Contrasting Physiological and Environmental Controls of Evapotranspiration over Kernza Perennial Crop, Annual Crops, and C4 and Mixed C3/C4 Grasslands


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
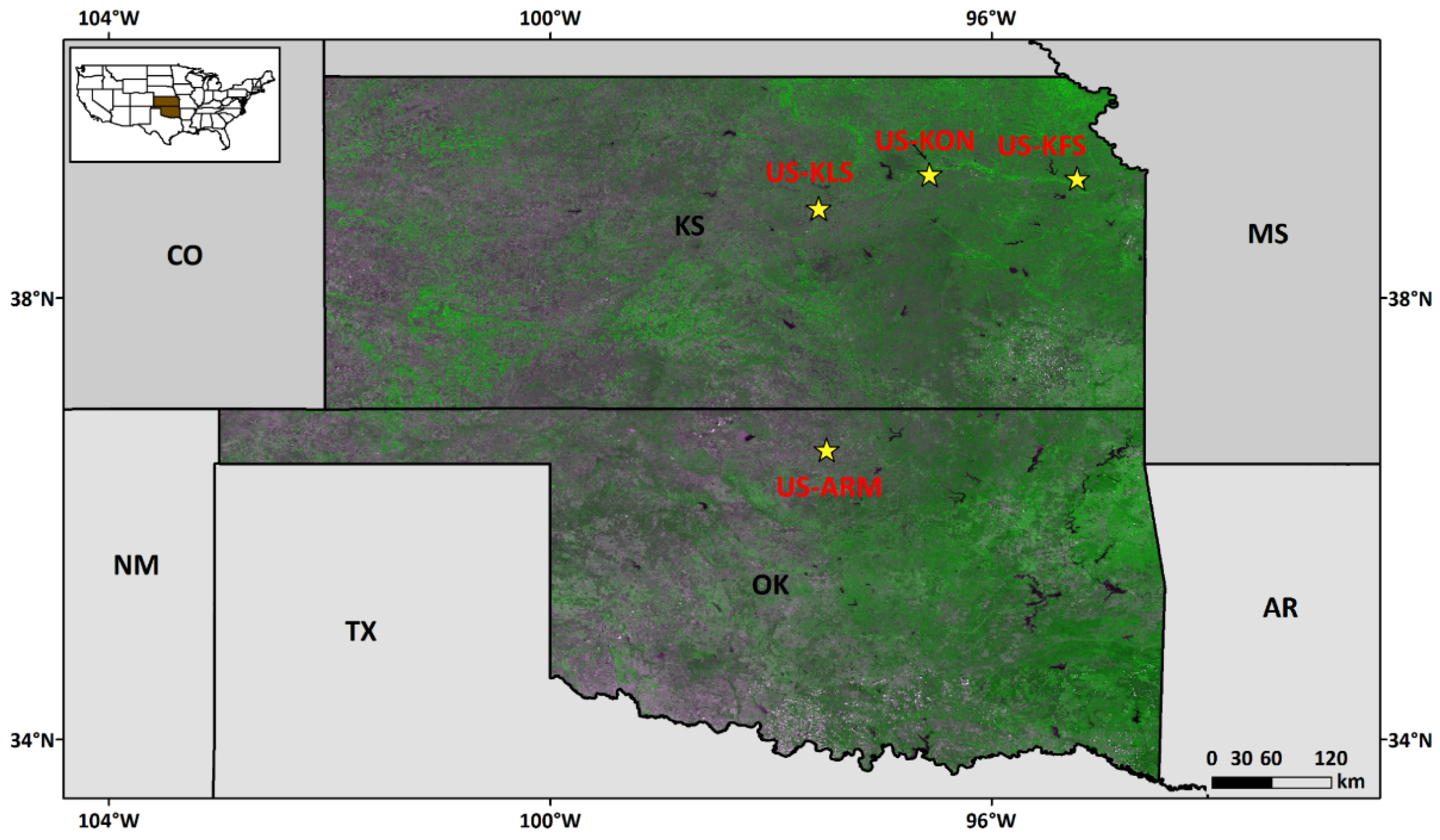
2.1. Sites
2.2. Calculation of the Decoupling Coefficient
2.3. Desing of Research
3. Results and Discussion
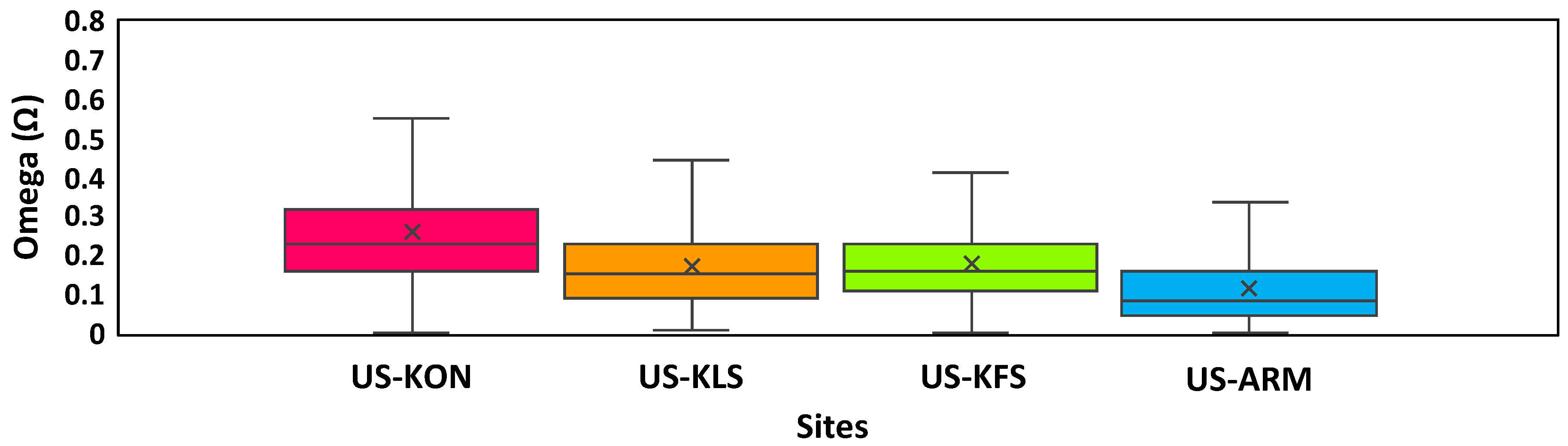
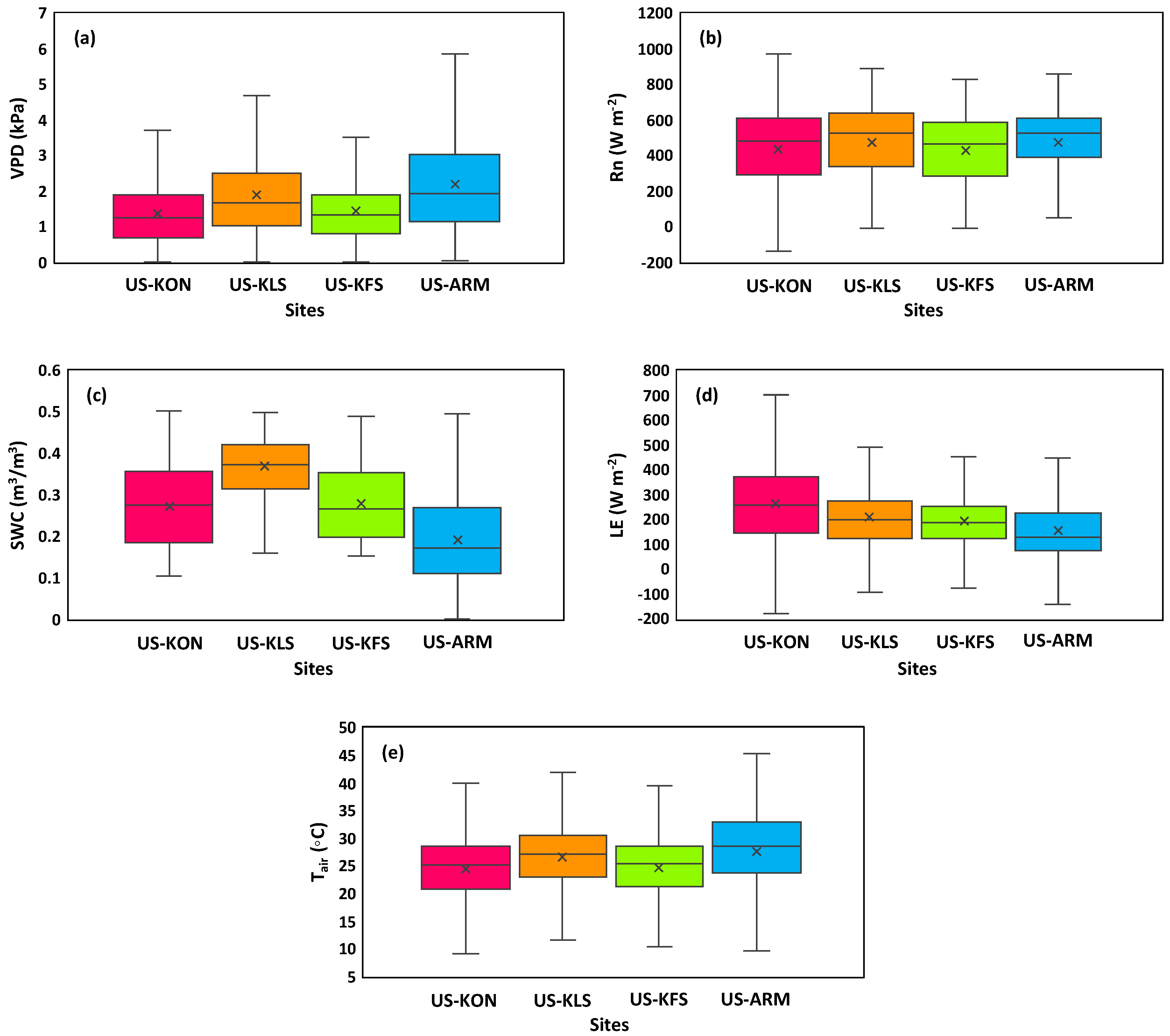
3.1. Site Comparisons
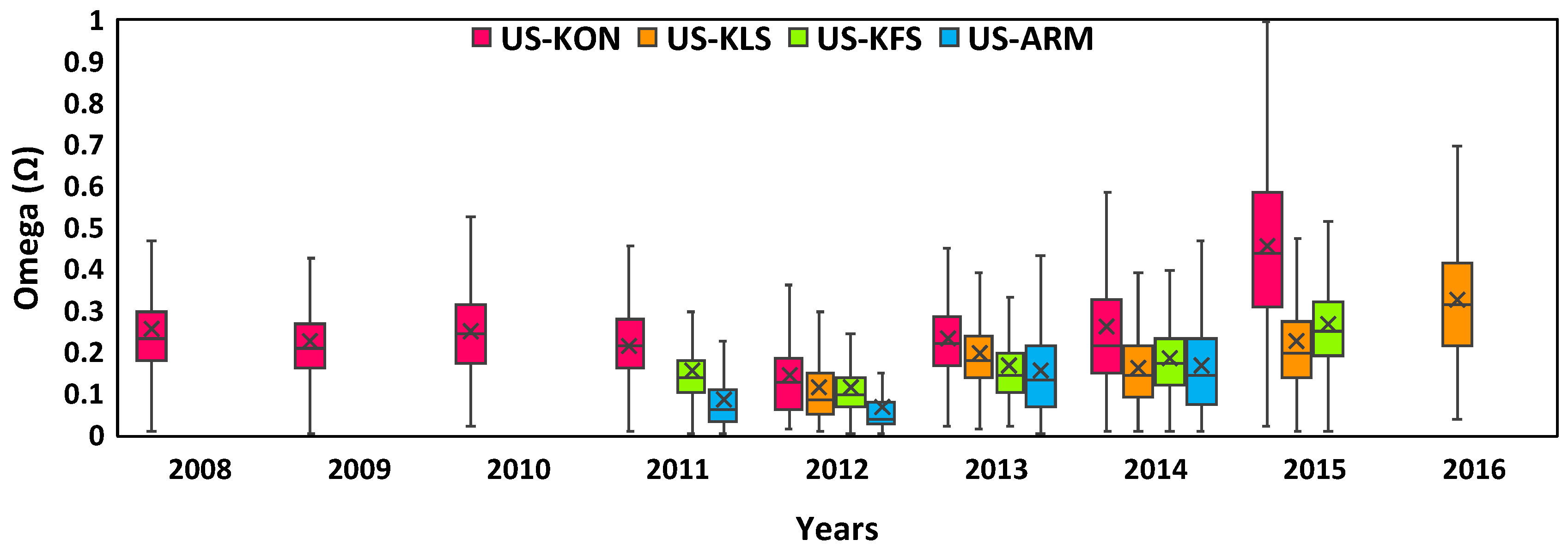
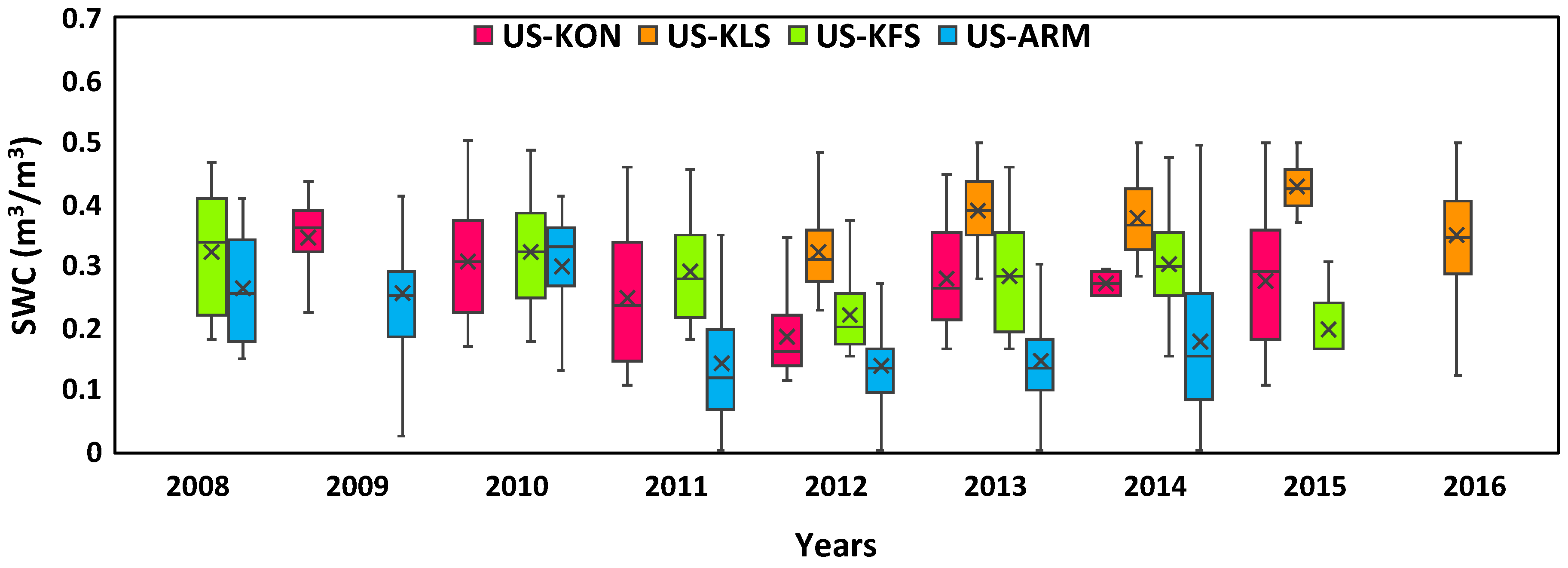
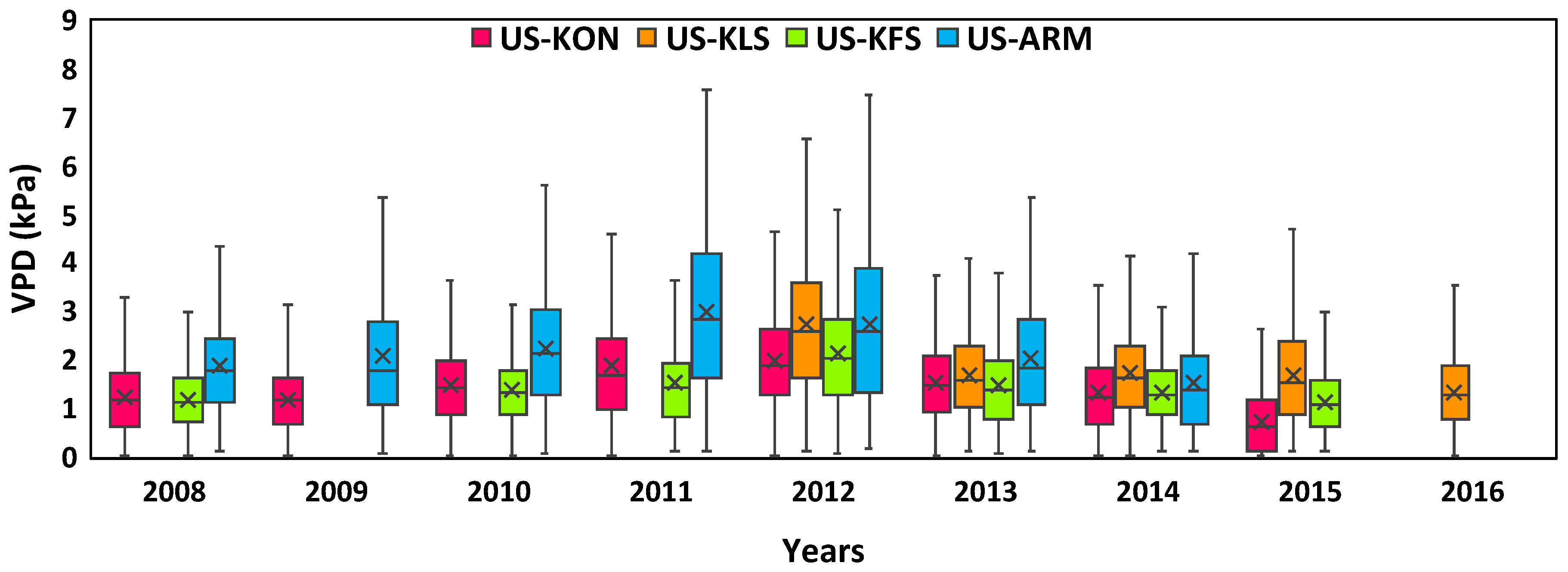
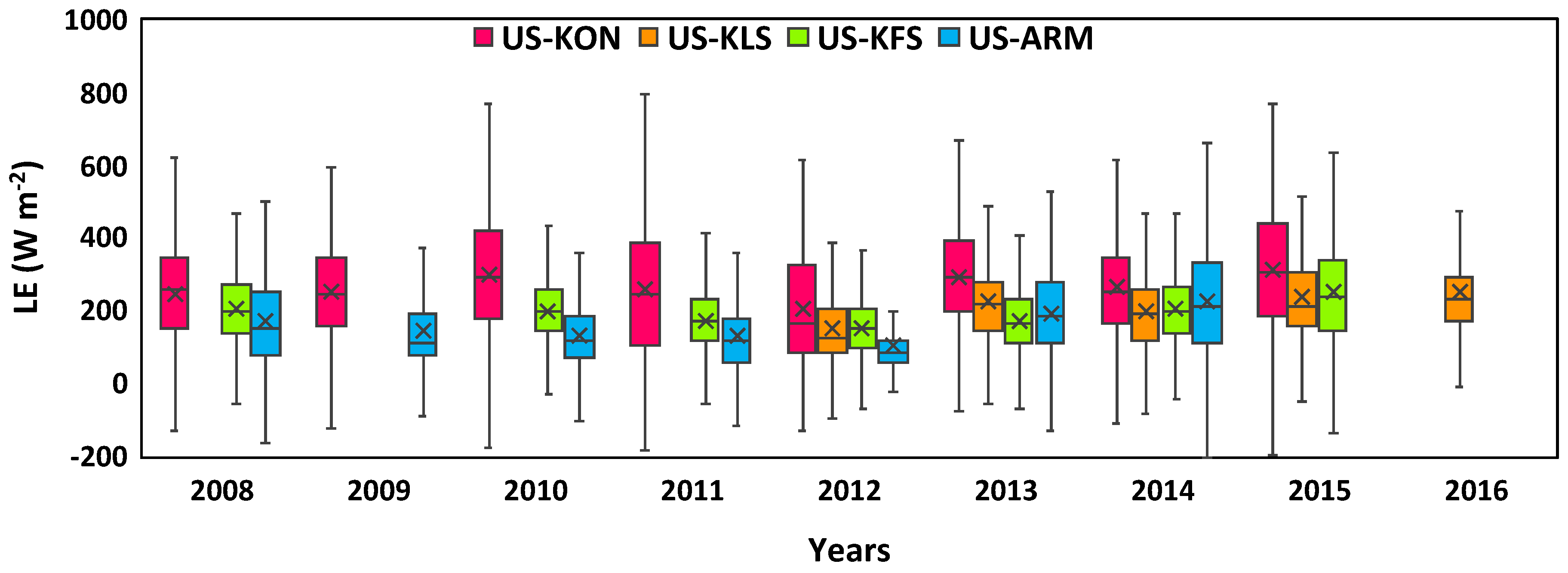
3.2. Annual Fluctuations
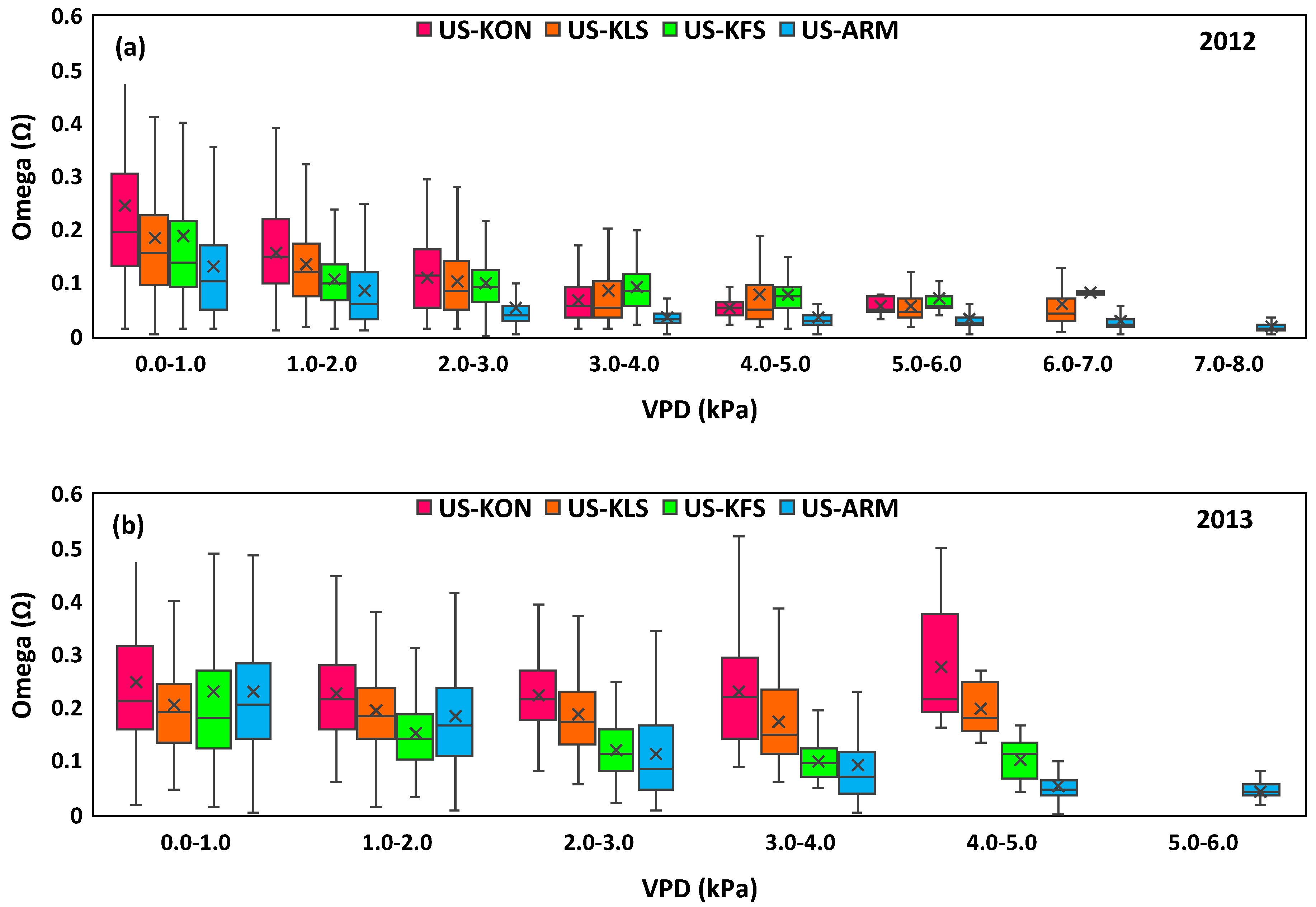
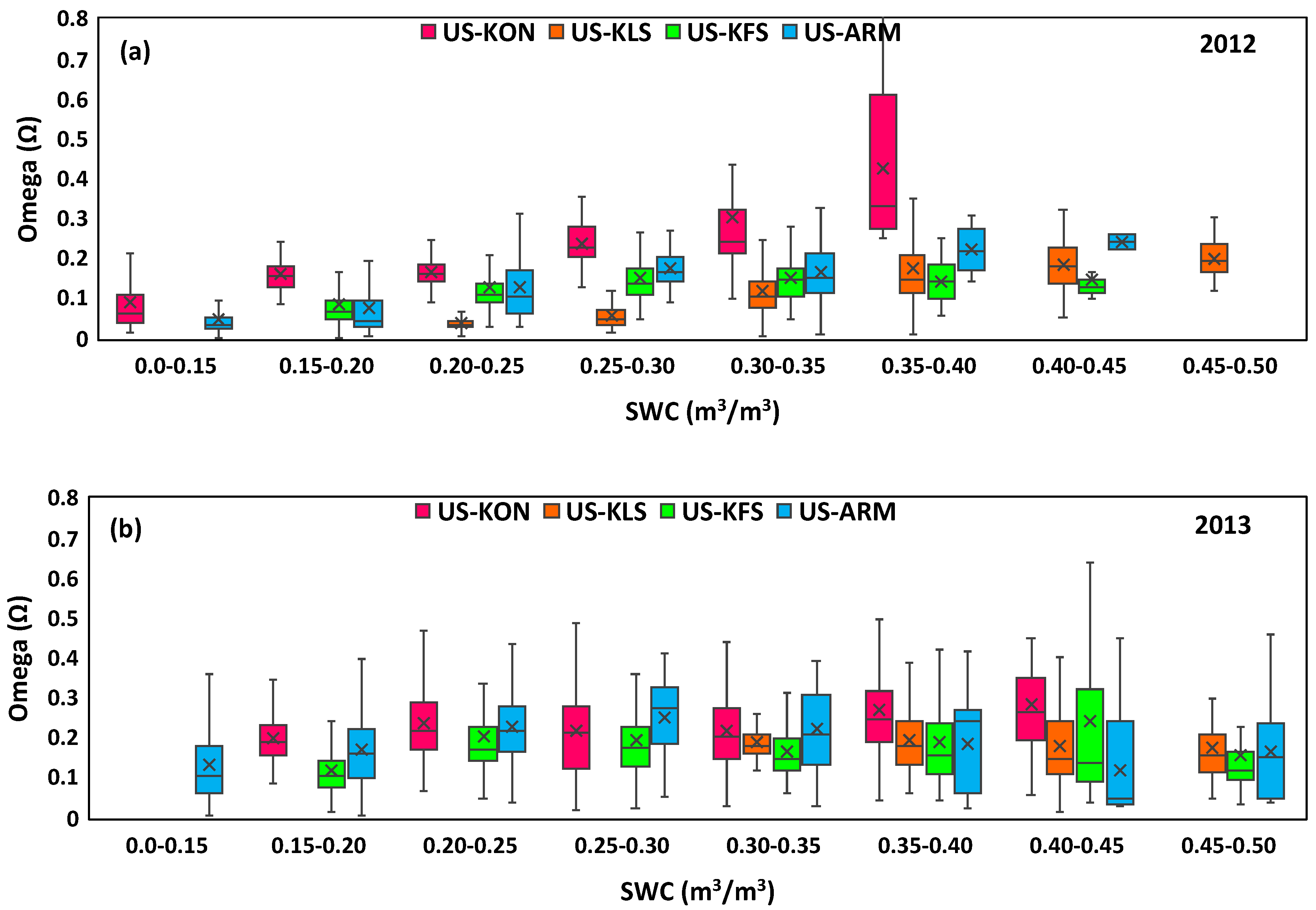
3.3. Water Fluctuation Effects
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pachauri, R.K.; Meyer, L.A. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; 151 pp., ISBN 978-92-9169-143-2. [Google Scholar]
- United Nations. World Population Prospects: The 2017 Revision, Key Findings and Advance Tables; United Nations: New York, NY, USA, 2017; 46 p. [Google Scholar]
- Zhang, Y.; Li, Y.; Jiang, L.; Tian, C.; Li, J.; Xiao, Z. Potential of perennial crop on environmental sustainability of agriculture. Procedia Environ. Sci. 2011, 10, 1141–1147. [Google Scholar] [CrossRef]
- Pimentel, D.; Cerasale, D.; Stanley, R.C.; Perlman, R.; Newman, E.M.; Brent, L.C.; Mullan, A.; Chang, D.T.I. Annual vs. perennial grain production. Agric. Ecosyst. Environ. 2012, 161, 1–9. [Google Scholar] [CrossRef]
- Lobell, D.B.; Bala, G.; Duffy, P.B. Biogeophysical impacts of cropland management changes on climate. Geophys. Res. Lett. 2006, 33, 1–4. [Google Scholar] [CrossRef]
- Findell, K.L.; Berg, A.; Gentine, P.; Krasting, J.P.; Lintner, B.R.; Malyshev, S.; Santanello, J.A.; Shevliakova, E. The impact of anthropogenic land use and land cover change on regional climate extremes. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological consequences of ecosystem fragmentation: A review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Schipanski, M.E.; MacDonald, G.K.; Rosenzweig, S.; Chappell, M.J.; Bennett, E.M.; Kerr, R.B.; Blesh, J.; Crews, T.; Drinkwater, L.; Lundgren, J.G.; et al. Realizing resilient food systems. Bioscience 2016, 66, 600–610. [Google Scholar] [CrossRef]
- Crews, T.E.; Carton, W.; Olsson, L. Is the future of agriculture perennial? Imperatives and opportunities to reinvent agriculture by shifting from annual monocultures to perennial polycultures. Glob. Sust. 2018, 1, 1–18. [Google Scholar] [CrossRef]
- Monfreda, C.; Ramankutty, N.; Foley, J.A. Farming the planet: 2 Geographic distribution of crop areas, yields, physiological types, and net primary production in the year 2000. Glob. Biogeochem. Cycles 2008, 22, 1–19. [Google Scholar] [CrossRef]
- Glover, J.D.; Reganold, J.P.; Bell, L.W.; Borevitz, J.; Brummer, E.C.; Buckler, E.S.; Cox, C.M.; Cox, T.S.; Crews, T.E.; Culman, S.W.; et al. Increased food and ecosystem security via perennial grains. Science 2010, 328, 1638–1639. [Google Scholar] [CrossRef]
- Zeri, M.; Hussain, M.Z.; Anderson-Teixeira, K.J.; DeLucia, E.; Bernacchi, C.J. Water use efficiency of perennial and annual bioenergy crops in central Illinois. J. Geophys. Res.-Biogeo. 2013, 118, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Chimento, C.; Amaducci, S. Characterization of fine root system and potential contribution to soil organic carbon of six perennial bioenergy crops. Biomass Bioenerg. 2015, 83, 116–122. [Google Scholar] [CrossRef]
- Glover, J.D.; Culman, S.W.; DuPont, S.T.; Broussard, W.; Young, L.; Mangan, M.E.; Mai, J.G.; Crews, T.E.; DeHaan, L.R.; Buckley, D.H.; et al. Harvested perennial grasslands provide ecological benchmarks for agricultural sustainability. Agric. Ecosyst. Environ. 2010, 137, 3–12. [Google Scholar] [CrossRef]
- Crews, T.E.; DeHaan, L.R. The strong perennial vision: A response. Agroecol. Sust. Food. 2015, 39, 500–515. [Google Scholar] [CrossRef]
- DeHaan, L.R.; Ismail, B.P. Perennial cereals provide ecosystem benefits. Cereal Foods World 2017, 62, 278–281. [Google Scholar] [CrossRef]
- de Oliveira, G.; Brunsell, N.A.; Sutherlin, C.E.; Crews, T.E.; DeHaan, L.R. Energy, water and carbon exchange over a perennial Kernza wheatgrass crop. Agric. Forest Meteorol. 2018, 249, 120–137. [Google Scholar] [CrossRef]
- Jarvis, P.G.; McNaughton, K.G. Stomatal control of transpiration: Scaling up from leaf to region. In Advances in Ecological Research; Macfadyen, A., Ford, E.D., Eds.; Academic Press: London, UK, 1986; Volume 15, pp. 1–49. ISBN 0-12-013915-4. [Google Scholar]
- Steduto, P.; Hsiao, T.C. Maize canopies under two soil water regimes: III. Variation in coupling with the atmosphere and the role of leaf area index. Agric. Forest Meteorol. 1998, 89, 201–213. [Google Scholar] [CrossRef]
- Oguntunde, P.G. Whole-plant water use and canopy conductance of cassava under limited available soil water and varying evaporative demand. Plant Soil 2005, 278, 371–383. [Google Scholar] [CrossRef]
- Marin, F.R.; Angelocci, L.R.; Nassif, D.S.; Costa, L.G.; Vianna, M.S.; Carvalho, K.S. Crop coefficient changes with reference evapotranspiration for highly canopy-atmosphere coupled crops. Agric. Water Manag. 2016, 163, 139–145. [Google Scholar] [CrossRef]
- Spinelli, G.M.; Snyder, R.L.; Sanden, B.L.; Gilbert, M.; Shackel, K.A. Low and variable atmospheric coupling in irrigated Almond (Prunus dulcis) canopies indicates a limited influence of stomata on orchard evapotranspiration. Agric. Water Manag. 2018, 196, 57–65. [Google Scholar] [CrossRef]
- Brunsell, N.A.; Schymanski, S.J.; Kleidon, A. Quantifying the thermodynamic entropy budget of the land surface: Is this useful? Earth Syst. Dynam. 2011, 2, 87–103. [Google Scholar] [CrossRef]
- Hammerle, A.; Haslwanter, A.; Schmitt, M.; Bahn, M.; Tappeiner, U.; Cernusca, A.; Wohlfahrt, G. Eddy covariance measurements of carbon dioxide, latent and sensible energy fluxes above a meadow on a mountain slope. Bound.-Lay. Meteorol. 2007, 122, 397–416. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Change Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Boden, T.A.; Krassovski, M.; Yang, B. The AmeriFlux data activity and data system: An evolving collection of data management techniques, tools, products and services. Geosci. Instrum. Meth. 2013, 2, 165–176. [Google Scholar] [CrossRef]
- Ganser, C.; Wisely, S.M. Patterns of spatio-temporal distribution, abundance, and diversity in a mosquito community from the eastern Smoky Hills of Kansas. J. Vector Ecol. 2013, 38, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Brunsell, N.A.; Nippert, J.B.; Buck, T.L. Impacts of seasonality and surface heterogeneity on water-use efficiency in mesic grasslands. Ecohydrology 2014, 7, 1223–1233. [Google Scholar] [CrossRef]
- Raz-Yaseef, N.; Billesbach, D.P.; Fischer, M.L.; Biraud, S.C.; Gunter, S.A.; Bradford, J.A.; Torn, M.S. Vulnerability of crops and native grasses to summer drying in the US Southern Great Plains. Agric. Ecosyst. Environ. 2015, 213, 209–218. [Google Scholar] [CrossRef]
- Cook, B.I.; Seager, R.; Smerdon, J.E. The worst North American drought year of the last millennium: 1934. Geophys. Res. Lett. 2014, 41, 7298–7305. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.; Keenan, T.F.; Fisher, J.B.; Baldocchi, D.D.; Desai, A.R.; Richardson, A.D.; Scott, R.L.; Law, B.E.; Litvak, M.E.; Brunsell, N.A.; et al. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc. Natl. Acad. Sci. USA 2016, 113, 5880–5885. [Google Scholar] [CrossRef] [Green Version]
- Crews, T.E.; Blesh, J.; Culman, S.W.; Hayes, R.C.; Jensen, E.S.; Mack, M.C.; Peoples, M.B.; Schipanski, M.E. Going where no grains have gone before: From early to mid-succession. Agric. Ecosyst. Environ. 2016, 223, 223–238. [Google Scholar] [CrossRef]
- Kelemen, A.; Tothmeresz, B.; Valko, O.; Miglecz, T.; Deak, B.; Torok, P. New aspects of grassland recovery in old-fields revealed by trait-based analyses of perennial-crop-mediated succession. Ecol. Evol. 2017, 7, 2432–2440. [Google Scholar] [CrossRef]
- Khatun, R.; Ohta, T.; Kotani, A.; Asanuma, J.; Gamo, M.; Han, S.; Hirano, T.; Nakai, Y.; Saigusa, N.; Takagi, K.; et al. Spatial variations in evapotranspiration over East Asian forest sites. I. Evapotranspiration and decoupling coefficient. Hydrol. Res. Lett. 2011, 5, 83–87. [Google Scholar] [CrossRef]
- Culman, S.W.; Snapp, S.S.; Ollenburger, M.; Basso, B.; DeHaan, L.R. Soil and water quality rapidly responds to the perennial grain Kernza wheatgrass. Agron. J. 2013, 105, 735–744. [Google Scholar] [CrossRef]
- Abraha, M.; Chen, J.; Chu, H.; Zenone, T.; John, R.; Su, Y.J.; Hamilton, S.K.; Robertson, G.P. Evapotranspiration of annual and perennial biofuel crops in a variable climate. Gcb. Bioenergy 2015, 7, 1344–1356. [Google Scholar] [CrossRef]
- Ferchaud, F.; Vitte, G.; Bornet, F.; Strullu, L.; Mary, B. Soil water uptake and root distribution of different perennial and annual bioenergy crops. Plant Soil 2015, 388, 307–322. [Google Scholar] [CrossRef]
- Georgescu, M.; Lobell, D.B.; Field, C.B. Direct climate effects of perennial bioenergy crops in the United States. Proc. Natl. Acad. Sci. USA 2011, 108, 4307–4312. [Google Scholar] [CrossRef] [Green Version]
- Eichelmann, E.; Wagner-Riddle, C.; Warland, J.; Deen, B.; Voroney, P. Evapotranspiration, water use efficiency, and energy partitioning of a mature switchgrass stand. Agric. Forest Meteorol. 2016, 217, 108–119. [Google Scholar] [CrossRef]
- Sindhoj, E.; Andrén, O.; Katterer, T.; Marissink, M.; Pettersson, R. Root biomass dynamics in a semi-natural grassland exposed to elevated atmospheric CO2 for five years. Acta Agric. Scand. B-S. P. 2004, 54, 50–59. [Google Scholar]
- Hainaut, P.; Remacle, T.; Decamps, C.; Lambert, R.; Sadok, W. Higher forage yields under temperate drought explained by lower transpiration rates under increasing evaporative demand. Eur. J. Agron. 2016, 72, 91–98. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Coefficient Estimates | Pr (>|t|) |
|---|---|---|
| LE | 4.827 × 10−4 | <2 × 10−16 |
| VPD | −4.210 × 10−2 | <2 × 10−16 |
| SWC | 5.688 × 10−2 | <2 × 10−16 |
| Rn | −1.248 × 10−5 | 0.00257 |
| Tair | −3.306 × 10−3 | <2 × 10−16 |
| Coupling Mechanisms | p-values | Multiple R2 | Adjusted R2 |
|---|---|---|---|
| Water Fluxes | <2 × 10−16 | 0.4556 | 0.4555 |
| Radiative Fluxes | <2 × 10−16 | 0.02753 | 0.02747 |
| Site | US-KON | US-KLS | US-KFS | US-ARM |
|---|---|---|---|---|
| LE Coefficient | 4.897 × 10−4 | 6.108 × 10−4 | 5.938 × 10−4 | 5.030 × 10−4 |
| VPD Coefficient | −6.716 × 10−2 | −4.897 × 10−2 | −6.757 × 10−2 | −2.325 × 10−2 |
| SWC Coefficient | 6.247 × 10−2 | −1.326 × 10−1 | −1.020 × 10−1 | 1.032 × 10−2 |
| Multiple R2 | 0.3772 | 0.5287 | 0.3527 | 0.5436 |
| Adjusted R2 | 0.3769 | 0.5285 | 0.3525 | 0.5434 |
| Site | US-KON | US-KLS | US-KFS | US-ARM |
|---|---|---|---|---|
| LE Pr (>|t|) | <2 × 10−16 | <2 × 10−16 | <2 × 10−16 | <2 × 10−16 |
| VPD Pr (>|t|) | <2 × 10−16 | <2 × 10−16 | <2 × 10−16 | <2 × 10−16 |
| SWC Pr (>|t|) | 1.99 × 10−5 | <2 × 10−16 | <2 × 10−16 | 0.134 |
| Overall p-value | <2.2 × 10−16 | <2.2 × 10−16 | <2.2 × 10−16 | <2.2 × 10−16 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutherlin, C.E.; Brunsell, N.A.; de Oliveira, G.; Crews, T.E.; R. DeHaan, L.; Vico, G. Contrasting Physiological and Environmental Controls of Evapotranspiration over Kernza Perennial Crop, Annual Crops, and C4 and Mixed C3/C4 Grasslands. Sustainability 2019, 11, 1640. https://doi.org/10.3390/su11061640
Sutherlin CE, Brunsell NA, de Oliveira G, Crews TE, R. DeHaan L, Vico G. Contrasting Physiological and Environmental Controls of Evapotranspiration over Kernza Perennial Crop, Annual Crops, and C4 and Mixed C3/C4 Grasslands. Sustainability. 2019; 11(6):1640. https://doi.org/10.3390/su11061640
Chicago/Turabian StyleSutherlin, Caitlyn E., Nathaniel A. Brunsell, Gabriel de Oliveira, Timothy E. Crews, Lee R. DeHaan, and Giulia Vico. 2019. "Contrasting Physiological and Environmental Controls of Evapotranspiration over Kernza Perennial Crop, Annual Crops, and C4 and Mixed C3/C4 Grasslands" Sustainability 11, no. 6: 1640. https://doi.org/10.3390/su11061640