Integrating LiDAR, Multispectral and SAR Data to Estimate and Map Canopy Height in Tropical Forests

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
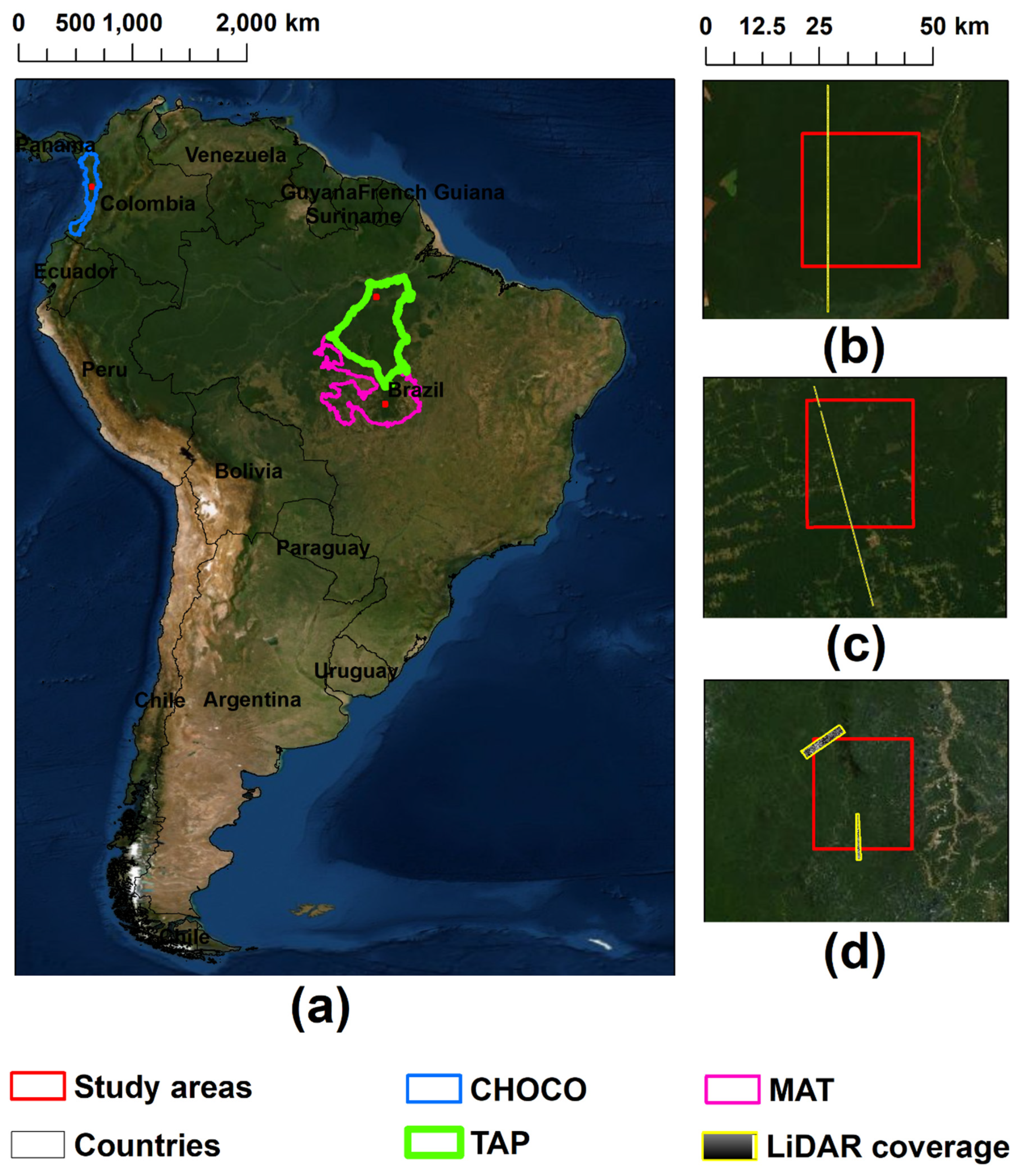
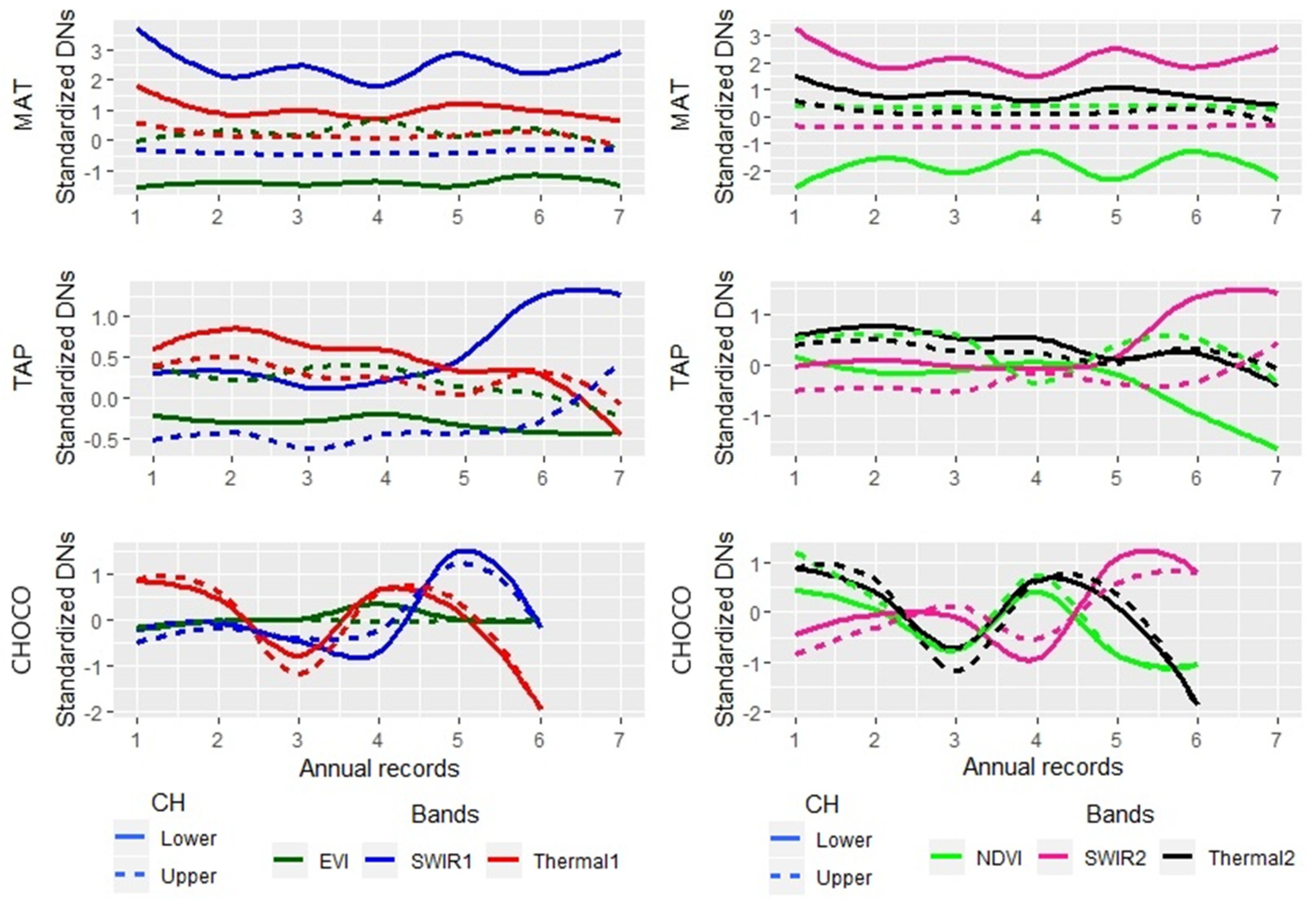
2.1. Study Areas and Forest Types
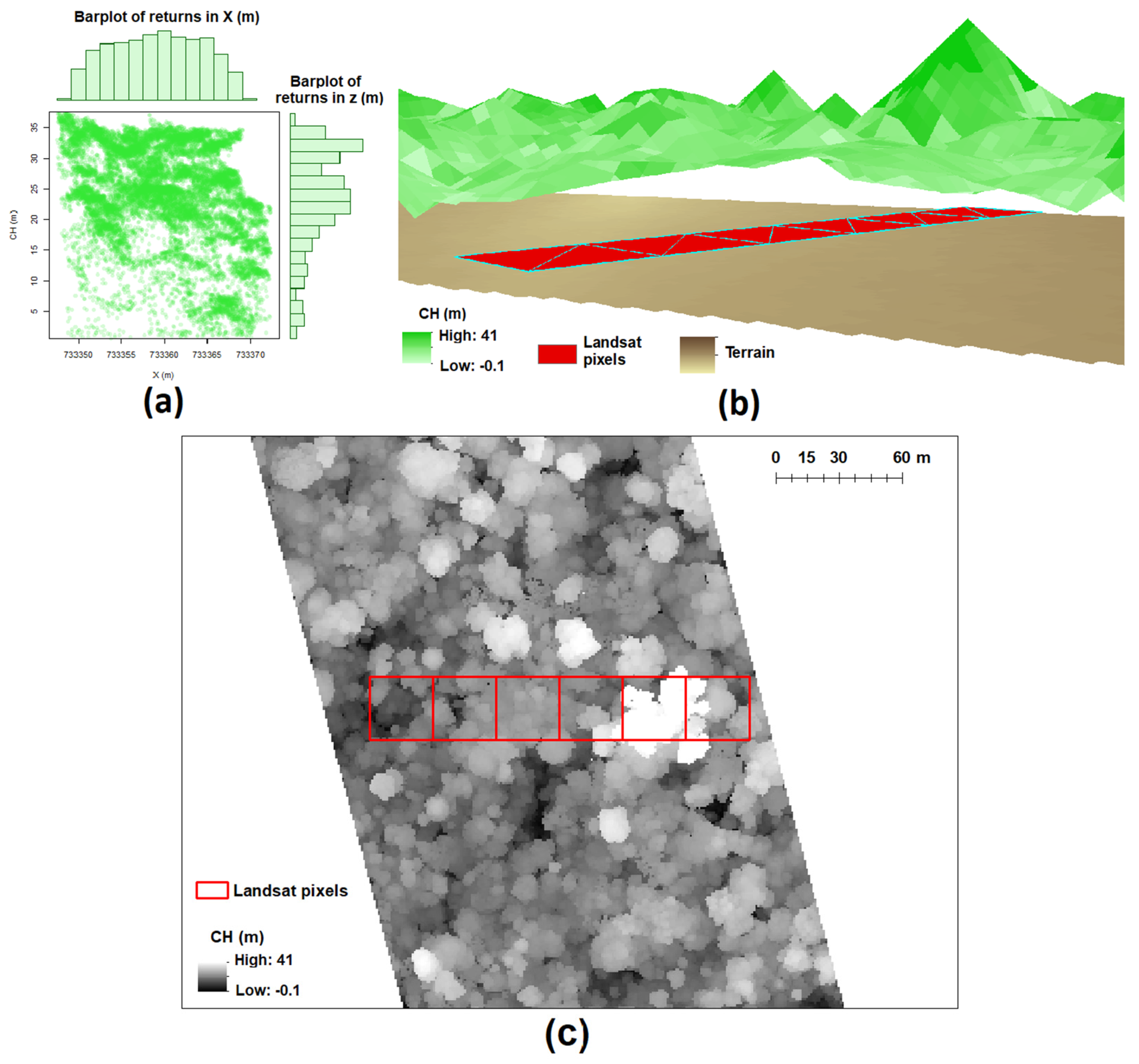
2.2. LiDAR Data and Canopy Height
2.3. Multispectral and SAR Data
2.4. Modeling and Mapping
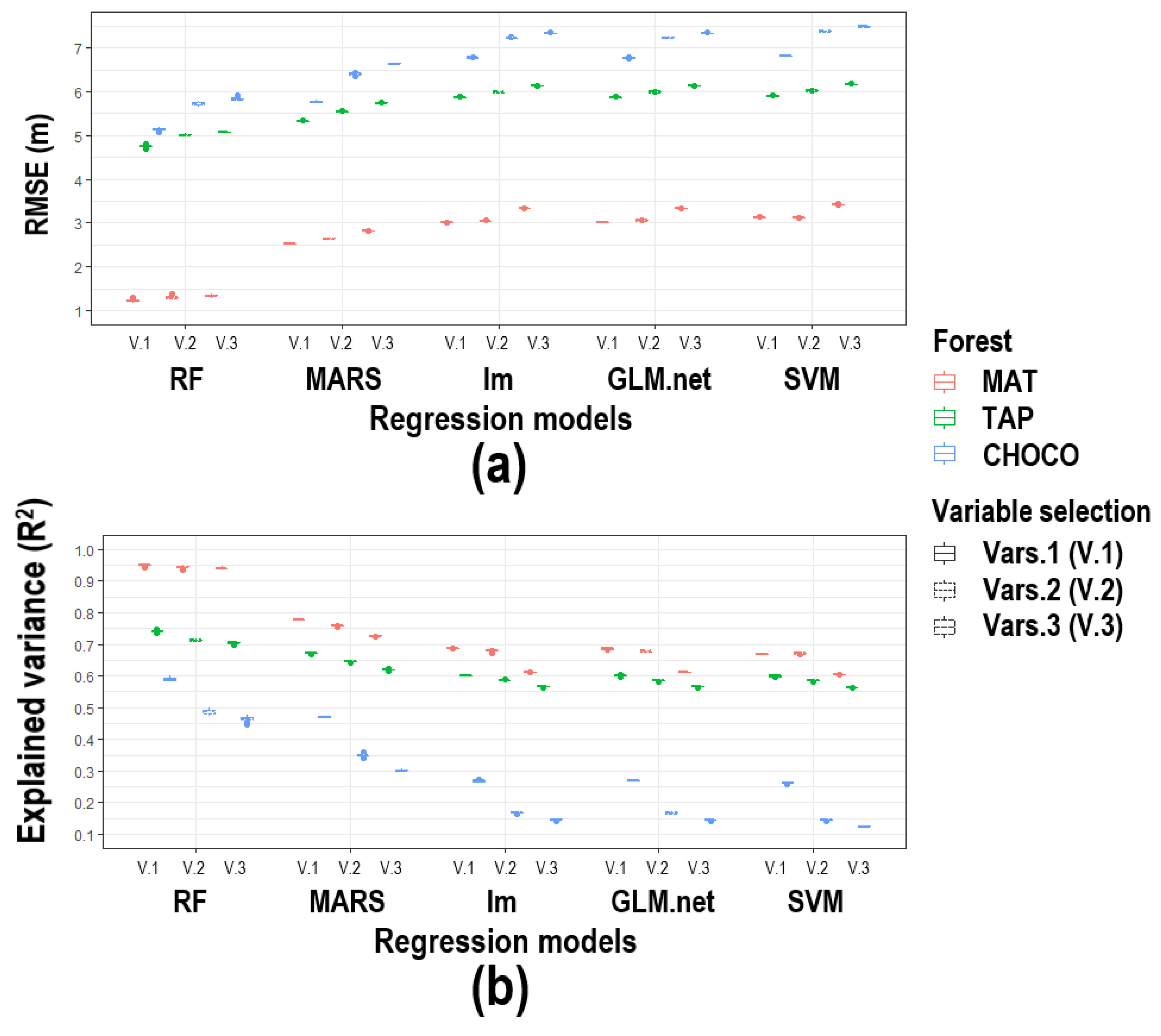
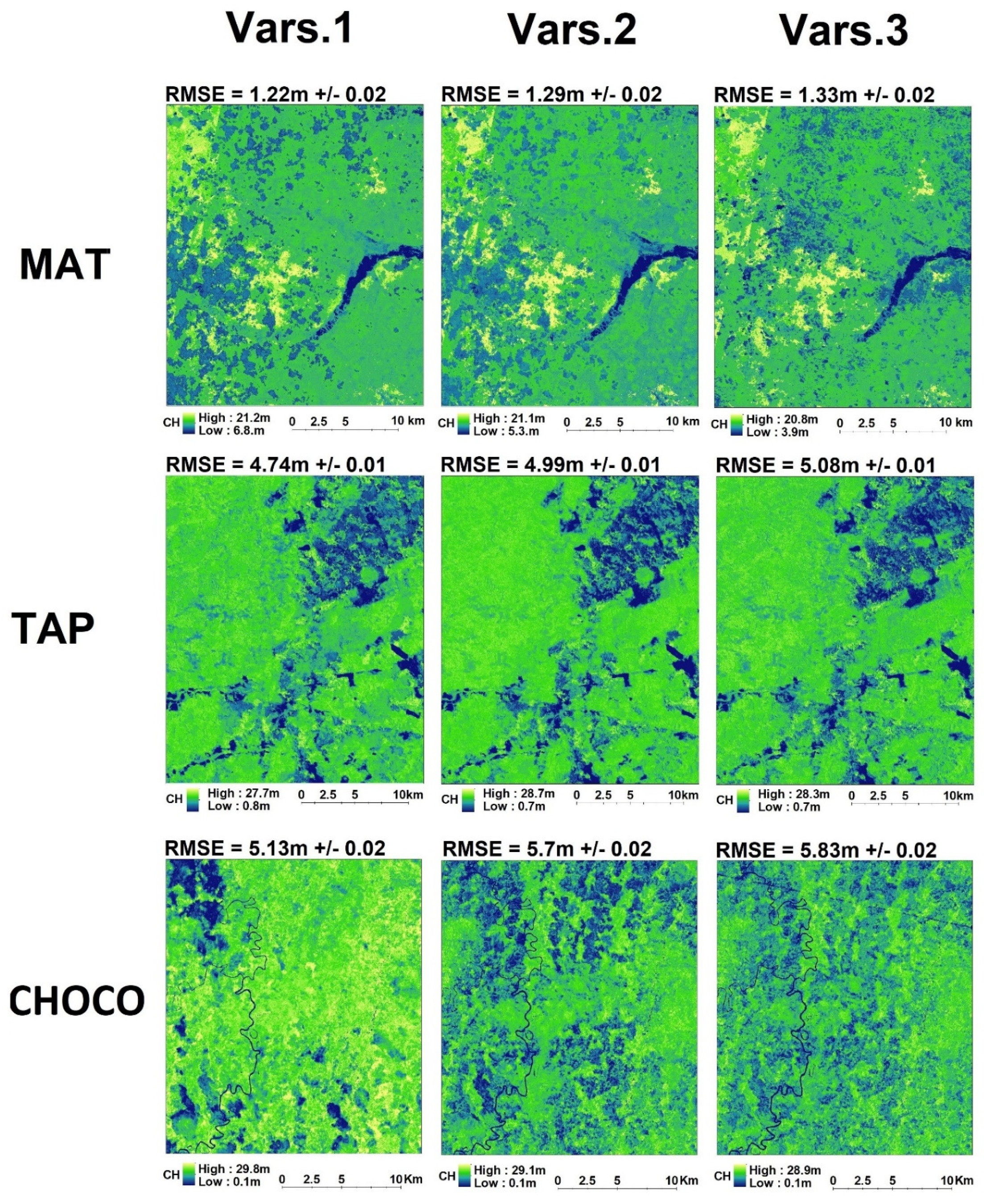
3. Results
4. Discussion
4.1. Improving CH Mapping
4.2. Map Accuracy Among Forest Types
4.3. Map Accuracy Among Predictor Sets
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pereira, H.M.; Ferrier, S.; Walters, M.; Geller, G.N.; Jongman, R.H.G.; Scholes, R.J.; Bruford, M.W.; Brummitt, N.; Butchart, S.H.M.; Cardoso, A.C.; et al. Essential biodiversity variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Skidmore, A.K.; Pettorelli, N.; Coops, N.C.; Geller, G.N.; Hansen, M.; Lucas, R.; Muecher, C.A.; O’Connor, B.; Paganini, M.; Pereira, H.M.; et al. Agree on biodiversity metrics to track from space. Nature 2015, 523, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Vihervaara, P.; Auvinen, A.-P.; Mononen, L.; Törmä, M.; Ahlroth, P.; Anttila, S.; Böttcher, K.; Forsius, M.; Heino, J.; Heliölä, J.; et al. How essential biodiversity variables and remote sensing can help national biodiversity monitoring. Glob. Ecol. Conserv. 2017, 10, 43–59. [Google Scholar] [CrossRef]
- Primack, R.B.; Corlett, R.T. Tropical Rain Forests: An Ecological and Biogeographical Comparison; Blackwell Publishing: Oxford, UK, 2009; ISBN 978-1-4051-4109-3. [Google Scholar]
- Hubbell, S.P.; Foster, R.B.; O’Brien, S.T.; Harms, K.E.; Condit, R.; Wechsler, B.; Wright, S.J.; de Lao, S.L. Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science 1999, 283, 554–557. [Google Scholar] [CrossRef]
- Kellner, J.R.; Clark, D.B.; Hubbell, S.P. Pervasive canopy dynamics produce short-term stability in a tropical rain forest landscape. Ecol. Lett. 2009, 12, 155–164. [Google Scholar] [CrossRef]
- Clark, D.B.; Olivas, P.C.; Oberbauer, S.F.; Clark, D.A.; Ryan, M.G. First direct landscape-scale measurement of tropical rain forest Leaf Area Index, a key driver of global primary productivity. Ecol. Lett. 2008, 11, 163–172. [Google Scholar] [CrossRef]
- Smith, M.N.; Stark, S.C.; Taylor, T.C.; Ferreira, M.L.; de Oliveira, E.; Restrepo-Coupe, N.; Chen, S.; Woodcock, T.; dos Santos, D.B.; Alves, L.F.; et al. Seasonal and drought-related changes in leaf area profiles depend on height and light environment in an Amazon forest. NEW Phytol. 2019, 222, 1284–1297. [Google Scholar] [CrossRef]
- LaFrankie, J.V.; Ashton, P.S.; Chuyong, G.B.; Co, L.; Condit, R.; Davies, S.J.; Foster, R.; Hubbell, S.P.; Kenfack, D.; Lagunzad, D.; et al. Contrasting structure and composition of the understory in species-rich tropical rain forests. Ecology 2006, 87, 2298–2305. [Google Scholar] [CrossRef]
- Tang, H.; Dubayah, R. Light-driven growth in Amazon evergreen forests explained by seasonal variations of vertical canopy structure. Proc. Natl. Acad. Sci. USA 2017, 114, 2640–2644. [Google Scholar] [CrossRef]
- Wu, J.; Albert, L.P.; Lopes, A.P.; Restrepo-Coupe, N.; Hayek, M.; Wiedemann, K.T.; Guan, K.; Stark, S.C.; Christoffersen, B.; Prohaska, N.; et al. Leaf development and demography explain photosynthetic seasonality in Amazon evergreen forests. Science 2016, 351, 972–976. [Google Scholar] [CrossRef]
- De Frenne, P.; Zellweger, F.; Rodriguez-Sanchez, F.; Scheffers, B.R.; Hylander, K.; Luoto, M.; Vellend, M.; Verheyen, K.; Lenoir, J. Global buffering of temperatures under forest canopies. Nat. Ecol. Evol. 2019, 3, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, U.; Nino, S.; Barrientoso, L.; Trevino, J.; Almaguer, P. Seasonal microclimatic variation in a succession gradient of low thorn forest in Northeastern Mexico. Rev. Biol. Trop. 2019, 67, 266–277. [Google Scholar]
- Jucker, T.; Hardwick, S.R.; Both, S.; Elias, D.M.O.; Ewers, R.M.; Milodowski, D.T.; Swinfield, T.; Coomes, D.A. Canopy structure and topography jointly constrain the microclimate of human-modified tropical landscapes. Glob. Chang. Biol. 2018, 24, 5243–5258. [Google Scholar] [CrossRef]
- Hadi; Pfeifer, M.; Korhonen, L.; Wheeler, C.; Rautiainen, M. Forest canopy structure and reflectance in humid tropical Borneo: A physically-based interpretation using spectral invariants. Remote Sens. Environ. 2017, 201, 314–330. [Google Scholar] [CrossRef]
- Drake, J.B.; Knox, R.G.; Dubayah, R.O.; Clark, D.B.; Condit, R.; Blair, J.B.; Hofton, M. Above-ground biomass estimation in closed canopy Neotropical forests using lidar remote sensing: Factors affecting the generality of relationships. Glob. Ecol. Biogeogr. 2003, 12, 147–159. [Google Scholar] [CrossRef]
- Goetz, S.; Dubayah, R. Advances in remote sensing technology and implications for measuring and monitoring forest carbon stocks and change. CARBON Manag. 2011, 2, 231–244. [Google Scholar] [CrossRef]
- Guan, H.; Li, J.; Yu, Y.; Zhong, L.; Ji, Z. DEM generation from lidar data in wooded mountain areas by cross-section-plane analysis. Int. J. Remote Sens. 2014, 35, 927–948. [Google Scholar] [CrossRef]
- McRoberts, R.E.; Chen, Q.; Gormanson, D.D.; Walters, B.F. The shelf-life of airborne laser scanning data for enhancing forest inventory inferences. Remote Sens. Environ. 2018, 206, 254–259. [Google Scholar] [CrossRef]
- Huang, W.; Dolan, K.; Swatantrad, A.; Johnson, K.; Tang, H.; O’Neil-Dunne, J.; Dubayah, R.; Hurtt, G. High-resolution mapping of aboveground biomass for forest carbon monitoring system in the Tri-State region of Maryland, Pennsylvania and Delaware, USA. Environ. Res. Lett. 2019, 14, 1–16. [Google Scholar] [CrossRef]
- Silva, C.A.; Saatchi, S.; Garcia, M.; Labriere, N.; Klauberg, C.; Ferraz, A.; Meyer, V.; Jeffery, K.J.; Abernethy, K.; White, L.; et al. Comparison of small-and large-footprint lidar characterization of tropical forest aboveground structure and biomass: A case study from central gabon. IEEE J. Sel. Top. Appl. EARTH Obs. Remote Sens. 2018, 11, 3512–3526. [Google Scholar] [CrossRef]
- Asner, G.P.; Powell, G.V.N.; Mascaro, J.; Knapp, D.E.; Clark, J.K.; Jacobson, J.; Kennedy-Bowdoin, T.; Balaji, A.; Paez-Acosta, G.; Victoria, E.; et al. High-resolution forest carbon stocks and emissions in the Amazon. Proc. Natl. Acad. Sci. USA 2010, 107, 16738–16742. [Google Scholar] [CrossRef] [PubMed]
- Dubayah, R.O.; Sheldon, S.L.; Clark, D.B.; Hofton, M.A.; Blair, J.B.; Hurtt, G.C.; Chazdon, R.L. Estimation of tropical forest height and biomass dynamics using lidar remote sensing at La Selva, Costa Rica. J. Geophys. Res. 2010, 115, 1–17. [Google Scholar] [CrossRef]
- Tang, H.; Song, X.-P.; Zhao, F.A.; Strahler, A.H.; Schaaf, C.L.; Goetz, S.; Huang, C.; Hansen, M.C.; Dubayah, R. Definition and measurement of tree cover: A comparative analysis of field-, lidar-and landsat-based tree cover estimations in the Sierra national forests, USA. Agric. For. Meteorol. 2019, 268, 258–268. [Google Scholar] [CrossRef]
- Blair, J.B.; Hofton, M.A. Modeling laser altimeter return waveforms over complex vegetation using high-resolution elevation data. Geophys. Res. Lett. 1999, 26, 2509–2512. [Google Scholar] [CrossRef]
- Hancock, S.; Armston, J.; Hofton, M.; Sun, X.; Tang, H.; Duncanson, L.I.; Kellner, J.R.; Dubayah, R. The GEDI Simulator: A Large-footprint waveform lidar simulator for calibration and validation of spaceborne missions. EARTH Space Sci. 2019, 6, 294–310. [Google Scholar] [CrossRef] [PubMed]
- Popescu, S.C.; Zhao, K.; Neuenschwander, A.; Lin, C. Satellite lidar vs. small footprint airborne lidar: Comparing the accuracy of aboveground biomass estimates and forest structure metrics at footprint level. Remote Sens. Environ. 2011, 115, 2786–2797. [Google Scholar] [CrossRef]
- Bi, J.; Knyazikhin, Y.; Choi, S.; Park, T.; Barichivich, J.; Ciais, P.; Fu, R.; Ganguly, S.; Hall, F.; Hilker, T.; et al. Sunlight mediated seasonality in canopy structure and photosynthetic activity of Amazonian rainforests. Environ. Res. Lett. 2015, 10, 064014. [Google Scholar] [CrossRef]
- Doughty, C.E.; Goulden, M.L. Seasonal patterns of tropical forest leaf area index and CO2 exchange. J. Geophys. Res. 2008, 113, 1–12. [Google Scholar] [CrossRef]
- Rappaport, D.I.; Morton, D.C.; Longo, M.; Keller, M.; Dubayah, R.; dos-Santos, M.N. Quantifying long-term changes in carbon stocks and forest structure from Amazon forest degradation. Environ. Res. Lett. 2018, 13, 065013. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Brando, P.M.; Goetz, S.J.; Baccini, A.; Nepstad, D.C.; Beck, P.S.A.; Christman, M.C. Seasonal and interannual variability of climate and vegetation indices across the Amazon. Proc. Natl. Acad. Sci. USA 2010, 107, 14685–14690. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Armston, J.; Hancock, S.; Marselis, S.; Goetz, S.; Dubayah, R. Characterizing global forest canopy cover distribution using spaceborne lidar. Remote Sens. Environ. 2019, 231, 111262. [Google Scholar] [CrossRef]
- Brown, L.; Chen, J.M.; Leblanc, S.G.; Cihlar, J. A shortwave infrared modification to the simple ratio for LAI retrieval in boreal forests: An image and model analysis. Remote Sens. Environ. 2000, 71, 16–25. [Google Scholar] [CrossRef]
- Meyer, V.; Saatchi, S.; Ferraz, A.; Xu, L.; Duque, A.; Garcia, M.; Chave, J. Forest degradation and biomass loss along the Choco region of Colombia. Carbon Balance Manag. 2019, 14, 2. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Goetz, S.J.; Turubanova, S.; Tyukavina, A.; Krylov, A.; Kommareddy, A.; Egorov, A. Mapping tree height distributions in Sub-Saharan Africa using Landsat 7 and 8 data. Remote Sens. Environ. 2016, 185, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Cartus, O.; Santoro, M.; Kellndorfer, J. Mapping forest aboveground biomass in the Northeastern United States with ALOS PALSAR dual-polarization L-band. Remote Sens. Environ. 2012, 124, 466–478. [Google Scholar] [CrossRef]
- Berninger, A.; Lohberger, S.; Staengel, M.; Siegert, F. SAR-based estimation of above-ground biomass and its changes in tropical forests of kalimantan using L- and C-Band. Remote Sens. 2018, 10, 831. [Google Scholar] [CrossRef] [Green Version]
- Saatchi, S.; Moghaddam, M. Estimation of crown and stem water content and biomass of boreal forest using polarimetric SAR imagery. IEEE Trans. Geosci. Remote Sens. 2000, 38, 697–709. [Google Scholar] [CrossRef] [Green Version]
- Shimada, M.; Itoh, T.; Motooka, T.; Watanabe, M.; Shiraishi, T.; Thapa, R.; Lucas, R. New global forest/non-forest maps from ALOS PALSAR data (2007–2010). Remote Sens. Environ. 2014, 155, 13–31. [Google Scholar] [CrossRef]
- Saatchi, S.; Marlier, M.; Chazdon, R.L.; Clark, D.B.; Russell, A.E. Impact of spatial variability of tropical forest structure on radar estimation of aboveground biomass. Remote Sens. Environ. 2011, 115, 2836–2849. [Google Scholar] [CrossRef]
- Urbazaev, M.; Cremer, F.; Migliavacca, M.; Reichstein, M.; Schmullius, C.; Thiel, C. Potential of multi-temporal ALOS-2 PALSAR-2 ScanSAR data for vegetation height estimation in tropical forests of Mexico. Remote Sens. 2018, 10, 1277. [Google Scholar] [CrossRef] [Green Version]
- Luckman, A.; Baker, J.; Honzák, M.; Lucas, R. Tropical forest biomass density estimation using JERS-1 SAR: Seasonal variation, confidence limits, and application to image mosaics. Remote Sens. Environ. 1998, 63, 126–139. [Google Scholar] [CrossRef]
- Saatchi, S.; Harris, N.L.; Brown, S.; Lefsky, M.; Mitchard, E.T.A.; Salas, W.; Zutta, B.R.; Buermann, W.; Lewis, S.L.; Hagen, S.; et al. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl. Acad. Sci. USA 2011, 108, 9899–9904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kugler, F.; Schulze, D.; Hajnsek, I.; Pretzsch, H.; Papathanassiou, K.P. TanDEM-X Pol-InSAR performance for forest height estimation. IEEE Trans. Geosci. Remote Sens. 2014, 52, 6404–6422. [Google Scholar] [CrossRef]
- Qi, W.; Dubayah, R.O. Combining Tandem-X InSAR and simulated GEDI lidar observations for forest structure mapping. Remote Sens. Environ. 2016, 187, 253–266. [Google Scholar] [CrossRef]
- Qi, W.; Lee, S.-K.; Hancock, S.; Luthcke, S.; Tang, H.; Armston, J.; Dubayah, R. Improved forest height estimation by fusion of simulated GEDI Lidar data and TanDEM-X InSAR data. Remote Sens. Environ. 2019, 221, 621–634. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Levick, S.R.; Heidrich, L.; Magdon, P.; Leutner, B.F.; Woellauer, S.; Serebryanyk, A.; Nauss, T.; Krzystek, P.; Gossner, M.M.; et al. Radar vision in the mapping of forest biodiversity from space. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Cartus, O.; Santoro, M.; Wegmuller, U.; Rommen, B. Benchmarking the retrieval of biomass in boreal forests using P-Band SAR backscatter with multi-temporal C- and L-Band observations. Remote Sens. 2019, 11, 1695. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.D.; Yoshino, K.; Le, N.N.; Bui, D.T. Estimating aboveground biomass of a mangrove plantation on the Northern coast of Vietnam using machine learning techniques with an integration of ALOS-2 PALSAR-2 and Sentinel-2A data. Int. J. Remote Sens. 2018, 39, 7761–7788. [Google Scholar] [CrossRef]
- Abernethy, K.; Bush, E.R.; Forget, P.-M.; Mendoza, I.; Morellato, L.P.C. Current issues in tropical phenology: A synthesis. Biotropica 2018, 50, 477–482. [Google Scholar] [CrossRef] [Green Version]
- WWF. Mato Grosso Dry Forest. Available online: https://www.worldwildlife.org/ecoregions/nt0140 (accessed on 1 August 2018).
- Vourlitis, G.L.; Priante, N.; Hayashi, M.M.S.; Nogueira, J.D.; Caseiro, F.T.; Campelo, J.H. Seasonal variations in the evapotranspiration of a transitional tropical forest of Mato Grosso, Brazil. WATER Resour. Res. 2002, 38, 30-1–30-11. [Google Scholar] [CrossRef]
- WWF. Tapajós-Xingu Moist Forest. Available online: https://www.worldwildlife.org/ecoregions/nt0168 (accessed on 1 August 2018).
- WWF. Choco-Darien Moist Forests. Available online: http://wwf.panda.org/about_our_earth/ecoregions/chocodarien_moist_forests.cfm (accessed on 1 August 2018).
- Gentry, A.H. Species richness and floristic composition of Choco Region plant communities. Caldasia 1986, 15, 5. [Google Scholar]
- Gregory-Wodzicki, K.M. Uplift history of the central and northern Andes: A review. Geol. Soc. Am. Bull. 2000, 7, 14. [Google Scholar] [CrossRef]
- Poveda, G.; Mesa, O.J. On the existence of Lloro (the rainiest locality on earth): Enhanced ocean-land-atmosphere interaction by a low-level jet. Geophys. Res. Lett. 2000, 27, 1675–1678. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.L.; Roberts, D.A.; Ewel, J.J.; Clark, D.B. Estimation of tropical rain forest aboveground biomass with small-footprint lidar and hyperspectral sensors. Remote Sens. Environ. 2011, 115, 2931–2942. [Google Scholar] [CrossRef]
- Hijmans, R.; van Etten, J.; Cheng, J.; Mattiuzzi, M.; Sumner, M.; Greenberg, J.; Perpinan, O.; Bevan, A.; Racine, E.; Shortridge, A. Package ‘Raster’: Geographic Data Analysis and Modeling. Package ‘Raster’. 2016. Available online: https://www.r-project.org/ (accessed on 1 August 2018).
- Roussel, J.-R.; David, A.; De Boissieu, F.; Meador, A.S. Package ‘lidR’: Airborne LiDAR Data Manipulation and Visualization for Forestry Applications. Package ‘lidR’. 2019. Available online: https://www.r-project.org/ (accessed on 1 August 2018).
- Huete, A.R.; Justice, C.; Leeuwen, W. MODIS Vegetation Index, Algorithm Theoretical Basis Document; University of Arizona and University of Virginia: Tucson, AZ, USA, 1999. [Google Scholar]
- Li, Z.; Li, X.; Wei, D.; Xu, X.; Wang, H. An assessment of correlation on MODIS-NDVI and EVI with natural vegetation coverage in Northern Hebei Province, China. Procedia Environ. Sci. 2010, 2, 964–969. [Google Scholar] [CrossRef] [Green Version]
- Fagua, J.C.; Ramsey, R.D. Geospatial modeling of land cover change in the Chocó-Darien global ecoregion of South America; one of most biodiverse and rainy areas in the world. PLoS ONE 2019, 14, e0211324. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google earth engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- USGS, U.G.S. USGS Landsat 8 Surface Reflectance Tier 1. Available online: https://developers.google.com/earth-engine/datasets/catalog/LANDSAT_LC08_C01_T1_SR (accessed on 1 August 2018).
- Anderson, M.C.; Allen, R.G.; Morse, A.; Kustas, W.P. Use of Landsat thermal imagery in monitoring evapotranspiration and managing water resources. Remote Sens. Environ. 2012, 122, 50–65. [Google Scholar] [CrossRef]
- ESA, E.S.A. Sentinel-1 SAR GRD: C-band Synthetic Aperture Radar Ground Range Detected, Log Scaling. Available online: https://developers.google.com/earth-engine/datasets/catalog/COPERNICUS_S1_GRD (accessed on 1 August 2018).
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carre, G.; Garcia Marquez, J.R.; Gruber, B.; Lafourcade, B.; Leitao, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Babak, N. Package ‘usdm’: Uncertainty Analysis for Species Distribution Models. Package ‘usdm’. 2015. Available online: https://www.r-project.org/ (accessed on 1 August 2018).
- Chai, T.; Draxler, R.R. Root mean square error (RMSE) or mean absolute error (MAE)?—Arguments against avoiding RMSE in the literature. Geosci. Model Dev. 2014, 7, 1247–1250. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, M. Package ‘caret’: Classification and Regression Training. Available online: https://github.com/topepo/caret/ (accessed on 1 August 2018).
- Ripley, B.; Venables, B.; Bates, D.; Hornik, K.; Gebhardt, A.; Firth, D. Package ‘MASS’; Springer: New York, NY, USA, 2019; ISBN 0-387-95457-0. [Google Scholar]
- Liaw, A. Package ‘randomForest’: Breiman and Cutler’s Random Forests for Classification and Regression; Package ‘Raster’; 2018. Available online: https://www.r-project.org/ (accessed on 1 August 2018).
- Meyer, D.; Dimitriadou, E.; Hornik, K.; Weingessel, A.; Leisch, F.; Chang, C.-C.; Lin, C.-C. Package ‘e1071’. 2017. Available online: https://www.r-project.org/ (accessed on 1 August 2018).
- Milborrow, S.; Tibshirani, R. Package ‘earth’: Multivariate Adaptive Regression Splines. Package ‘earth’. 2019. Available online: https://www.r-project.org/ (accessed on 1 August 2018).
- Friedman, J.; Hastie, T.; Tibshirani, R.; Simon, N.; Narasimhan, B.; Qian, J. Package ‘glmnet’: Lasso and Elastic-Net Regularized Generalized Linear Models. Package ‘glmnet’. 2019. Available online: https://www.r-project.org/ (accessed on 1 August 2018).
- Friedman, J.H. Multivariate adaptive regression splines. Ann. Stat. 1991, 19, 1–67. [Google Scholar] [CrossRef]
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization paths for generalized linear models via coordinate descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potapov, P.; Tyukavina, A.; Turubanova, S.; Talero, Y.; Hernandez-Serna, A.; Hansen, M.C.; Saah, D.; Tenneson, K.; Poortinga, A.; Aekakkararungroj, A.; et al. Annual continuous fields of woody vegetation structure in the Lower Mekong region from 2000–2017 Landsat time-series. Remote Sens. Environ. 2019, 232, 111278. [Google Scholar] [CrossRef]
- Fagua, J.C.; Ramsey, R.D. Comparing the accuracy of MODIS data products for vegetation detection between two environmentally dissimilar ecoregions: The chocó-darien of South America and the great basin of North America. GIScience Remote Sens. 2019, 56, 1–19. [Google Scholar] [CrossRef]
- Nitze, I.; Barrett, B.; Cawkwell, F. Temporal optimisation of image acquisition for land cover classification with Random Forest and MODIS time-series. Int. J. Appl. Earth Obs. Geoinf. 2015, 34, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, H.; Miura, T.; Dematte, J.A.M.; Batchily, K.; Huete, A.R. Soil line influences on two-band vegetation indices and vegetation isolines: A numerical study. Remote Sens. 2010, 2, 545–561. [Google Scholar] [CrossRef] [Green Version]
- Hardwick, S.R.; Toumi, R.; Pfeifer, M.; Turner, E.C.; Nilus, R.; Ewers, R.M. The relationship between leaf area index and microclimate in tropical forest and oil palm plantation: Forest disturbance drives changes in microclimate. Agric. For. Meteorol. 2015, 201, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Thi, D.N.; Ha, N.T.T.; Dang, Q.T.; Koike, K.; Trong, N.M. Effective Band ratio of landsat 8 images based on VNIR-SWIR reflectance spectra of topsoils for soil moisture mapping in a tropical region. Remote Sens. 2019, 11, 716. [Google Scholar]
- Asner, G.P. Biophysical and biochemical sources of variability in canopy reflectance. Remote Sens. Environ. 1998, 64, 234–253. [Google Scholar] [CrossRef]
- Bayat, B.; van der Tol, C.; Verhoef, W. Integrating satellite optical and thermal infrared observations for improving daily ecosystem functioning estimations during a drought episode. Remote Sens. Environ. 2018, 209, 375–394. [Google Scholar] [CrossRef]
- Liu, W.; Hong, Y.; Khan, S.I.; Huang, M.; Vieux, B.; Caliskan, S.; Grout, T. Actual evapotranspiration estimation for different land use and land cover in urban regions using Landsat 5 data. J. Appl. Remote Sens. 2010, 4, 041873. [Google Scholar]
- Lu, D.; Song, K.; Zang, S.; Jia, M.; Du, J.; Ren, C. The effect of urban expansion on urban surface temperature in Shenyang, China: An analysis with landsat imagery. Environ. Model. Assess. 2015, 20, 197–210. [Google Scholar] [CrossRef]
- Gomis-Cebolla, J.; Carlos Jimenez, J.; Antonio Sobrino, J.; Corbari, C.; Mancini, M. Intercomparison of remote-sensing based evapotranspiration algorithms over amazonian forests. Int. J. Appl. EARTH Obs. Geoinf. 2019, 80, 280–294. [Google Scholar] [CrossRef]
- Bartkowiak, P.; Castelli, M.; Notarnicola, C. Downscaling land surface temperature from MODIS dataset with random forest approach over alpine vegetated areas. Remote Sens. 2019, 11, 1319. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Im, J.; Jang, E.; Rhee, J. Drought assessment and monitoring through blending of multi-sensor indices using machine learning approaches for different climate regions. Agric. For. Meteorol. 2016, 216, 157–169. [Google Scholar] [CrossRef]
- Mermoz, S.; Le Toan, T.; Villard, L.; Réjou-Méchain, M.; Seifert-Granzin, J. Biomass assessment in the Cameroon savanna using ALOS PALSAR data. Remote Sens. Environ. 2014, 155, 109–119. [Google Scholar] [CrossRef]
- Liao, Z.; He, B.; Quan, X.; van Dijk, A.I.J.M.; Qiu, S.; Yin, C. Biomass estimation in dense tropical forest using multiple information from single-baseline P-band PolInSAR data. Remote Sens. Environ. 2019, 221, 489–507. [Google Scholar] [CrossRef]
- Grote, S.; Condit, R.; Hubbell, S.; Wirth, C.; Rueger, N. Response of demographic rates of tropical trees to light availability: Can position-based competition indices replace information from canopy census data? PLoS ONE 2013, 8, e81787. [Google Scholar] [CrossRef] [Green Version]
- Guan, K.; Pan, M.; Li, H.; Wolf, A.; Wu, J.; Medvigy, D.; Caylor, K.K.; Sheffield, J.; Wood, E.F.; Malhi, Y.; et al. Photosynthetic seasonality of global tropical forests constrained by hydroclimate. Nat. Geosci. 2015, 8, 284–289. [Google Scholar] [CrossRef]
- Moura, M.M.; dos Santos, A.R.; Pezzopane, J.E.M.; Alexandre, R.S.; da Silva, S.F.; Pimentel, S.M.; de Andrade, M.S.S.; Silva, F.G.R.; Branco, E.R.F.; Moreira, T.R.; et al. Relation of El Niño and La Niña phenomena to precipitation, evapotranspiration and temperature in the Amazon basin. Sci. Total Environ. 2019, 651, 1639–1651. [Google Scholar] [CrossRef]
- Álvarez-Dávila, E.; Cayuela, L.; González-Caro, S.; Aldana, A.M.; Stevenson, P.R.; Phillips, O.; Cogollo, Á.; Peñuela, M.C.; von Hildebrand, P.; Jiménez, E.; et al. Forest biomass density across large climate gradients in northern South America is related to water availability but not with temperature. PLoS ONE 2017, 12, e0171072. [Google Scholar] [CrossRef]
- Shi, L.; Westerhuis, J.A.; Rosen, J.; Landberg, R.; Brunius, C. Variable selection and validation in multivariate modelling. Bioinformatics 2019, 35, 972–980. [Google Scholar] [CrossRef]
- Kulkarni, A.; Shrestha, A. Multispectral image analysis using decision trees. Int. J. Adv. Comput. Sci. Appl. 2017, 8, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Liu, H.; Zhao, W.; Shi, T.; Hu, Z.; Li, Q.; Wu, G. Comparison of machine learning techniques in inferring phytoplankton size classes. Remote Sens. 2018, 10, 191. [Google Scholar] [CrossRef] [Green Version]
- Safari, A.; Sohrabi, H.; Powell, S.; Shataee, S. A comparative assessment of multi-temporal Landsat 8 and machine learning algorithms for estimating aboveground carbon stock in coppice oak forests. Int. J. Remote Sens. 2017, 38, 6407–6432. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Study Area (km2) | LiDAR Coverage (km2) | Number of Sites for the Regressions | Training Sites for Each 5-cross Validation Iteration | Validation Sites for Each 5-cross Validation Iteration |
|---|---|---|---|---|---|
| MAT | 734.1 | 10.0 | 7408 | ~5927 | ~1481 |
| TAP | 650.4 | 9.7 | 3207 | ~2566 | ~641 |
| CHOCO | 504.9 | 27.7 | 3024 | ~2420 | ~604 |
| MAT Seasonal Dry forest (2050 mm) | TAP Moist Forests (2147 mm) | CHO Super Moist Forest (8000 mm) | ||||
|---|---|---|---|---|---|---|
| P values | t values | P values | t values | P values | t values | |
| SWIR1 | 0.000 *** | −7.8 | 0.000 *** | −5.1 | 0.649 | −0.48 |
| SWIR2 | 0.000 *** | −7.3 | 0.04 * | −2.6 | 0.616 | −0.53 |
| Thermal1 | 0.001 ** | −6.3 | 0.01 * | −3.4 | 0.721 | −0.37 |
| Thermal2 | 0.001 ** | −5.8 | 0.04 * | −2.6 | 0.798 | −0.27 |
| EVI | 0.000 *** | 9.0 | 0.000 *** | 12.9 | 0.340 | 1.04 |
| NDVI | 0.000 *** | −13.5 | 0.000 *** | −17.7 | 0.011 * | −3.63 |
| L band; pol. HH | 0.05 * | 11.2 | 0.12 | 5.2 | 0.038 * | 16.63 |
| L band; pol. HV | 0.049 * | 12.8 | 0.03 * | 18.6 | 0.194 | 3.17 |
| C band; pol. VH | 0.000 *** | 6.2 | 0.000 *** | 6.1 | 0.000 *** | 6417 |
| C band; pol. VV | 0.01 * | 3.2 | 0.03 * | 2.5 | 0.36 | 0.97 |
| Mato Grosso Seasonal Dry Forest (2050 mm) | Tapajós-Xingu Moist Forest (2147 mm) | Chocó-Darien Moist Forest (8000 mm) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pearson | Kendall | Pearson | Kendall | Pearson | Kendall | |||||||
| r value | p value | tau value | p value | r value | p value | tau value | p value | r value | p value | tau value | p value | |
| SWIR 1 | −0.72 | *** | −0.23 | *** | −0.62 | *** | −0.39 | *** | −0.24 | *** | −0.15 | *** |
| SWIR 2 | −0.70 | *** | −0.24 | *** | −0.54 | *** | −0.35 | *** | −0.22 | *** | −0.14 | *** |
| Thermal 1 | −0.55 | *** | −0.21 | *** | −0.54 | *** | −0.34 | *** | −0.02 | 0.38 | 0.00 | 0.87 |
| Thermal 2 | −0.47 | *** | −0.19 | *** | −0.47 | *** | −0.30 | *** | −0.04 | 0.05 | −0.03 | 0.02 |
| EVI | 0.56 | *** | 0.16 | *** | 0.21 | *** | 0.11 | *** | 0.19 | *** | 0.02 | 0.04 |
| NDVI | 0.66 | *** | 0.17 | *** | 0.36 | *** | 0.21 | *** | 0.20 | *** | 0.12 | *** |
| L band; pol. HH | 0.26 | *** | 0.09 | *** | 0.40 | *** | 0.26 | *** | 0.10 | *** | 0.02 | 0.04 |
| L band; pol. HV | 0.41 | *** | 0.15 | *** | 0.55 | *** | 0.35 | *** | 0.21 | *** | 0.05 | *** |
| C band; pol. VH | 0.37 | *** | 0.01 | 0.08 | 0.12 | *** | 0.07 | *** | 0.20 | *** | 0.02 | 0.09 |
| C band; pol. VV | 0.38 | *** | 0.01 | 0.14 | 0.12 | *** | 0.07 | *** | 0.21 | *** | 0.02 | 0.07 |
| Mato Grosso Seasonal Dry Forest (2100 mm) | Tapajós-Xingu Moist Forests (1500–2000 mm) | Chocó-Darien Moist Forest (8000–13,000 mm) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pearson | Kendall | Pearson | Kendall | Pearson | Kendall | |||||||
| r Value | p Value | tau Value | p Value | r Value | p Value | tau Value | p Value | r Value | p Value | tau Value | p Value | |
| SWIR 1 | −0.15 | *** | −0.14 | *** | −0.05 | 0.002 | −0.05 | *** | −0.10 | *** | −0.06 | *** |
| SWIR 2 | −0.10 | *** | −0.09 | *** | −0.04 | 0.008 | −0.03 | 0.003 | −0.11 | *** | −0.07 | *** |
| Thermal 1 | −0.17 | *** | −0.18 | *** | −0.06 | *** | −0.01 | 0.34 | −0.15 | *** | −0.16 | *** |
| Thermal 2 | −0.13 | *** | −0.14 | *** | −0.04 | 0.009 | 0.00 | 0.95 | −0.13 | *** | −0.10 | *** |
| EVI | −0.56 | *** | −0.37 | *** | −0.06 | *** | −0.05 | *** | 0.01 | 0.65 | −0.03 | 0.01 |
| NDVI | −0.29 | *** | −0.19 | *** | 0.11 | *** | 0.05 | *** | −0.06 | 0.002 | −0.03 | 0.01 |
| L band; pol. HH | 0.04 | *** | 0.03 | *** | 0.09 | *** | 0.06 | *** | −0.03 | 0.06 | 0.00 | 0.98 |
| L band; pol. HV | 0.17 | *** | 0.14 | *** | 0.13 | *** | 0.09 | *** | 0.03 | 0.1 | 0.02 | 0.03 |
| C band; pol. VH | −0.65 | *** | −0.43 | 0.08 | −0.25 | *** | −0.14 | *** | −0.15 | *** | −0.05 | *** |
| C band; pol. VV | −0.64 | *** | −0.39 | 0.14 | −0.21 | *** | −0.11 | *** | −0.08 | *** | −0.01 | 0.27 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fagua, J.C.; Jantz, P.; Rodriguez-Buritica, S.; Duncanson, L.; Goetz, S.J. Integrating LiDAR, Multispectral and SAR Data to Estimate and Map Canopy Height in Tropical Forests. Remote Sens. 2019, 11, 2697. https://doi.org/10.3390/rs11222697
Fagua JC, Jantz P, Rodriguez-Buritica S, Duncanson L, Goetz SJ. Integrating LiDAR, Multispectral and SAR Data to Estimate and Map Canopy Height in Tropical Forests. Remote Sensing. 2019; 11(22):2697. https://doi.org/10.3390/rs11222697
Chicago/Turabian StyleFagua, J. Camilo, Patrick Jantz, Susana Rodriguez-Buritica, Laura Duncanson, and Scott J. Goetz. 2019. "Integrating LiDAR, Multispectral and SAR Data to Estimate and Map Canopy Height in Tropical Forests" Remote Sensing 11, no. 22: 2697. https://doi.org/10.3390/rs11222697