Association between Brain and Plasma Glutamine Levels in Healthy Young Subjects Investigated by MRS and LC/MS


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. MRS Acquisition
2.3. MRS Data Analysis
2.4. Biochemical Analysis, LC/MS
2.5. Statistics
3. Results
3.1. Spectral Quality Assessment for MRS in PCC and Cbll
3.2. Test–Retest Reproducibility of MRS Measurements
3.3. Correlations between Glu–Gln Concentrations in Plasma and Brain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MRS | magnetic resonance spectroscopy |
| LC/MS | liquid chromatography-mass spectrometry |
| SPECIAL | short-TE spin-echo full-intensity acquired localized single voxel spectroscopy |
| Gln | glutamine |
| Glu | glutamate |
| PCC | posterior cingulate cortex |
| Cbll | cerebellum |
| Glx | sum of Glu and Gln |
| MSG | monosodium glutamate |
| BBB | blood brain barrier |
| VOI | volume of interest |
| OVS | outer volume suppression |
| VAPOR | variable-pulse power and optimized relaxation delays |
| %CoV | percent coefficient of variance |
| CSF | cerebrospinal fluid |
| WM | white matter |
| GM | grey matter |
| SNR | signal-to-noise ratio |
| NAA | N-acetylaspartate |
| MM | macromolecules |
| Ala | alanine |
| Asp | aspartate |
| PCh | phosphocholine |
| Cr | creatine |
| PCr | phosphocreatine |
| GABA | γ-aminobutyric acid |
| GSH | glutathione |
| Gly | glycine |
| mI | myo-inositol |
| Lac | lactate |
| Scyllo | scyllo-inositol |
| Tau | taurine |
| Glc | glucose |
| NAAG | N-acetylaspartylglutamate |
| GPC | glycerophosphocholine |
| PE | phosphorylethanolamine |
| Ser | serine |
| 2Na·EDTA | disodium ethylenediaminetetraacetate |
| HPLC-ESI-MS | high-performance liquid chromatography and electrospray ionization mass spectrometry |
| tCr | sum of Cr and PCr |
| tNAA | sum of NAA and NAAG |
| tCho | sum of GPC and PCh |
| CRLB | Cramér–Rao lower bound |
References
- Smith, Q.R. Transport of glutamate and other amino acids at the blood-brain barrier. J. Nutr. 2000, 130, 1016s–1022s. [Google Scholar] [CrossRef] [PubMed]
- Oz, G.; Alger, J.R.; Barker, P.B.; Bartha, R.; Bizzi, A.; Boesch, C.; Bolan, P.J.; Brindle, K.M.; Cudalbu, C.; Dincer, A.; et al. Clinical proton MR spectroscopy in central nervous system disorders. Radiology 2014, 270, 658–679. [Google Scholar] [CrossRef] [PubMed]
- Bustillo, J.R.; Chen, H.; Jones, T.; Lemke, N.; Abbott, C.; Qualls, C.; Canive, J.; Gasparovic, C. Increased glutamine in patients undergoing long-term treatment for schizophrenia: A proton magnetic resonance spectroscopy study at 3 T. JAMA Psychiatry. 2014, 71, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Kubo, H.; Nakataki, M.; Sumitani, S.; Iga, J.I.; Numata, S.; Kameoka, N.; Watanabe, S.Y.; Umehara, H.; Kinoshita, M.; Inoshita, M.; et al. 1H-magnetic resonance spectroscopy study of glutamate-related abnormality in bipolar disorder. J. Affect. Disord. 2017, 208, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Takado, Y.; Igarashi, H.; Terajima, K.; Shimohata, T.; Ozawa, T.; Okamoto, K.; Nishizawa, M.; Nakada, T. Brainstem metabolites in multiple system atrophy of cerebellar type: 3.0-T magnetic resonance spectroscopy study. Mov. Disord. 2011, 26, 1297–1302. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.A. Glutamine and cancer. J. Nutr. 2001, 131, 2539s–2542s. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, Y.; Vail, G.; Chow, H.; Zhang, Y.; Louie, L.; Li, J.; Hart, R.P.; Plummer, M.R.; Herrup, K. Erratum to: The impact of glutamine supplementation on the symptoms of ataxia-telangiectasia: a preclinical assessment. Mol. Neurodegener. 2017, 12, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Lee, S.J.; Teunissen, C.E.; Pool, R.; Shipley, M.J.; Teumer, A.; Chouraki, V.; Melo van Lent, D.; Tynkkynen, J.; Fischer, K.; Hernesniemi, J.; et al. Circulating metabolites and general cognitive ability and dementia: Evidence from 11 cohort studies. Alzheimer’s dement. J. Alzheimer’s Assoc. 2018, 14, 707–722. [Google Scholar] [CrossRef]
- Cioccari, L.; Gautschi, M.; Etter, R.; Weck, A.; Takala, J. Further Concerns About Glutamine: A Case Report on Hyperammonemic Encephalopathy. Crit. Care Med. 2015, 43, 458–460. [Google Scholar] [CrossRef]
- Helling, G.; Wahlin, S.; Smedberg, M.; Pettersson, L.; Tjader, I.; Norberg, A.; Rooyackers, O.; Wernerman, J. Plasma Glutamine Concentrations in Liver Failure. PLoS ONE 2016, 11, e0150440. [Google Scholar] [CrossRef]
- Cruzat, V.; Macedo Rogero, M.; Noel Keane, K.; Curi, R.; Newsholme, P. Glutamine: Metabolism and Immune Function, Supplementation and Clinical Translation. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Shulman, Y.; Grant, S.; Seres, P.; Hanstock, C.; Baker, G.; Tibbo, P. The relation between peripheral and central glutamate and glutamine in healthy male volunteers. J. Psychiatry Neurosci. 2006, 31, 406–410. [Google Scholar]
- Yamaguchi, S. Basic properties of umami and effects on humans. Physiol. Behav. 1991, 49, 833–841. [Google Scholar] [CrossRef]
- Obayashi, Y.; Nagamura, Y. Does monosodium glutamate really cause headache? A systematic review of human studies. J. Headache Pain 2016, 17, 54. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Kelly, A.M.; Biswal, B.B.; Castellanos, F.X.; Milham, M.P. Functional connectivity of default mode network components: Correlation, anticorrelation, and causality. Hum. Brain Mapp. 2009, 30, 625–637. [Google Scholar] [CrossRef]
- Manto, M.; Bower, J.M.; Conforto, A.B.; Delgado-Garcia, J.M.; da Guarda, S.N.; Gerwig, M.; Habas, C.; Hagura, N.; Ivry, R.B.; Marien, P.; et al. Consensus paper: Roles of the cerebellum in motor control—The diversity of ideas on cerebellar involvement in movement. Cerebellum (London, England) 2012, 11, 457–487. [Google Scholar] [CrossRef]
- Buckner, R.L. The cerebellum and cognitive function: 25 years of insight from anatomy and neuroimaging. Neuron 2013, 80, 807–815. [Google Scholar] [CrossRef]
- Mekle, R.; Mlynarik, V.; Gambarota, G.; Hergt, M.; Krueger, G.; Gruetter, R. MR spectroscopy of the human brain with enhanced signal intensity at ultrashort echo times on a clinical platform at 3T and 7T. Magn. Reson. Med. 2009, 61, 1279–1285. [Google Scholar] [CrossRef] [Green Version]
- Dhamala, E.; Abdelkefi, I.; Nguyen, M.; Hennessy, T.J.; Nadeau, H.; Near, J. Validation of in vivo MRS measures of metabolite concentrations in the human brain. NMR Biomed. 2019, 32, e4058. [Google Scholar] [CrossRef]
- Xin, L.; Mekle, R.; Fournier, M.; Baumann, P.S.; Ferrari, C.; Alameda, L.; Jenni, R.; Lu, H.; Schaller, B.; Cuenod, M.; et al. Genetic Polymorphism Associated Prefrontal Glutathione and Its Coupling With Brain Glutamate and Peripheral Redox Status in Early Psychosis. Schizophr. Bull. 2016, 42, 1185–1196. [Google Scholar] [CrossRef]
- Tkac, I.; Andersen, P.; Adriany, G.; Merkle, H.; Ugurbil, K.; Gruetter, R. In vivo 1H NMR spectroscopy of the human brain at 7 T. Magn. Reson. Med. 2001, 46, 451–456. [Google Scholar] [CrossRef]
- Harris, A.D.; Puts, N.A.; Edden, R.A. Tissue correction for GABA-edited MRS: Considerations of voxel composition, tissue segmentation, and tissue relaxations. J. Magn. Reson. Imaging 2015, 42, 1431–1440. [Google Scholar] [CrossRef] [Green Version]
- Simpson, R.; Devenyi, G.A.; Jezzard, P.; Hennessy, T.J.; Near, J. Advanced processing and simulation of MRS data using the FID appliance (FID-A)-An open source, MATLAB-based toolkit. Magn. Reson. Med. 2017, 77, 23–33. [Google Scholar] [CrossRef]
- Provencher, S.W. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn. Reson. Med. 1993, 30, 672–679. [Google Scholar] [CrossRef]
- Wilson, M.; Andronesi, O.; Barker, P.B.; Bartha, R.; Bizzi, A.; Bolan, P.J.; Brindle, K.M.; Choi, I.Y.; Cudalbu, C.; Dydak, U.; et al. Methodological consensus on clinical proton MRS of the brain: Review and recommendations. Magn. Reson. Med. 2019, 82, 527–550. [Google Scholar] [CrossRef] [Green Version]
- Shimbo, K.; Kubo, S.; Harada, Y.; Oonuki, T.; Yokokura, T.; Yoshida, H.; Amao, M.; Nakamura, M.; Kageyama, N.; Yamazaki, J.; et al. Automated precolumn derivatization system for analyzing physiological amino acids by liquid chromatography/mass spectrometry. Biomed. Chromatogr. BMC 2010, 24, 683–691. [Google Scholar] [CrossRef]
- Yoshida, H.; Kondo, K.; Yamamoto, H.; Kageyama, N.; Ozawa, S.; Shimbo, K.; Muramatsu, T.; Imaizumi, A.; Mizukoshi, T.; Masuda, J.; et al. Validation of an analytical method for human plasma free amino acids by high-performance liquid chromatography ionization mass spectrometry using automated precolumn derivatization. J. Chromatogr. B Anal. Technol. Biomed. life sci. 2015, 998–999, 88–96. [Google Scholar] [CrossRef]
- Adachi, Y.; Shimodaira, Y.; Nakamura, H.; Imaizumi, A.; Mori, M.; Kageyama, Y.; Noguchi, Y.; Seki, A.; Okabe, Y.; Miyake, Y.; et al. Low plasma tryptophan is associated with olfactory function in healthy elderly community dwellers in Japan. BMC Geriatr. 2017, 17, 239. [Google Scholar] [CrossRef]
- Yokoi, T.; Watanabe, H.; Yamaguchi, H.; Bagarinao, E.; Masuda, M.; Imai, K.; Ogura, A.; Ohdake, R.; Kawabata, K.; Hara, K.; et al. Involvement of the Precuneus/Posterior Cingulate Cortex Is Significant for the Development of Alzheimer’s Disease: A PET (THK5351, PiB) and Resting fMRI Study. Front. Aging Neurosci. 2018, 10, 304. [Google Scholar] [CrossRef]
- Johnson, K.A.; Schultz, A.; Betensky, R.A.; Becker, J.A.; Sepulcre, J.; Rentz, D.; Mormino, E.; Chhatwal, J.; Amariglio, R.; Papp, K.; et al. Tau positron emission tomographic imaging in aging and early Alzheimer disease. Ann. Neurol. 2016, 79, 110–119. [Google Scholar] [CrossRef]
- Hawkins, R.A.; O’Kane, R.L.; Simpson, I.A.; Vina, J.R. Structure of the blood-brain barrier and its role in the transport of amino acids. J. Nutr. 2006, 136, 218s–226s. [Google Scholar] [CrossRef]
- Bagga, P.; Behar, K.L.; Mason, G.F.; De Feyter, H.M.; Rothman, D.L.; Patel, A.B. Characterization of cerebral glutamine uptake from blood in the mouse brain: Implications for metabolic modeling of 13C NMR data. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2014, 34, 1666–1672. [Google Scholar] [CrossRef]
- O’Donnell-Luria, A.H.; Lin, A.P.; Merugumala, S.K.; Rohr, F.; Waisbren, S.E.; Lynch, R.; Tchekmedyian, V.; Goldberg, A.D.; Bellinger, A.; McFaline-Figueroa, J.R.; et al. Brain MRS glutamine as a biomarker to guide therapy of hyperammonemic coma. Mol. Genet. Metab. 2017, 121, 9–15. [Google Scholar] [CrossRef]
- Forslund, A.H.; Hambraeus, L.; van Beurden, H.; Holmback, U.; El-Khoury, A.E.; Hjorth, G.; Olsson, R.; Stridsberg, M.; Wide, L.; Akerfeldt, T.; et al. Inverse relationship between protein intake and plasma free amino acids in healthy men at physical exercise. Am. J. Physiol. Endocrinol. Metab. 2000, 278, 857–867. [Google Scholar] [CrossRef]
- Jauhar, S.; McCutcheon, R.; Borgan, F.; Veronese, M.; Nour, M.; Pepper, F.; Rogdaki, M.; Stone, J.; Egerton, A.; Turkheimer, F.; et al. The relationship between cortical glutamate and striatal dopamine in first-episode psychosis: A cross-sectional multimodal PET and magnetic resonance spectroscopy imaging study. Lancet Psychiatry 2018, 5, 816–823. [Google Scholar] [CrossRef]
- Martinez-Hernandez, A.; Bell, K.P.; Norenberg, M.D. Glutamine synthetase: Glial localization in brain. Science (New York, N.Y.) 1977, 195, 1356–1358. [Google Scholar] [CrossRef]
- Dolgodilina, E.; Imobersteg, S.; Laczko, E.; Welt, T.; Verrey, F.; Makrides, V. Brain interstitial fluid glutamine homeostasis is controlled by blood-brain barrier SLC7A5/LAT1 amino acid transporter. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2016, 36, 1929–1941. [Google Scholar] [CrossRef]
- Aldridge, D.R.; Tranah, E.J.; Shawcross, D.L. Pathogenesis of hepatic encephalopathy: Role of ammonia and systemic inflammation. J. Clin. Exp. Hepatol. 2015, 5, s7–s20. [Google Scholar] [CrossRef]
- Olabarria, M.; Noristani, H.N.; Verkhratsky, A.; Rodriguez, J.J. Age-dependent decrease in glutamine synthetase expression in the hippocampal astroglia of the triple transgenic Alzheimer’s disease mouse model: Mechanism for deficient glutamatergic transmission? Mol. Neurodegener. 2011, 6, 55. [Google Scholar] [CrossRef]
- Madeira, C.; Vargas-Lopes, C.; Brandao, C.O.; Reis, T.; Laks, J.; Panizzutti, R.; Ferreira, S.T. Elevated Glutamate and Glutamine Levels in the Cerebrospinal Fluid of Patients With Probable Alzheimer’s Disease and Depression. Front. Psychiatry 2018, 9, 561. [Google Scholar] [CrossRef]
- Smith, C.C.; Bowen, D.M.; Francis, P.T.; Snowden, J.S.; Neary, D. Putative amino acid transmitters in lumbar cerebrospinal fluid of patients with histologically verified Alzheimer’s dementia. J. Neurol. Neurosurg. Psychiatry 1985, 48, 469–471. [Google Scholar] [CrossRef]
- Procter, A.W.; Palmer, A.M.; Francis, P.T.; Lowe, S.L.; Neary, D.; Murphy, E.; Doshi, R.; Bowen, D.M. Evidence of glutamatergic denervation and possible abnormal metabolism in Alzheimer’s disease. J. Neurochem. 1988, 50, 790–802. [Google Scholar] [CrossRef]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake into astrocytes stimulates aerobic glycolysis: A mechanism coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef]
- Bowman, G.L.; Kaye, J.A.; Moore, M.; Waichunas, D.; Carlson, N.E.; Quinn, J.F. Blood-brain barrier impairment in Alzheimer disease: Stability and functional significance. Neurology 2007, 68, 1809–1814. [Google Scholar] [CrossRef] [Green Version]



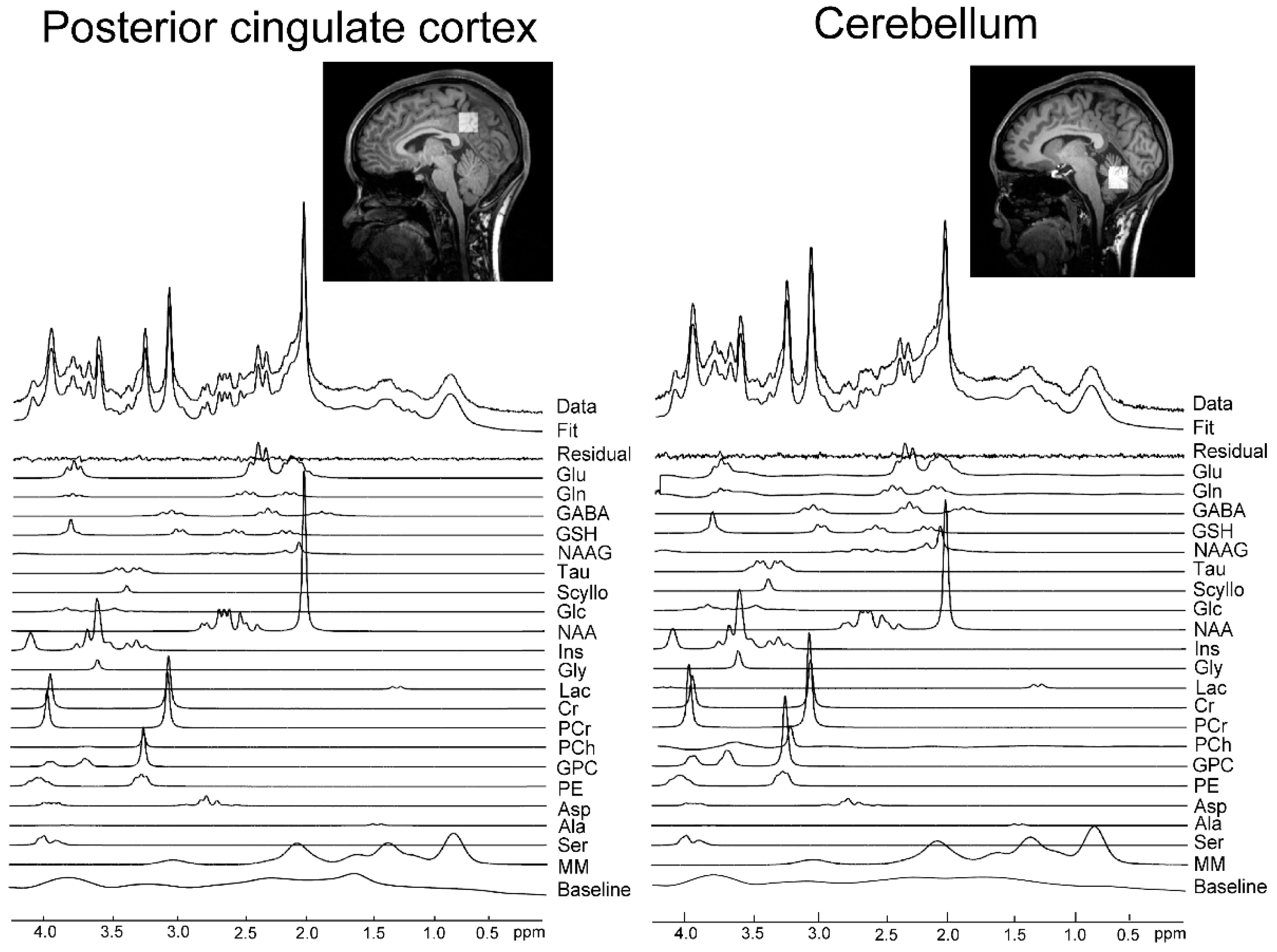{kind=link}
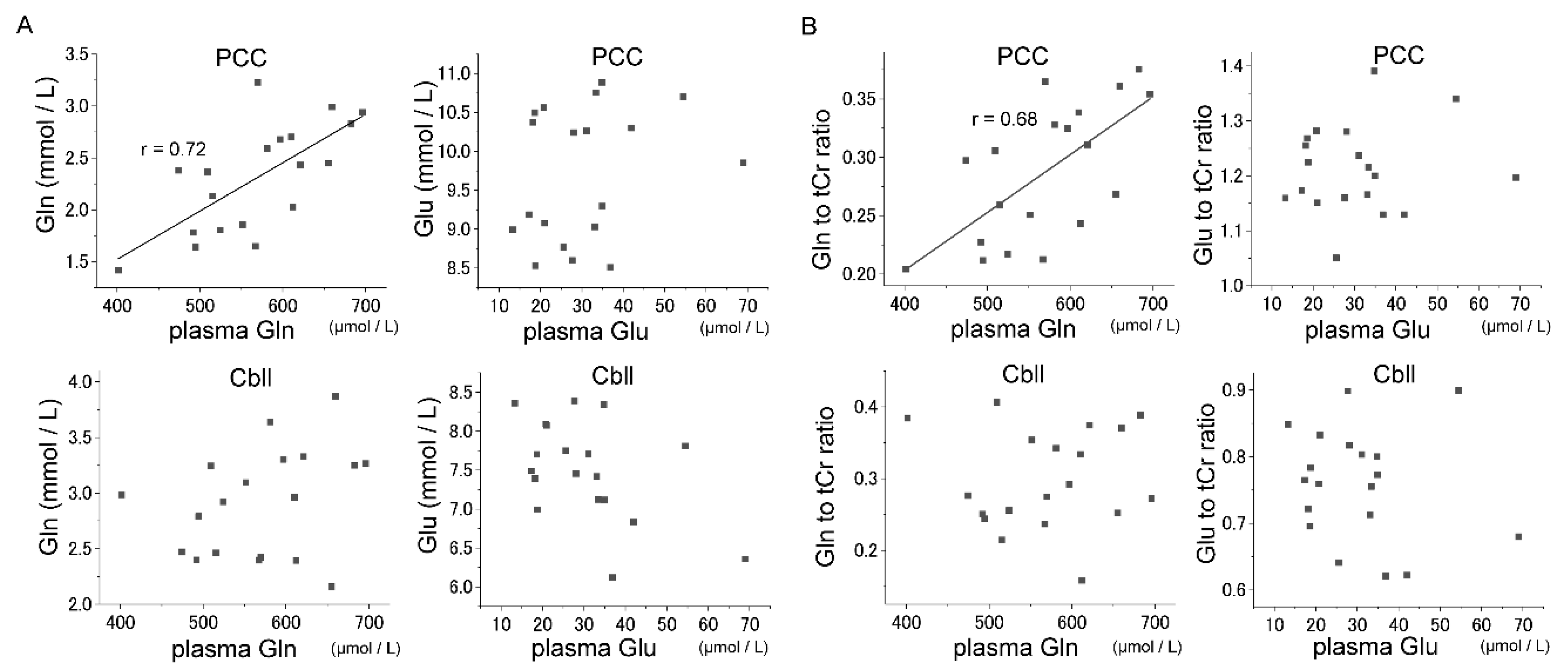{kind=link}
| PCC | Cbll | |||
|---|---|---|---|---|
| Metabolites | Mean (%) | SD | Mean (%) | SD |
| tCr | 1.5 | 0.5 | 1.3 | 0.5 |
| GABA | 10.7 | 1.1 | 9.7 | 2.2 |
| Gln | 10.9 | 1.6 | 9.3 | 1.8 |
| Glu | 3.1 | 0.3 | 4 | 0.7 |
| GSH | 6.2 | 0.7 | 5.8 | 1.2 |
| mI | 4.4 | 0.7 | 4.1 | 0.7 |
| tNAA | 1.5 | 0.5 | 1.7 | 0.5 |
| tCho | 3.3 | 0.5 | 2.4 | 0.5 |
| PCC | Cbll | |
|---|---|---|
| Mean (%) | Mean (%) | |
| corrected Gln | 9.4 | 14.7 |
| corrected Glu | 5.0 | 8.7 |
| corrected tCr | 3.5 | 3.7 |
| Gln/tCr | 7.6 | 15.2 |
| Glu/tCr | 2.3 | 6.2 |
| PCC 1st | PCC 2nd | Cbll 1st | Cbll 2nd | |||||
|---|---|---|---|---|---|---|---|---|
| Mean (%) | SD | Mean (%) | SD | Mean (%) | SD | Mean (%) | SD | |
| GM | 75.6 | 3.1 | 75.9 | 3.2 | 67.0 | 3.4 | 67.0 | 3.8 |
| WM | 12.9 | 2.9 | 13.7 | 2.4 | 29.2 | 3.9 | 30.0 | 4.2 |
| CSF | 11.6 | 3.5 | 10.4 | 3.3 | 3.9 | 2.8 | 3.0 | 2.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takado, Y.; Sato, N.; Kanbe, Y.; Tomiyasu, M.; Xin, L.; Near, J.; Yoshikawa, K.; Sahara, N.; Higashi, T.; Suhara, T.; et al. Association between Brain and Plasma Glutamine Levels in Healthy Young Subjects Investigated by MRS and LC/MS. Nutrients 2019, 11, 1649. https://doi.org/10.3390/nu11071649
Takado Y, Sato N, Kanbe Y, Tomiyasu M, Xin L, Near J, Yoshikawa K, Sahara N, Higashi T, Suhara T, et al. Association between Brain and Plasma Glutamine Levels in Healthy Young Subjects Investigated by MRS and LC/MS. Nutrients. 2019; 11(7):1649. https://doi.org/10.3390/nu11071649
Chicago/Turabian StyleTakado, Yuhei, Naoto Sato, Yuta Kanbe, Moyoko Tomiyasu, Lijing Xin, Jamie Near, Kohki Yoshikawa, Naruhiko Sahara, Tatsuya Higashi, Tetsuya Suhara, and et al. 2019. "Association between Brain and Plasma Glutamine Levels in Healthy Young Subjects Investigated by MRS and LC/MS" Nutrients 11, no. 7: 1649. https://doi.org/10.3390/nu11071649