethylation in Salt Resistence of Triticum aestivum L. J. Wuhan Bot. Res. 2007, 1, 19. [Google Scholar]
- Erturk, F.A.; Aydin, M.; Sigmaz, B.; Taspinar, M.S.; Arslan, E.; Agar, G.; Yagci, S. Effects of As2O3 on DNA methylation, genomic instability, and LTR retrotransposon polymorphism in Zea mays. Environ. Sci. Pollut. Res. 2015, 22, 18601–18606. [Google Scholar] [CrossRef]
- Steward, N.; Ito, M.; Yamaguchi, Y.; Koizumi, N.; Sano, H. Periodic DNA methylation in maize nucleosomes and demethylation by environmental stress. J. Biol. Chem. 2002, 277, 37741–37746. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Rong, T.; Cao, M. Analysis of DNA methylation in different maize tissues. J. Genet. Genom. 2008, 35, 41–48. [Google Scholar] [CrossRef]
- Zhang, C.; Shi, W.; Ma, K.; Li, H.; Zhang, F. EGTA, a calcium chelator, affects cell cycle and increases DNA methylation in root types of Triticum aestivum L. Acta Soc. Bot. Pol. 2016, 85, 3502. [Google Scholar] [CrossRef] [Green Version]
- Bolukbasi, E.; Aras, E. Determination of DNA Methylation Levels with CRED-RA Technique in the Genome of Sunflower Seedlings (Helianthus annuus L.) Subjected to Zinc Stress. Int. J. Environ. Agric. Biotechnol. 2016, 1, 438–444. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′→3′) |
|---|---|
| OPA-13 | CAGCACCCAC |
| OPB-8 | GTCCACACGG |
| OPB-10 | CTGCTGGGAC) |
| OPH-17 | CACTCTCCTC |
| OPH-18 | GAATCGGCCA |
| OPW-4 | CAGAAGCGGA |
| OPW-5 | GGCGGATAAG |
| OPY-8 | AGGCAGAGCA |
| OPY-15 | AGTCGCCCTT |
| OPY-16 | GGGCCAATGT |
| Experimental Group | FLW (cm) 1 | PH (cm) | SD (mm) | LFW (g/plant) | LDW (g/plant) | RFW (g/plant) | RDW (g/plant) |
|---|---|---|---|---|---|---|---|
| Control | 8.5 ± 1.2 e 2 | 17.25 ± 1.6 f | 2.19 ± 0.42 d | 5.72 ± 1.8 ef | 0.68 ± 0.24 ab | 0.64 ± 0.09 ed | 0.1 ± 0.08 e |
| 250 mM NaCl | 5 ± 0.5 e | 12 ± 2 f | 1.78 ± 0.4 e | 3.19 ± 0.8 f | 0.49 ± 0.9 f | 0.41 ± 0.15 d | 0.08 ± 0.02 e |
| Salinity stress + Bacillus pumilus + 0 mg L−1 ZnO-NPs | 21 ± 1.5 abc | 24.3 ± 1.7 de | 4.45 ± 0.23 c | 15.46 ± 5.9 d | 1.68 ± 1.2 de | 1.57 ± 0.7 ef | 0.17 ± 0.12 cde |
| Salinity stress + Bacillus pumilus + 20 mg L−1 ZnO-NPs | 20 ± 3 c | 28.45 ± 0.7 bcd | 4.95 ± 0.23 a | 18.43 ± 3.9 c | 2.78 ± 1.2 bc | 2.17 ± 0.8 be | 0.21 ± 0.81 bc |
| Salinity stress + Bacillus pumilus + 40 mg L−1 ZnO-NPs | 22.1 ± 2.0 bc | 29.75 ± 1.5 cde | 5.37 ± 1.13 a | 20.43 ± 3.1 bc | 3.20 ± 1.7 b | 2.58 ± 1.4 bc | 0.25 ± 0.13 ab |
| Salinity stress + Lactobacillus casei + 0 mg L−1 ZnO-NPs | 24.3 ± 1.2 bc | 33.0 ± 1.3 abc | 4.83 ± 0.93 bc | 21.38 ± 4.5 bc | 2.30 ± 0.6 cd | 3.01 ± 1.2 b | 0.27 ± 0.80 bc |
| Salinity stress + Lactobacillus casei + 20 mg L−1 ZnO-NPs | 26 ± 0.7 b | 32.53 ± 0.76 ab | 5.90 ± 0.42 a | 22.34 ± 3.4 b | 3.25 ± 1.4 ab | 3.10 ± 1.7 ab | 0.29 ± 0.12 ab |
| Salinity stress + Lactobacillus casei + 40 mg L−1 ZnO-NPs | 28 ± 0.9 a | 39.50 ± 0.45 a | 6.12 ± 0.76 a | 24.67 ± 2.7 a | 3.45 ± 0.9 a | 3.19 ± 1.7 a | 0.31 ± 0.7 a |
| Salinity stress + Bacillus subtilis + 0 mg L−1 ZnO-NPs | 11 ± 1.5 d | 18.25 ± 1.8 ef | 3.11 ± 0.29 d | 10.05 ± 1.4 f | 1.34 ± 1.1 def | 1.09 ± 0.2 ed | 0.08 ± 0.05 e |
| Salinity stress + Bacillus subtilis + 20 mg L−1 ZnO-NPs | 13 ± 5 dc | 18.6 ± 1.8 def | 3.6 ± 0.31 bc | 13.50 ± 3.3 de | 1.24 ± 0.14 ef | 1.26 ± 0.4 def | 0.15 ± 0.03 de |
| Salinity stress + Bacillus subtilis + 40 mg L−1 ZnO-NPs | 18.5 ± 2.2 c | 26.5 ± 1.4 de | 4.41 ± 0.94 c | 14.95 ± 2.3 d | 1.45 ± 0.3 def | 1.28 ± 0.2 cf | 0.18 ± 0.1 de |
| F value | 17.70 ** | 29.38 ** | 9.31 ** | 24.00 ** | 14.20 ** | 11.06 ** | 8.85 ** |
| Primer | ± | Control | Experimental Group | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 250 mM NaCl | Salinity Stress + Bacillus pumilus | Salinity Stress + Lactobacillus Casei | Salinity Stress + Bacillus subtilis | |||||||||
| 0 mgL−1 ZnO-NPs | 20 mgL−1 ZnO-NPs | 40 mgL−1 ZnO-NPs | 0 mgL−1 ZnO-NPs | 20 mgL−1 ZnO-NPs | 40 mgL−1 ZnO-NPs | 0 mgL−1 ZnO-NP | 20 mgL−1 ZnO-NPs | 40 mgL−1 ZnO-NPs | ||||
| OPA-13 | + | 11 | 520; 140 | - | 518; 434 | - | 487; 175 | 500 | - | 175 | - | - |
| − | 635; 396 | 665 | - | 665 | 665; 587 | 665 | 665; 587 | 665; 587; 539 | 665; 202; 110 | 665; 587 | ||
| OPB-8 | + | 7 | 635 | 537; 274 | 534 | 547 | 366; 300 | 500 | - | 534 | 491 | - |
| − | 390 | 173 | 377 | 306 | - | 377 | - | - | - | - | ||
| OPB-10 | + | 5 | 375 | 348 | - | - | - | - | - | - | - | - |
| − | 443; 325; 278 | - | - | - | - | - | - | 274 | - | - | ||
| OPH-17 | + | 6 | - | 414 | 400 | 400 | 414 | 414 | 406 | 428 | 400 | 510; 410 |
| − | 530; 340 | 147 | - | - | 147 | 147 | 147 | 340; 147 | 147 | 147 | ||
| OPH-18 | + | 7 | 372 | - | - | - | 664 | 355 | - | - | - | - |
| − | 420; 392; 125 | 328; 291 | 291 | - | - | - | - | 709; 451 | - | - | ||
| OPW-4 | + | 5 | 337 | 317; 192 | 693; 400; 271; 163 | 427; 311 | 486; 200 | 422 | 490 | 486 | 427; 176 | 486; 264 |
| − | 585; 395 | 585 | - | - | - | - | - | 633; 536 | 536 | 633 | ||
| OPW-5 | + | 7 | 665; 428 | 464; 167 | 748; 152 | 484; 143 | 571; 109 | 484; 134 | - | 588; 148 | 420; 143 | 148 |
| − | 120 | 258 | - | - | - | - | - | - | 258 | - | ||
| OPY-8 | + | 5 | - | 269; 211; 145 | 426; 300; 206; 161 | 339; 291; 222; 100 | 434; 288; 195; 122 | 400; 291; 200 | 393; 278; 228; 167 | 300; 248; 195 | 282; 222; 183 | 274; 167 |
| − | 445; 372 | - | - | 580 | - | - | - | - | - | - | ||
| OPY-15 | + | 9 | 639 | - | 554 | 588; 193; 147 | - | 558; 152 | - | 660 | 617 | 617 |
| − | 410; 322 | 400; 359 | 359; 119 | 359 | 446; 359 | 359 | 446; 359 | 400; 227; 119 | 537 | 169; 119 | ||
| OPY-16 | + | 9 | 428 | 215; 149 | 164 | - | - | 215 | 416 | - | - | - |
| − | 395; 252 | 525 | 368 | 292 | - | 368 | 586; 118 | 586; 525; 246 | 586 | 586; 525 | ||
| Total band | 71 | 30 | 23 | 21 | 18 | 19 | 18 | 14 | 26 | 18 | 16 | |
| Polymorphism (%) | 42.26 | 32.39 | 29.58 | 25.35 | 26.76 | 25.35 | 19.72 | 36.62 | 25.35 | 22.54 | ||
| GTS | 57.74 | 67.61 | 70.42 | 74.65 | 73.24 | 74.65 | 80.28 | 63.38 | 74.65 | 77.46 | ||
| Primer | M/H 1 | ± | Control 2 | Experimental Group | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 250 mM NaCl | Salinity Stress + Bacillus pumilus | Salinity Stress + Lactobacillus casei | Salinity Stress + Bacillus subtilis | ||||||||||
| 0 mgL−1 ZnO-NPs | 20 mgL−1 ZnO-NPs | 40 mgL−1 ZnO-NPs | 0 mgL−1 ZnO-NPs | 20 mgL−1 ZnO-NPs | 40 mgL−1 ZnO-NPs | 0 mgL−1 ZnO-NP | 20 mgL−1 ZnO-NPs | 40 mgL−1 ZnO-NPs | |||||
| OPA-13 | M | + | 8 | 552; 356; 252 | - | - | - | 394; 238 | 574;414 | - | - | - | 100, 190 |
| − | 325; 232 | 146 | 464; 115 | 464; 115 | 464 | - | 464; 432 | 464 | 473; 329 | 464 | |||
| H | + | 9 | 225; 182 | 209; 146 | - | - | 190 | - | - | - | - | 120 | |
| − | 585; 382 | 473; 439 | 473; 439 | 473; 329; 126 | 473 | 473; 429 | 473; 329 | 473; 386 | 464; 432 | 473; 429; 386 | |||
| Class | Class II | Class I | Class I | Class I | Class II | Class II | Class I | Class I | Class I | Class II | |||
| OPB-8 | M | + | 7 | 285; 220 | - | 211 | 194 | - | - | 138 | - | - | - |
| − | 650 | 685 | - | - | - | 685; 592 | - | 685 | 669; 126 | 685; 150 | |||
| H | + | 8 | 522 | 367 | - | 353 | 339 | - | 367 | - | - | - | |
| − | 750; 652; 225 | 150; 126 | 669; 600; 150 | 200 | 150; 126 | 126 | 150 | 600; 126 | - | 669; 126 | |||
| Class | Class II | Class I | Class II | Class II | Class I | Class I | Class III | Class I | Class I | Class I | |||
| OPB-10 | M | + | 6 | 485; 335 | 481; 435 | 674; 600; 459 | 623; 476 | 817; 687; 527; 431 | 600; 445 | 487 | 762; 661; 578; 440 | 674; 454; 427 | 578; 435 |
| − | 204 | 144 | 144 | 223; 144 | 223; 144 | 223; 144 | 144 | 223; 144 | - | 223; 144 | |||
| H | + | 5 | 628; 324 | 427 | 762; 600; 427 | 611; 260 | 661; 500; 272 | 440 | 623; 389 | 674; 379 | 635; 431; 372 | 266 | |
| − | - | - | - | - | - | 319 | - | - | 223; 144 | - | |||
| Class | Class IV | Class IV | Class IV | Class III | Class IV | Class I | Class III | Class IV | Class IV | Class III | |||
| OPH-17 | M | + | 6 | 424; 357; 189 | 431; 230 | 533; 167 | 230; 192 | 431; 300; 258 | 431 | 550; 400; 254 | 550; 330; 230 | - | 451; 220 |
| − | 285 | - | - | - | - | - | - | - | 320 | - | |||
| H | + | 8 | 580; 150 | 129 | - | - | 400; 300 | - | 407; 180 | 531; 415 | 170 | 150 | |
| − | 545; 450; 354 | - | - | 320 | - | 320 | - | - | - | - | |||
| Class | Class II | Class IV | Class II | Class II | Class IV | Class II | Class IV | Class IV | Class III | Class IV | |||
| OPH-18 | M | + | 12 | 423; 378; 310 | 223 | 472; 294 | 328; 287: 223 | 142 | 217; 120 | 462; 229 | 558; 467 | 431; 316; 205 | 216; 116 |
| − | 628; 487; 300 | - | - | 517 | - | - | - | - | 628 | - | |||
| H | + | 12 | 776; 505; 319; 257; 157 | 379; 316; 216; 157; 122 | 311; 211 | 305; 242; 205 | 359; 120 | 728; 323; 130 | 350; 229 | 242; 194 | 536; 223 | 200; 130 | |
| − | 309; 116 | - | 629 | 589; 528 | - | - | 589 | 589 | - | 528 | |||
| Class | Class III | Class IV | Class IV | Class IV | Class IV | Class IV | Class IV | Class IV | Class IV | Class IV | |||
| OPW-4 | M | + | 9 | 462; 476; 352 | - | 463; 304 | 454 | 561 | 459 | 250 | 504; 485 | - | - |
| − | 525; 439 | 332 | 565 | - | 332 | 332 | - | 332 | 404; 284; 135 | 593; 227 | |||
| H | + | 11 | 641; 521;402 | - | 528 | - | - | - | - | - | - | - | |
| − | 505; 450; 310 | 345; 284 | 284 | 284; 135 | 505; 284 | 474; 345; 284 | 345; 284; 135 | 345; 284 | - | 622; 505; 284 | |||
| Class | Class II | Class I | Class II | Class II | Class I | Class I | Class II | Class II | Class I | Class I | |||
| OPW-5 | M | + | 12 | 562; 424; 325; 282; 171 | - | 529; 171 | 656 | - | - | 680 | - | - | - |
| − | - | 557; 368 | 395; 303 | 512; 253 | 614; 557; 395; 230 | 614; 368 | 303 | 557; 395; 230 | 614; 557; 475; 409 | 614; 557; 395; 368 | |||
| H | + | 13 | 203 | - | 249 | - | - | - | - | - | - | - | |
| − | 585; 553; 475; 347; 255; 154 | 553; 347; 230 | 679; 553; 525; 409 | 679; 553 | 679; 614; 525; 492; 475; 409 | 492; 475; 409 | 679; 614; 475; 409; 347 | 679; 614; 492; 347 | 230 | 679; 614; 553; 525; 475 | |||
| Class | Class II | Class I | Class I | Class I | Class I | Class I | Class I | Class I | Class I | Class I | |||
| OPY-8 | M | + | 7 | 487; 215 | 706; 607 525; 281 | 613; 255 | 766; 281 | 693; 234; 100 | 712; 268; 214 | 600; 281 | 700; 274 | 500 | - |
| − | 342 | - | 381; 164 | - | 323 | - | 164 | - | - | 657; 562; 492 | |||
| H | + | 9 | 441; 414; 278;189 | 419; 281 | - | 687; 500 | 476; 143 | - | 300 | 508 | 693; 358; 281 | - | |
| − | - | - | 164 | - | 606 | - | 719; 164 | - | - | 719; 651; 606 | |||
| Class | Class IV | Class IV | Class I | Class IV | Class IV | Class I | Class III | Class IV | Class IV | Class I | |||
| OPY-15 | M | + | 5 | 550 | 433; 315 | 480; 414; 323 | 386; 358 | 344 | 364; 321 | 413; 344; 233 | 354; 307; 270 | 433; 347; 239 | 331 |
| − | 512; 386 | 542 | - | - | 491; 195 | 542 | - | - | - | 542 | |||
| H | + | 4 | - | 420; 358; 262 | 240 | 350; 243 | 427 | 413; 336 | 413; 355; 232 | 463; 413; 315; 232 | 362; 307; 206 | 354; 307 | |
| − | 579; 180 | 184 | 513 | - | 513; 184 | 184 | - | - | - | - | |||
| Class | Class I | Class IV | Class II | Class IV | Class I | Class IV | Class I | Class IV | Class IV | Class II | |||
| OPY-16 | M | + | 6 | - | 668; 400 | 280; 247 | 529; 405; 239 | 669 | 704 | 641; 474; 264 | 665; 509; 280; 247 | 509; 452; 224 | - |
| − | 519; 422; 375; 227 | 569 | 449 | - | 449 | - | - | - | 375 | 569 | |||
| H | + | 7 | 503; 454; 221 | 669; 517; 500 | 546; 506 | 557; 465; 351 | - | - | 466; 264 | 646; 448 | 641; 227 | 679 | |
| − | 223 | 339 | 260 | - | 422; 339; 260 | 260 | 339 | 339 | - | 485; 422; 339 | |||
| Class | Class III | Class IV | Class IV | Class IV | Class I | Class II | Class IV | Class IV | Class IV | Class I | |||
| Polymorphism % | M | 53.88 | 39.03 | 38.98 | 32.73 | 36.79 | 35.42 | 30.46 | 40.70 | 36.12 | 33.78 | ||
| H | 51.53 | 37.31 | 30.53 | 30.11 | 35.91 | 29.19 | 30.37 | 34.01 | 29.60 | 33.32 | |||
| Type | Methylation Status | Digestibility by Enzymes | Average (%) Rate of Methylation Class | |
|---|---|---|---|---|
| Restriction Pattern | ||||
| HpaII | MspI | |||
| Class I | CCGG CCGG | Active | Active | 35.00% |
| GGCC GGCC | ||||
| Class II | CCGG | Active | Inactive | 20.00% |
| GGCC | ||||
| Class III | CCGG | Inactive | Active | 8.00% |
| GGCC | ||||
| Class IV | CCGG | Inactive | Inactive | 37.00% |
| GGCC | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosseinpour, A.; Haliloglu, K.; Tolga Cinisli, K.; Ozkan, G.; Ozturk, H.I.; Pour-Aboughadareh, A.; Poczai, P. Application of Zinc Oxide Nanoparticles and Plant Growth Promoting Bacteria Reduces Genetic Impairment under Salt Stress in Tomato (Solanum lycopersicum L. ‘Linda’). Agriculture 2020, 10, 521. https://doi.org/10.3390/agriculture10110521
Hosseinpour A, Haliloglu K, Tolga Cinisli K, Ozkan G, Ozturk HI, Pour-Aboughadareh A, Poczai P. Application of Zinc Oxide Nanoparticles and Plant Growth Promoting Bacteria Reduces Genetic Impairment under Salt Stress in Tomato (Solanum lycopersicum L. ‘Linda’). Agriculture. 2020; 10(11):521. https://doi.org/10.3390/agriculture10110521
Chicago/Turabian StyleHosseinpour, Arash, Kamil Haliloglu, Kagan Tolga Cinisli, Guller Ozkan, Halil Ibrahim Ozturk, Alireza Pour-Aboughadareh, and Peter Poczai. 2020. "Application of Zinc Oxide Nanoparticles and Plant Growth Promoting Bacteria Reduces Genetic Impairment under Salt Stress in Tomato (Solanum lycopersicum L. ‘Linda’)" Agriculture 10, no. 11: 521. https://doi.org/10.3390/agriculture10110521