MALAT1 Long Non-Coding RNA: Functional Implications


1
Envisagenics, 101 Avenue of the Americas, New York, NY 10013, USA
2
Graduate Program in Genetics, Stony Brook University, Stony Brook, New York, NY 11794, USA
3
Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, NY 11724, USA
*
Author to whom correspondence should be addressed.
Non-Coding RNA 2020, 6(2), 22; https://doi.org/10.3390/ncrna6020022
Submission received: 11 April 2020
/
Revised: 27 May 2020
/
Accepted: 29 May 2020
/
Published: 3 June 2020
(This article belongs to the Special Issue Non-Coding RNA and Intracellular Structures)
{kind=link}
{kind=link}
Abstract
:The mammalian genome is pervasively transcribed and the functional significance of many long non-coding RNA (lncRNA) transcripts are gradually being elucidated. Metastasis Associated Lung Adenocarcinoma Transcript 1 (MALAT1) is one of the most well-studied lncRNAs. MALAT1 is a highly conserved nuclear retained lncRNA that is abundantly expressed in cells and tissues and has been shown to play a role in regulating genes at both the transcriptional and post-transcriptional levels in a context-dependent manner. However, Malat1 has been shown to be dispensable for normal development and viability in mice. Interestingly, accumulating evidence suggests that MALAT1 plays an important role in numerous diseases including cancer. Here, we discuss the current state-of-knowledge in regard to MALAT1 with respect to its function, role in diseases, and the potential therapeutic opportunities for targeting MALAT1 using antisense oligonucleotides and small molecules.
1. Introduction
The eukaryotic genome is transcribed into tens of thousands of non-coding RNAs adding additional complexity to the regulatory framework of cells and organisms. Long non-coding RNAs (lncRNAs) are emerging as a critical class of transcripts participating in a wide range of cellular functions. The current Gencode annotation estimates that there are at least 17,952 lncRNA genes (48,438 transcripts) in humans and 13,197 lncRNA genes (18,864 transcripts) in mice [1]. These genes are transcribed by RNA polymerase II and exhibit classical promoter and enhancer elements. The majority are capped, spliced, and polyadenylated, although some are single exon transcripts. Many lncRNAs undergo alternative pre-mRNA splicing and/or alternative cleavage and polyadenylation leading to multiple isoforms from the same locus [2,3]. While almost all eukaryotic species encode lncRNA genes, conservation of lncRNAs has been a topic of intense debate for over a decade [4,5,6]. Many lncRNAs show poor sequence level conservation, with some of them demonstrating structural conservation to some degree. In many instances, orthologs are identified by syntenic conservation followed by functional rescue experiments [7].
Despite the large number of annotated lncRNAs, a large number (>80%) are expressed at very low levels, few transcripts per cell, and the majority of these transcripts are restricted to just one or a few cell types and/or specific developmental stages or physiologic conditions [8,9]. In addition to cell type specific expression, many lncRNAs also display a specific sub-cellular localization being restricted to cytoplasmic or nuclear compartments or association with specific organelles, such as polycomb bodies, stress granules, nuclear speckles, paraspeckles, etc. [10]. More recent evidence suggests that lncRNAs actively contribute to phase separation in large ribonucleoprotein complexes giving rise to membrane-less organelles inside the cell [11,12,13,14]. Given the remarkable diversity of this class of RNAs, it is reasonable to envision that they contribute in a structural, functional, and/or regulatory capacity in a wide range of cellular/nuclear processes [15]. Metastasis Associated Lung Adenocarcinoma Transcript 1 (MALAT1) is one of the most widely studied nuclear restricted lncRNAs that has gained much attention in recent years due to its abundance, rather ubiquitous expression, and apparent role in various disease manifestations. Here, we discuss the current state-of-knowledge in regard to MALAT1 function and its putative role in several disease states, including cancer.
2. MALAT1–Basic Characteristics
MALAT1, also known as Nuclear Enriched Abundant Transcript 2 (NEAT2) [16] was first identified in a microarray screen of tumors from patients with non-small cell lung cancer, and was found to be upregulated in the tumors with a higher propensity to metastasize [17]. The MALAT1 gene is encoded on human chromosome 11q13.1 and mouse chromosome 19qA. It is located in a gene dense region with a very high syntenic evolutionary conservation [8]. MALAT1 exhibits a remarkable sequence conservation with greater than 50% overall conservation in vertebrates and greater than 80% conservation at the 3′ end of the transcript [18,19,20]. This is one of the key distinguishing features of MALAT1 as very few lncRNAs show such a high level of evolutionary conservation. Less than 10% of all lncRNAs show exonic as well as structural conservation equivalent to that of protein coding genes [21,22]. The MALAT1 transcript has been confirmed as a non-coding RNA as it exhibits low protein-coding potential using two independent coding potential calculating algorithms CPC2 [23] and CPAT [24].
Human MALAT1 is ~8.7 knt long, whereas the mouse RNA is 6.7 knt long [8]. It is transcribed by RNA polymerase II and its promoter has an accessible open chromatin architecture, which has been shown in several high-throughput studies and DNAse sensitivity assays [8]. The steady state expression level of MALAT1 is very high and is comparable to highly transcribed housekeeping genes, such as β-Actin [8]. Further, MALAT1 is ubiquitously expressed across all tissues with an average median expression of about 150 TPM (transcripts per million) with highest expression in ovaries with a median expression of 287 TPM [25]. The abundance of MALAT1 in cells is likely the consequence of strong promoter activity coupled with increased stability of the transcribed RNA [26,27]. MALAT1 was originally classified as an intron-less transcript with a genomically-encoded poly A tract. However, with a number of deep sequencing efforts, several alternatively spliced isoforms and transcripts with alternative transcription start sites have been identified that are expressed during different physiological states such as cancer [8,28]. In addition, a natural antisense transcript TALAM1 has also been identified and has been suggested to play a role in a feed-forward positive regulatory loop to maintain the high cellular levels of MALAT1 and contribute to its stability [29]. Interestingly, simultaneous knockdown of both MALAT1 and TALAM1 in breast cancer cells led to a stronger, synergistic decrease in migration and invasion of these cells and reduced metastasis to the lungs in a mouse model [30].
The MALAT1 primary transcript is processed to yield the well characterized nuclear retained MALAT1 transcript, and from its 3′ end a tRNA-like small RNA [27]. The biogenesis of the small RNA is mediated by the tRNA processing machinery, RNase P and RNase Z. The 61-nucleotide tRNA- like MALAT1-associated small cytoplasmic RNA (mascRNA) is exported to the cytoplasm [27]. The resultant 3′ end of the nuclear MALAT1 transcript post-processing is not polyadenylated, however, it contains a genomically-encoded poly(A)-rich stretch which pairs with an upstream U-rich region and then adopts a unique triple helical confirmation [31,32,33,34]. This triple helical structure was first identified in the PAN (polyadenylated nuclear) RNA produced by the human oncogenic Kaposi sarcoma-associated γ-herpesvirus (KSHV) PAN RNA [35,36]. The only other human or mouse RNA that exhibits such a structure is the ~20 knt Men-β (NEAT1_2) RNA [33,34]. The triple helical structure has been shown to confer stability and nuclear localization to MALAT1 in the absence of a true poly(A) tail and has been shown to bind several RNA binding proteins (RBPs) including METTL16 which is an m6A RNA methyl-transferase [33,37]. Specific triplex-disrupting mutations lead to MALAT1 degradation and loss of nuclear accumulation [38]. Further, a class of RNAs containing a similar 3′ end including a triple helix structure have been identified in several other species including lizards, where they have been shown to play a role in piRNA biogenesis in testicular cells [32]. Taken together, the MALAT1 locus displays remarkable evolutionarily conserved secondary and tertiary structural features and an unusual 3′ end processing mechanism. It is not fully apparent whether the full length MALAT1 RNA with its 3′ end triplex structure, the processed tRNA-like RNA, and the natural antisense RNA have a concerted mechanism of action or if each component derived from this interesting locus has a disparate function. Further, high-throughput chemical mapping experiments have highlighted extensive epi-transciptomic changes in the MALAT1 transcript, for example, m6A, pseudouridylation and 5-methyl cytosine [39,40,41]. It has been shown that the addition of m6A at the A2577 position could destabilize the hairpin stem of MALAT1, making it accessible for RNA-binding proteins such as HNRNPC [42]. Additional detailed molecular studies elucidating the transcriptional and post-transcriptional regulation of the MALAT1 locus will address these issues and allow us to further understand the regulation and function of the MALAT1 locus.
3. MALAT1 Localization
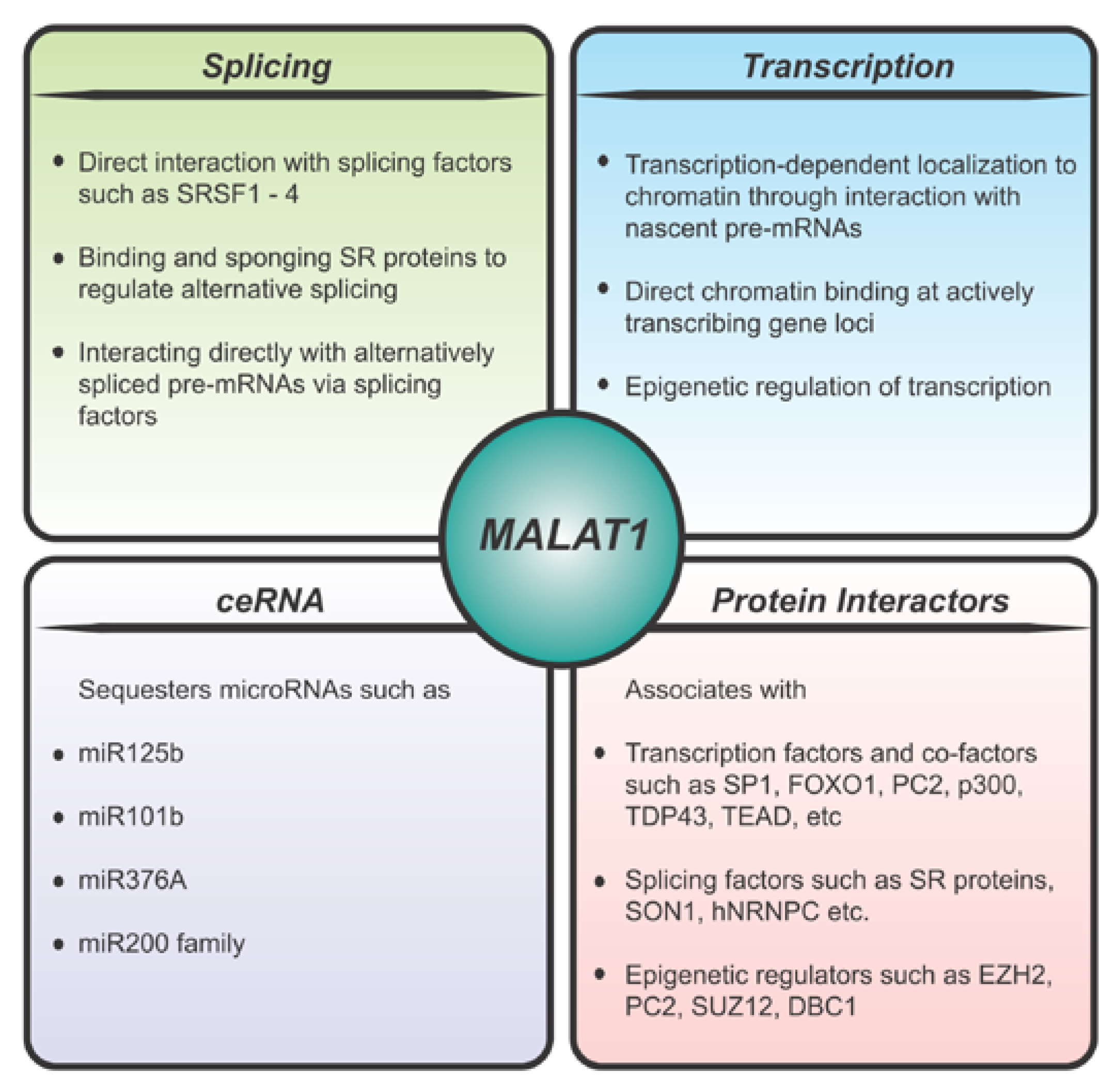
MALAT1 is a nuclear retained RNA that localizes to nuclear domains known as nuclear speckles [16]. Nuclear speckles are enriched in pre-mRNA processing factors, as well as some transcription factors, and play a critical role in coordinating transcriptional and post-transcriptional gene regulation [43]. MALAT1 has been shown to be enriched at the periphery of the nuclear speckles with pre-mRNA splicing factors localized more internally [44]. The previously described triple-helix element and other MALAT1 domains have been shown to facilitate the nuclear retention and localization of MALAT1 [31,45]. Although Malat1 localizes to nuclear speckles it is not required for the formation of nuclear speckles [16] and knockout of Malat1 has no overall effect on the assembly, number, size, distribution, or maintenance of nuclear speckles [46]. As such Malat1 is not a candidate to play a role in regulating the process of phase separation in the formation of nuclear speckles. Several nuclear speckle components such as, RNPS1, SRm160, and IBP160, were found to be essential factors for the localization of MALAT1 to nuclear speckles, and the proper localization was found to be mediated by two distinct regions of MALAT1 (1777–3600 nt and 5185–6982 nt) [45,47]. In addition, Malat1 has been shown to bind to several other pre-mRNA splicing factors that are enriched in nuclear speckles such as SRSF1, SON1, hnRNPC, hnRNPH1, etc. (Figure 1) [47,48,49]. MALAT1 has also been shown to modulate recruitment of splicing factors to actively transcribing loci in human cell lines [26,47,50] thereby regulating alternative pre-mRNA splicing of a number of pre-mRNAs (Figure 1). Further, CRISPR screening studies, identified both positive (DHX15, DDX4,2, hnRNPH1 and hnRNPK) and negative (hnRNPA1, hnRNPL, and PCBP1) regulators of the nuclear speckle localization of MALAT1. It was suggested that negative regulators could compete with the factors that recruit MALAT1 to nuclear speckles thereby dissociating MALAT1 from nuclear speckles upon transcriptional inhibition suggesting a role for MALAT1 in transcriptional regulation [51]. In addition to the above mentioned components contributing to the localization of Malat1, recently it has been shown that a SINE element in the Malat1 5′ end associates with HNRNPK, KHDRBS1, and TRA2A contributing to its nuclear localization [52]. A MALAT1 SINE deletion mutant localizes diffusely in the nucleus and is frequently transported to the cytoplasm resulting in the formation of cytotoxic insoluble TDP-43 inclusions in both the cytoplasm and nucleus [52].
In addition to its nuclear speckle localization, MALAT1 has also been shown to be associated with chromatin. High-throughput chromatin-RNA binding assays such as CHART and ChIRP [53,54] have identified MALAT1 as a highly enriched RNA in the chromatin fraction, where it has been shown to be associated with transcriptionally active genes. In another study, MALAT1 was shown to coordinate the relocation of genes from polycomb bodies to transcriptionally active sites in a serum responsive manner. This mechanism was directed through binding to several members of the polycomb group proteins such as PC2, EZH2, and SUZ12, thereby regulating the transcriptional status of a number of PRC2 target genes by relieving their repression (Figure 1) [55,56,57]. Further, it has been shown that MALAT1 can target CTCF binding sites and active promoters [58]. Using CHIA-PET technology, it was shown that 3D genome organization impacts MALAT1 binding to target genes and MALAT1 binding sites were involved in both CTCF- and RNAPII-mediated chromatin interactions [58]. Specifically, such a long-range interaction was shown on the LTBP3 promoter which has been previously shown to be regulated by MALAT1 [59]. Additional high-throughput experiments, such as MARGI and GRID-seq, have also revealed extensive binding of MALAT1 to thousands of genomic loci in a cell-type specific manner [60,61].
4. Molecular Function of MALAT1
Numerous mechanisms of action have been proposed to explain the role of MALAT1 (Figure 1) in a wide range of physiological states (Figure 2). A significant number of studies have supported a function for MALAT1 based on its defined subnuclear localization and have proposed that MALAT1 either plays a role in transcription, directly or indirectly, and/or regulates alternative pre-mRNA splicing [62]. The splicing role for MALAT1 is directly related to its localization in nuclear speckles, a sub-nuclear body enriched in pre-mRNA splicing factors [16,26,47]. A number of studies have demonstrated altered pre-mRNA splicing upon MALAT1 knockdown in cells [47,63,64]. In addition, MALAT1 has been shown to regulate the phosphorylation status of an SR splicing factor thereby regulating its speckle localization and its role in alternative pre-mRNA splicing [47]. Other studies have shown that MALAT1 may directly participate in pre-mRNA splicing of actively transcribed genes by recruiting splicing factors to the pre-mRNA [65]. In addition to those factors described in the previous section, MALAT1 has been shown to bind to several SR proteins such as SRSF1, SRSF3, SRSF2, and other RBPs, such as HNRNPL and TDP-43 [47,49,64,66,67]. Taken together these studies indicate a role for MALAT1 directly or indirectly in the regulation of pre-mRNA splicing.
A number of studies have also shown a role for MALAT1 at the transcriptional level (Figure 1). For example, in vivo cross-linking studies have shown MALAT1 binding to chromatin of actively transcribing genes and regulating their expression at the transcriptional level [53]. In addition, MALAT1 also binds to a number of transcription factors and transcriptional co-activators, such as LTBP3, FOXO1, PC2, HMGA2, etc. [55,59,68,69]. Using RNA reverse transcription-associated trap sequencing (RAT-seq), MALAT1 was shown to increase proliferation and migration of breast cancer cells via binding to the EEF1A1 promoter and upregulating its expression epigenetically [70]. MALAT1 has also been shown to epigenetically upregulate transcriptional activators of proteosome subunit genes in multiple myeloma cells [71]. MALAT1 has been shown to bind DBC1 causing deacetylation of p53, thus promoting cell proliferation and inhibiting cell apoptosis [49]. Overexpression or knockdown of MALAT1 in mammalian cells under a wide range of physiological conditions influences transcriptional changes in a context specific manner [63,70,72,73,74,75]. Based on multiple lines of evidence MALAT1 may likely influence both transcription as well as pre-mRNA splicing (Figure 1). With more recent studies demonstrating transcription-coupled splicing in determining alternative splice-site choice and alternative polyadenylation [76,77], it is tempting to speculate that MALAT1 may directly coordinate these events in a context specific manner by either scaffolding the protein complexes or acting as a chaperone targeting the transcription/pre-mRNA splicing machinery to the appropriate genes.
In addition to influencing splicing and transcription, MALAT1 has been shown to act as a competing endogenous RNA (ceRNA) or miRNA sponge to sequester miRNAs under various conditions (Figure 1). For example, it has been shown that miR-125b can bind to MALAT1 to downregulate its expression and inhibit bladder cancer development [78]. It has also been reported that MALAT1 regulates Rac1 expression by acting as a ceRNA for miR-101b in liver fibrosis [79]. MALAT1 has also been shown to promote development of osteosarcoma by targeting TGFA via miR-376A [80]. Further, MALAT1 has been shown to induce EMT during endometriosis [81] and metastasis in clear cell kidney carcinoma mouse models [82] via the miR200/Zeb axis by acting as a sponge for the miR200 family. While these studies allude to a role of MALAT1 in sequestering miRNAs, it is not clear if this is attributed to the nuclear function of MALAT1 as the majority of these miRNAs are enriched in the cytoplasmic compartment. Perhaps MALAT1 sequesters miRNAs in the early nuclear stage of pre-miRNA processing as MALAT1 does not appear to shuttle between the nuclear and cytoplasmic compartments. For a better understanding of these processes, the context of MALAT1 function and localization needs to be more thoroughly investigated.
5. MALAT1 is Dispensable for Normal Physiology
While it is compelling to speculate that MALAT1 plays a very critical cellular function, three independent knockout (KO) mouse models generated by different groups concluded that loss of Malat1 did not have an impact on normal mouse physiology or development [46,83,84]. In these mouse KO models generated using different strategies, Malat1 loss did not affect the normal development of the mice and adult mice did not exhibit any aberrant phenotypes. Zhang et al. have reported a 1.5–2-fold upregulation of several genes neighboring Malat1 in brain tissues upon KO of a 3.5 kb region surrounding the Malat1 promoter suggesting a cis acting role for Malat1. However, this upregulation was not observed in other tissues, including mammary tissue (unpublished data). A second group reported a modest down-regulation of the Neat1 transcript in intestines of Malat1 KO mice [84]. A third group reported KO of Malat1 did not affect proliferation or cell cycle progression in human lung or liver cancer cells. In addition, a KO Malat1 mouse did not result in any obvious phenotype or histological abnormality [83]. One logical explanation of these findings is that there may be functional redundancy for the Malat1 RNA under normal physiologic conditions as is the case for many critical genes [85,86].
6. MALAT1 and Cancer
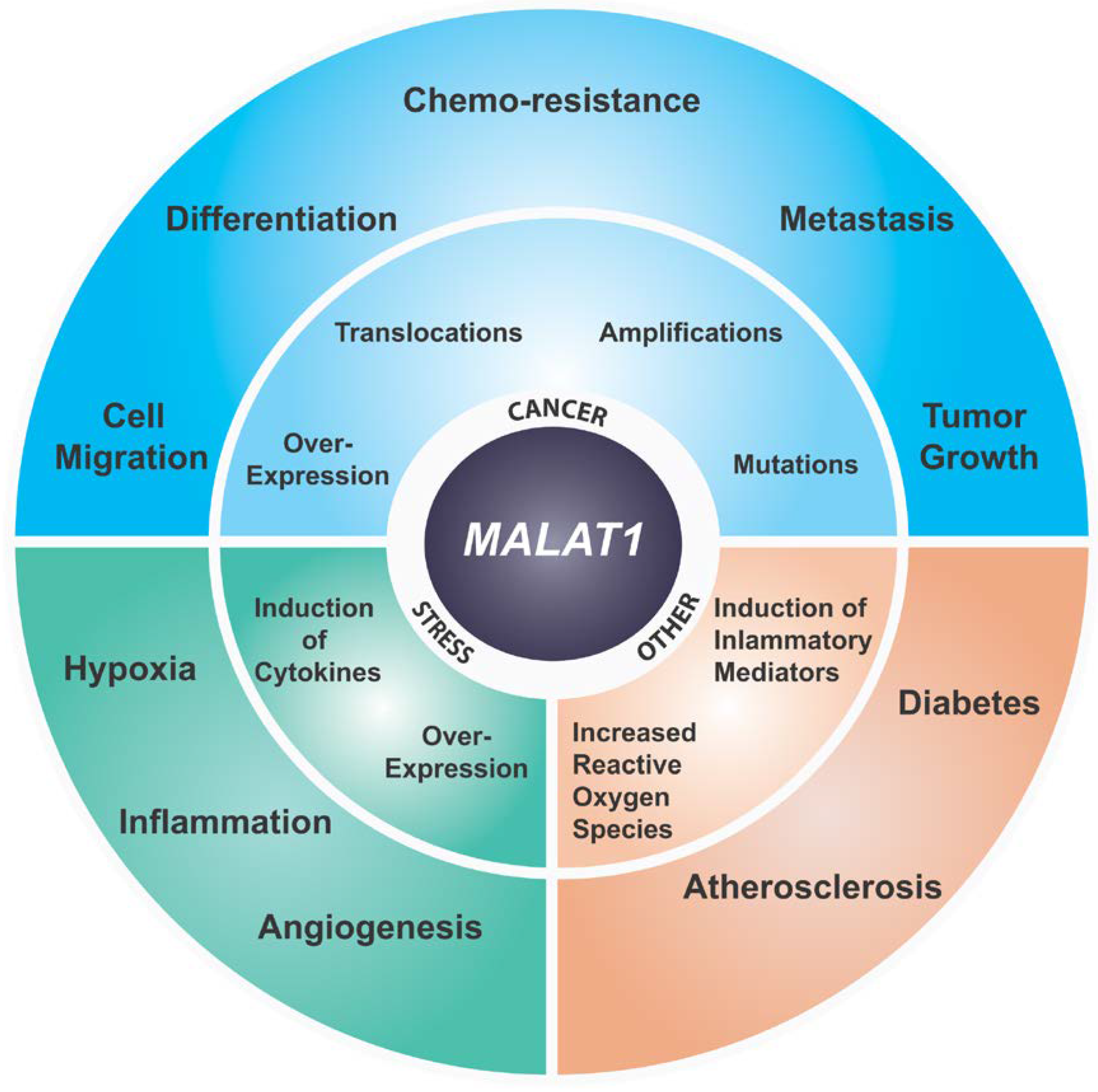
MALAT1 was initially identified as an RNA whose expression is elevated in primary lung tumor that had a higher propensity to metastasize [17]. Since this initial study overexpression of this lncRNA has been reported in over 20 different solid or lymphoid tumors specifically correlating its higher expression to tumor progression and metastasis (Figure 2) [17,63,71,87,88,89,90,91,92,93,94,95,96,97,98,99]. Depending on the type and stage of cancer, the relative upregulation was found be from 1.5–10 fold [17,63,100,101]. Higher expression of MALAT1 has been shown to be associated with poor prognosis in a variety of solid cancers and hematopoietic cancers [102,103,104,105]. Additionally, MALAT1 overexpression has been associated with metastasis in lung, breast and liver cancers [17,63,64,102]. Malat1 loss or knockdown in a murine metastatic cancer model resulted in differentiation of primary tumors and a significant reduction in metastasis [63]. Additionally, both Malat1 knockdown and genetic KO in a lung cancer homing model reduced homing to the lungs of lung cancer cells [101]. Similar observations have been reported in CRC, esophageal carcinoma, gallbladder, cervical cancer, and prostate cancer where knockdown of MALAT1 abrogated tumor growth and/or metastasis in the respective cell line-derived models and/or PDX mouse models [69,87,106,107,108,109]. In many of these studies MALAT1 knockdown affected transcription and/or pre-mRNA splicing of critical genes involved in migration and cell adhesion in addition to genes involved in critical cancer pathways. Additionally, overexpression of the 5′ Malat1 fragment was found to be sufficient to transform mouse primary embryonic fibroblast cells resulting in increased colony formation in soft a gar assays [110]. Interestingly, Gao et al. have demonstrated that expression of the Malat1 5′ region can induce metastasis in the non-metastatic 4T07 murine mammary cancer cell line suggesting a gain of function for the Malat1 5′ fragment in promoting metastasis [111]. In addition, MALAT1 overexpression has been shown to be associated with drug resistance in breast cancer, CRC, prostate cancer, etc. (Figure 2) [112,113,114]. Despite accumulating evidence that MALAT1 plays a pro-oncogenic and pro-metastatic role in a wide range of cancers, including mammary cancer, as discussed above, a few recent studies reported a tumor suppressor-like role for Malat1 [74,115,116]. The significance of these later findings is unclear as they are contradictory to a large body of data supporting a pro-oncogenic role for MALAT1. Additional studies are necessary to clarify these differences [117].
MALAT1 is a highly-conserved noncoding RNA gene transcribed from the human 11q13 locus which has been shown to exhibit copy number changes, translocations, or mutations in several cancer types. MALAT1 translocation to TFEB has been reported in renal cell carcinoma [118]. Translocation of the 5′ region of MALAT1 to Gli1 has been found to be oncogenic in an aggressive form of Gastroblastoma [119]. Further, the MALAT1 locus was shown to exhibit tandem duplication in some breast cancers resulting in increased dosage of the gene [120]. Apart from chromosomal aberrations in the MALAT1 locus in cancer, WGS studies from patient tumors have found that MALAT1 is a frequently mutated gene in breast and other cancer types [121,122]. A number of hotspot mutations have been identified in the MALAT1 gene that are mostly clustered in the 3 kb–4.3 kb region, although the role of such short indels and point mutations in this gene is unclear [122]. Recent PCWGA suggests that MALAT1 mutations may be a consequence of the high level of transcription associated with the gene and an inherently fragile genomic locus, and may not necessarily represent driver mutations [123,124]. However, additional studies are warranted to assess whether these aberrations may interfere with the above functions of MALAT1 or represent mutations that promote tumorigenesis.
Finally, meta-analysis of transcriptomic datasets has also shown MALAT1 to be upregulated in several cancer tissues such as lung, CRC, prostate, breast, etc. cancer compared to normal tissues [125]. Analysis of TCGA data from breast, lung, prostate, and glioma cancers have identified overexpression of MALAT1 associated with poor prognosis and reduced metastasis-free survival [102,103,104,105]. Higher levels of MALAT1 have also been observed in circulating RNAs, and also RNAs extracted from exosomes from cancer patients [126,127,128]. MALAT1 levels in urine and urinary exosomes have been evaluated in prostate and bladder cancer respectively for developing MALAT1 as a non-invasive prognostic biomarker [129,130]. It is compelling that this wide range of studies have identified MALAT1 as being strongly enriched in various body fluids of cancer patients and warrants MALAT1 to be further evaluated as a potential prognostic or diagnostic marker. However, the specificity of such a diagnostic assay will be challenging, as MALAT1 is also an abundant RNA in most normal tissues, which can contribute to significant noise in such analysis.
7. MALAT1 and Stress Responses
While no apparent phenotype has been observed upon Malat1 loss in knockout mice, differential expression of MALAT1 has been reported under various physiological stresses such as serum starvation, hypoxia etc. [55,64,131]. Additionally, it has been shown that MALAT1 enhances glycolysis, and inhibits gluconeogenesis, via elevated translation of the transcription factor TCF7L2 and as such also plays a role in metabolic stress [132]. Knockout of other important genes, such as RPL, Cyclin D, etc., show a lack of phenotype under normal conditions due to functional redundancy, whereas upon physiological stress they manifest a phenotype [85,86]. Consistent with this hypothesis, Malat1 KO mice crossed with breast tumor bearing models display a tumor differentiation phenotype [63]. In addition, MALAT1 localization and function has been shown to be altered during serum starvation [55]. Malat1 was also demonstrated to be induced in kidneys of hypoxic mice [133], and Malat1 was identified as one of the most upregulated non-coding transcripts upon hypoxia in a breast cancer cell line [134]. MALAT1 has been shown to be regulated by HIF1α, a key transcription factor during the hypoxic response [134,135]. MALAT1 knockdown also influences the expression of proangiogenic isoforms of VEGFa which is a classic HIF1α regulated gene [73]. In multiple myeloma, MALAT1 was shown to be a target of KDM3A, whose upregulation resulted in accumulation of HIF-1α, and induction of glycolytic genes under hypoxia conditions [136]. An additional study has demonstrated that cancer cell-specific chromatin-chromatin interactions are formed at the MALAT1 locus under hypoxic stress, thereby implicating a novel role of MALAT1 in regulating hypoxic response in cancer [131]. Collectively, these studies indicate a direct role for MALAT1 in hypoxic stress which is responsible for significant pathological consequences in cancer including angiogenesis and metastasis.
MALAT1 has also been shown to play a critical role in regulating the A-NHEJ pathway during B cell class switch recombination [137]. Further, several studies have identified MALAT1 as a regulator of TRP53 [75,138] and knockdown of MALAT1 was shown to result in increased H2Ax foci [75] suggesting that MALAT1 plays a more general role in the double-strand break response and genotoxic stress. Chemotherapeutic agents are known to cause genotoxic stress and, interestingly, MALAT1 was significantly upregulated by chemotherapeutic agents in extramedullary myeloma suggesting that it could be a stress responsive gene [139]. MALAT1 has also been shown to be a target for chemo-sensitization of GBM wherein it is regulated by members of the TP53 family [104,140]. Similar observations of upregulation of MALAT1 has been reported in drug resistance phenotypes in lung, prostate and other cancers [112,141,142,143]. This is a rather intriguing observation as many studies mentioned above have observed a strong correlation between MALAT1 expression and the development of chemo-resistance in cancer. Further investigations along this line are warranted in order to understand the role of MALAT1 in the development of the drug resistance phenotype in cancers and to identify potential combinatorial therapeutic opportunities to target MALAT1 to augment chemotherapeutic response.
8. MALAT1 in Other Diseases
In addition to cancer, studies have identified MALAT1 upregulation in a wide range of other pathological indications as summarized in Figure 2 [144]. A significant number of studies have directly implicated MALAT1 in development of diabetes and insulin signaling. An early study identified MALAT1 upregulation in endothelial cells subjected to high glucose treatment [145]. MALAT1 was also found to play an important role in regulating insulin sensitivity by regulating NRF2 activity and suppressing JNK signaling with concomitant insulin-induced phosphorylation of Akt [146]. Additionally, a novel signaling nexus involving MALAT1 and SAA3 has been identified which turns on inflammatory mediators in the endothelium in response to glucose level suggesting a role for MALAT1 in micro- and macro-vascular complications of diabetes [145]. More recently, several studies have identified dysregulation of MALAT1 expression in multiple pathophysiological complications of diabetes including retinopathy, artherosclerosis, cerebrovascular disorder, renal disorders, etc. [144]. Further, molecular studies of several of these pathological indications have converged upon identifying a deregulated inflammatory response induced by altered MALAT1 level. For example, a number of inflammatory molecules such as TNFα and IL6 have been shown to be increased in MALAT1 upregulated cells [147]. Additionally, shRNA mediated the knockdown of MALAT1 ameliorated the inflammatory injury after lung transplant ischemia-reperfusion by inhibiting chemotaxis of neutrophils through p300-mediated downregulation of IL-8 [69]. Further, using Malat1 KO mice it was demonstrated that reduced levels of Malat1 augment atherosclerotic lesion formation in mice and are associated with human atherosclerotic disease [148]. They also showed that pro-atherosclerotic effects observed in Malat1-/- mice were mainly caused by enhanced accumulation of hematopoietic cells involved in inflammatory response [148].
9. Therapeutic Targeting of MALAT1
Given the diverse role of MALAT1 in cancer and other disease areas such as diabetes and inflammation, MALAT1 is being actively investigated as a potential therapeutic target using different modalities. Pre-clinical studies using breast and lung cancer models targeting Malat1 using antisense Gapmer oligonucleotides have resulted in an anti-tumor and anti-metastatic outcome in both studies [63,101]. Gapmer oligonucleotides are short single-stranded RNA-DNA-RNA hybrids that bind to complementary RNA sequences and cause degradation by invoking an RNaseH response [149]. Gapmers are emerging as a promising approach to target multiple lncRNAs [62]. In addition, targeting the Malat1 and AR-v7 axis using Malat1-short interfering RNAs (siRNAs) in enzalutamide-resistant prostate cancer cell lines and mouse models suppressed enzalutamide-resistant prostate cancer progression [113]. Similar studies using Malat1-targeting siRNAs have been conducted in other cancer types, such as glioblastoma, ovarian, colorectal (CRC), gallbladder, gastric, osteosarcoma, and esophageal, etc. [88,106,107,140,142,150,151,152]. MALAT1 gapmer oligonucleotides conjugated to single- walled carbon nanotubes delivered systemically into mice resulted in significant inhibition of multiple myeloma growth [153]. Further, small molecules specifically targeting the MALAT1 triple helix structure have been identified and they lay the foundation for new classes of anticancer therapeutics for the treatment and investigation of MALAT1-driven cancers [154]. Together, these studies provide compelling evidence for targeting MALAT1 in multiple cancer types to achieve a therapeutic benefit. Given that Malat1 knockout mice are healthy and fertile, MALAT1 targeting in cancer can be a potentially viable mechanism to evade the emergence of a drug resistant phenotype in MALAT1 elevated chemo-resistant cancers or to achieve a significant anti-tumor and anti-metastatic effect in MALAT1 overexpressing cancers without causing any adverse side effects to healthy tissues. Based upon the significant body of pre-clinical data MALAT1 is poised to be targeted by antisense or small molecule drugs to impact cancer progression and other inflammatory and metabolic disease indications.
10. Summary and Conclusions
MALAT1 breaks all of the “rules” when it comes to a lncRNA: it is highly abundant, well-conserved, is expressed broadly among different cell types and tissues, and exhibits an unusual 3′-end processing mechanism. As discussed in this review MALAT1 appears to function in a context-dependent manner and as such has been implicated in a wide array of functions. Its expression level has been shown to be altered in many different physiologic states including being upregulated in a plethora of different cancer types, as well as exhibiting altered expression in many other diseases. One intriguing hypothesis derived from these studies is that MALAT1 functions in a context-dependent manner, at the level of pathways rather than individual gene(s), and as such may represent an outstanding therapeutic target as it may impact multiple nodes of particular pathways thereby minimizing the drug resistance problem in cancer treatment. Future studies will certainly add more to the intriguing basic biology of MALAT1 and bring it closer to having clinical impact.
Author Contributions
G.A. and D.A. wrote the manuscript. D.L.S. made suggestions and edited the manuscript and figures. D.A. prepared the figures. All authors have read and agreed to the published version of the manuscript.
Funding
Research in the Spector lab is funded by the National Cancer institute - NCI 5P01CA013106 and the National Institute of General Medical Sciences - NIGMS 1R35GM131833.
Conflicts of Interest
D.L.S. is a consultant to, and receives research support from, Ionis Pharmaceuticals.
References
- Frankish, A.; Diekhans, M.; Ferreira, A.-M.; Johnson, R.; Jungreis, I.; Loveland, J.; Mudge, J.M.; Sisu, C.; Wright, J.; Armstrong, J.; et al. GENCODE reference annotation for the human and mouse genomes. Nucleic Acids Res. 2019, 47, 766–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Schorderet, P.; Duboule, D. Structural and functional differences in the long non-coding RNA hotair in mouse and human. PLoS Genet. 2011, 7, e1002071. [Google Scholar] [CrossRef] [PubMed]
- Rivas, E.; Clements, J.; Eddy, S.R. A statistical test for conserved RNA structure shows lack of evidence for structure in lncRNAs. Nat. Methods 2016, 14, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Diederichs, S. The four dimensions of noncoding RNA conservation. Trends Genet. 2014, 30, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.-C.; Diermeier, S.D.; Yu, A.T.; Brine, L.D.; Bhatia, S.; Alsudani, H.; Kostroff, K.; Bhuiya, T. MaTAR25 LncRNA Regulates the Tensin1 Gene to Impact Breast Cancer Progression. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef]
- Ulitsky, I.; Bartel, D.P. lincRNAs: Genomics, Evolution, and Mechanisms. Cell 2013, 154, 26–46. [Google Scholar] [CrossRef] [Green Version]
- Prasanth, K.V.; Spector, D.L. Eukaryotic regulatory RNAs: An answer to the “genome complexity” conundrum. Genes Dev. 2007, 21, 11–42. [Google Scholar] [CrossRef] [Green Version]
- Fox, A.H.; Nakagawa, S.; Hirose, T.; Bond, C.S. Paraspeckles: Where Long Noncoding RNA Meets Phase Separation. Trends Biochem. Sci. 2018, 43, 124–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, T.; Souquere, S.; Chujo, T.; Kobelke, S.; Chong, Y.S.; Fox, A.H.; Bond, C.S.; Nakagawa, S.; Pierron, G.; Hirose, T. Functional Domains of NEAT1 Architectural lncRNA Induce Paraspeckle Assembly through Phase Separation. Mol. Cell 2018, 70, 1038–1053.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, K.; Dobersch, S.; Barreto, G. Functional interactions between scaffold proteins, noncoding RNAs, and genome loci induce liquid-liquid phase separation as organizing principle for 3-dimensional nuclear architecture: Implications in cancer. FASEB J. 2019, 33, 5814–5822. [Google Scholar] [CrossRef]
- Fay, M.M.; Anderson, P.J. The Role of RNA in Biological Phase Separations. J. Mol. Biol. 2018, 430, 4685–4701. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Genome Regulation by Long Noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchinson, J.N.; Ensminger, A.W.; Clemson, C.M.; Lynch, C.R.; Lawrence, J.B.; Chess, A. A screen for nuclear transcripts identifies two linked noncoding RNAs associated with SC35 splicing domains. BMC Genom. 2007, 8, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, P.; Diederichs, S.; Wang, W.; Böing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin β4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [Green Version]
- Johnsson, P.; Lipovich, L.; Grandér, D.; Morris, K.V. Evolutionary conservation of long non-coding RNAs; sequence, structure, function. Biochim. Biophys. Acta 2014, 1840, 1063–1071. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Müller, F.; Sanges, R. Examples of sequence conservation analyses capture a subset of mouse long non-coding RNAs sharing homology with fish conserved genomic elements. BMC Bioinform. 2013, 14, S14. [Google Scholar] [CrossRef] [Green Version]
- Marques, A.C.; Ponting, C.P. Catalogues of mammalian long noncoding RNAs: Modest conservation and incompleteness. Genome Biol. 2009, 10, R124. [Google Scholar] [CrossRef] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, S.R. Computational Analysis of Conserved RNA Secondary Structure in Transcriptomes and Genomes. Annu. Rev. Biophys. 2014, 43, 433–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.-J.; Yang, D.-C.; Kong, L.; Hou, M.; Meng, Y.-Q.; Wei, L.; Gao, G. CPC2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, 12–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.-P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, 74. [Google Scholar] [CrossRef]
- Aguet, F.; Brown, A.A.; Castel, S.E.; Davis, J.R.; He, Y.; Jo, B.; Mohammadi, P.; Park, Y.S.; Parsana, P.; Segrè, A.V.; et al. Genetic effects on gene expression across human tissues. Nature 2017, 550, 204–213. [Google Scholar]
- Bernard, D.; Prasanth, K.V.; Tripathi, V.; Colasse, S.; Nakamura, T.; Xuan, Z.; Zhang, M.Q.; Sedel, F.; Jourdren, L.; Coulpier, F.; et al. A long nuclear-retained non-coding RNA regulates synaptogenesis by modulating gene expression. EMBO J. 2010, 29, 3082–3093. [Google Scholar] [CrossRef] [Green Version]
- Wilusz, J.E.; Freier, S.M.; Spector, D.L. 3′ End Processing of a Long Nuclear-Retained Noncoding RNA Yields a tRNA-like Cytoplasmic RNA. Cell 2008, 135, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Meseure, D.; Vacher, S.; Lallemand, F.; Alsibai, K.D.; Hatem, R.; Chemlali, W.; Nicolas, A.; De Koning, L.; Pasmant, E.; Callens, C.; et al. Prognostic value of a newly identified MALAT1 alternatively spliced transcript in breast cancer. Br. J. Cancer 2016, 114, 1395–1404. [Google Scholar] [CrossRef] [Green Version]
- Zong, X.; Nakagawa, S.; Freier, S.M.; Fei, J.; Ha, T.; Prasanth, S.G.; Prasanth, K.V. Natural antisense RNA promotes 3′ end processing and maturation of MALAT1 lncRNA. Nucleic Acids Res. 2016, 44, 2898–2908. [Google Scholar] [CrossRef] [Green Version]
- Gomes, C.P.; Nóbrega-Pereira, S.; Domingues-Silva, B.; Rebelo, K.; Alves-Vale, C.; Marinho, S.P.; Carvalho, T.; Dias, S.; Bernardes de Jesus, B. An antisense transcript mediates MALAT1 response in human breast cancer. BMC Cancer 2019, 19, 771. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.A.; Bulkley, D.; Wang, J.; Valenstein, M.L.; Yario, T.A.; Steitz, T.A.; Steitz, J.A. Structural insights into the stabilization of MALAT1 noncoding RNA by a bipartite triple helix. Nat. Struct. Mol. Biol. 2014, 21, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Mao, Y.S.; Diermeier, S.D.; Novikova, I.V.; Nawrocki, E.P.; Jones, T.A.; Lazar, Z.; Tung, C.S.; Luo, W.; Eddy, S.R.; et al. Identification and Characterization of a Class of MALAT1-like Genomic Loci. Cell Rep. 2017, 19, 1723–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilusz, J.E.; JnBaptiste, C.K.; Lu, L.Y.; Kuhn, C.D.; Joshua-Tor, L.; Sharp, P.A. A triple helix stabilizes the 3′ ends of long noncoding RNAs that lack poly(A) tails. Genes Dev. 2012, 26, 2392–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.A.; Valenstein, M.L.; Yario, T.A.; Tycowski, K.T.; Steitz, J.A. Formation of triple-helical structures by the 3′-end sequences of MALAT1 and MENβ noncoding RNAs. Proc. Natl. Acad. Sci. USA 2012, 109, 19202–19207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrad, N.K.; Mili, S.; Marshall, E.L.; Shu, M.D.; Steitz, J.A. Identification of a Rapid Mammalian Deadenylation-Dependent Decay Pathway and Its Inhibition by a Viral RNA Element. Mol. Cell 2006, 24, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Conrad, N.K.; Steitz, J.A. A Kaposi’s sarcoma virus RNA element that increases the nuclear abundance of intronless transcripts. EMBO J. 2005, 24, 1831–1841. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.A.; Kinzig, C.G.; Degregorio, S.J.; Steitz, J.A. Methyltransferase-like protein 16 binds the 3’-terminal triple helix of MALAT1 long noncoding RNA. Proc. Natl. Acad. Sci. USA 2016, 113, 14013–14018. [Google Scholar] [CrossRef] [Green Version]
- Ageeli, A.A.; McGovern-Gooch, K.R.; Kaminska, M.M.; Baird, N.J. Finely tuned conformational dynamics regulate the protective function of the lncRNA MALAT1 triple helix. Nucleic Acids Res. 2019, 47, 1468–1481. [Google Scholar] [CrossRef] [Green Version]
- Carlile, T.M.; Rojas-Duran, M.F.; Zinshteyn, B.; Shin, H.; Bartoli, K.M.; Gilbert, W.V. Pseudouridine profiling reveals regulated mRNA pseudouridylation in yeast and human cells. Nature 2014, 515, 143–146. [Google Scholar] [CrossRef] [Green Version]
- Squires, J.E.; Patel, H.R.; Nousch, M.; Sibbritt, T.; Humphreys, D.T.; Parker, B.J.; Suter, C.M.; Preiss, T. Widespread occurrence of 5-methylcytosine in human coding and non-coding RNA. Nucleic Acids Res. 2012, 40, 5023–5033. [Google Scholar] [CrossRef]
- Li, X.; Zhu, P.; Ma, S.; Song, J.; Bai, J.; Sun, F.; Yi, C. Chemical pulldown reveals dynamic pseudouridylation of the mammalian transcriptome. Nat. Chem. Biol. 2015, 11, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N6-methyladenosine-dependent RNA structural switches regulate RNA–protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spector, D.L.; Lamond, A.I. Nuclear speckles. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, J.; Jadaliha, M.; Harmon, T.S.; Li, I.T.S.; Hua, B.; Hao, Q.; Holehouse, A.S.; Reyer, M.; Sun, Q.; Freier, S.M.; et al. Quantitative analysis of multilayer organization of proteins and RNA in nuclear speckles at super resolution. J. Cell Sci. 2017, 130, 4180–4192. [Google Scholar] [CrossRef] [Green Version]
- Miyagawa, R.; Tano, K.; Mizuno, R.; Nakamura, Y.; Ijiri, K.; Rakwal, R.; Shibato, J.; Masuo, Y.; Mayeda, A.; Hirose, T.; et al. Identification of cis-and trans-acting factors involved in the localization of MALAT-1 noncoding RNA to nuclear speckles. RNA 2012, 18, 738–751. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Arun, G.; Mao, Y.S.; Lazar, Z.; Hung, G.; Bhattacharjee, G.; Xiao, X.; Booth, C.J.; Wu, J.; Zhang, C.; et al. The lncRNA Malat1 is dispensable for mouse development but its transcription plays a cis-regulatory role in the adult. Cell Rep. 2012, 2, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Yi, F.; Han, X.; Du, Q.; Liang, Z. MALAT-1 interacts with hnRNP C in cell cycle regulation. FEBS Lett. 2013, 587, 3175–3181. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Liu, Y.; Zhuang, H.; Yang, B.; Hei, K.; Xiao, M.; Hou, C.; Gao, H.; Zhang, X.; Jia, C.; et al. Quantitative proteomics reveals that long non-coding RNA MALAT1 interacts with DBC1 to regulate p53 acetylation. Nucleic Acids Res. 2017, 45, 9947–9959. [Google Scholar] [CrossRef]
- Mao, Y.S.; Zhang, B.; Spector, D.L. Biogenesis and function of nuclear bodies. Trends Genet. 2011, 27, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Lu, T.; Emanuel, G.; Babcock, H.P.; Zhuang, X. Imaging-based pooled CRISPR screening reveals regulators of lncRNA localization. Proc. Natl. Acad. Sci. USA 2019, 166, 10842–10851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.M.; Kabotyanski, E.B.; Reineke, L.C.; Shao, J.; Xiong, F.; Lee, J.-H.; Dubrulle, J.; Johnson, H.; Stossi, F.; Tsoi, P.S.; et al. The SINEB1 element in the long non-coding RNA Malat1 is necessary for TDP-43 proteostasis. Nucleic Acids Res. 2020, 48, 2621–2642. [Google Scholar] [CrossRef] [PubMed]
- West, J.A.; Davis, C.P.; Sunwoo, H.; Simon, M.D.; Sadreyev, R.I.; Wang, P.I.; Tolstorukov, M.Y.; Kingston, R.E. The Long Noncoding RNAs NEAT1 and MALAT1 Bind Active Chromatin Sites. Mol. Cell 2014, 55, 791–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, C.; Qu, K.; Zhong, F.L.; Artandi, S.E.; Chang, H.Y. Genomic Maps of Long Noncoding RNA Occupancy Reveal Principles of RNA-Chromatin Interactions. Mol. Cell 2011, 44, 667–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Lin, C.; Liu, W.; Zhang, J.; Ohgi, K.A.; Grinstein, J.D.; Dorrestein, P.C.; Rosenfeld, M.G. ncRNA- and Pc2 Methylation-Dependent Gene Relocation between Nuclear Structures Mediates Gene Activation Programs. Cell 2011, 147, 773–788. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Ooi, H.S.; Wu, J.; Chen, J.; Zhang, X.; Tan, S.; Yu, Q.; Li, Y.Y.; Kang, Y.; Li, H.; et al. MALAT1 long ncRNA promotes gastric cancer metastasis by suppressing PCDH10. Oncotarget 2016, 7, 12693–12703. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Ding, L.; Wang, L.; Zhao, Y.; Sun, Z.; Karnes, R.J.; Zhang, J.; Huang, H. LncRNA MALAT1 enhances oncogenic activities of EZH2 in castration-resistant prostate cancer. Oncotarget 2015, 6, 41045–41055. [Google Scholar] [CrossRef]
- Cai, L.; Chang, H.; Fang, Y.; Li, G. A Comprehensive Characterization of the Function of LincRNAs in Transcriptional Regulation Through Long-Range Chromatin Interactions. Sci. Rep. 2016, 6, 36572. [Google Scholar] [CrossRef]
- Li, B.; Chen, P.; Qu, J.; Shi, L.; Zhuang, W.; Fu, J.; Li, J.; Zhang, X.; Sun, Y.; Zhuang, W. Activation of LTBP3 Gene by a Long Noncoding RNA (lncRNA) MALAT1 Transcript in Mesenchymal Stem Cells from Multiple Myeloma. J. Biol. Chem. 2014, 289, 29365–29375. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, B.; Rivas-Astroza, M.; Nguyen, T.C.; Chen, W.; Yan, Z.; Cao, X.; Hebert, L.; Zhong, S. Systematic Mapping of RNA-Chromatin Interactions In Vivo. Curr. Biol. 2017, 27, 602–609. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhou, B.; Chen, L.; Gou, L.T.; Li, H.; Fu, X.D. GRID-seq reveals the global RNA-chromatin interactome. Nat. Biotechnol. 2017, 35, 940–950. [Google Scholar] [CrossRef] [PubMed]
- Arun, G.; Diermeier, S.D.; Spector, D.L. Therapeutic Targeting of Long Non-Coding RNAs in Cancer. Trends Mol. Med. 2018, 24, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Arun, G.; Diermeier, S.; Akerman, M.; Chang, K.-C.; Wilkinson, J.E.; Hearn, S.; Kim, Y.; MacLeod, A.R.; Krainer, A.R.; Norton, L.; et al. Differentiation of mammary tumors and reduction in metastasis upon Malat1 lncRNA loss. Genes Dev. 2016, 30, 34–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malakar, P.; Shilo, A.; Mogilevsky, A.; Stein, I.; Pikarsky, E.; Nevo, Y.; Benyamini, H.; Elgavish, S.; Zong, X.; Prasanth, K.V.; et al. Long noncoding RNA MALAT1 promotes hepatocellular carcinoma development by SRSF1 upregulation and mTOR activation. Cancer Res. 2017, 77, 1155–1167. [Google Scholar] [CrossRef] [Green Version]
- Engreitz, J.M.; Sirokman, K.; McDonel, P.; Shishkin, A.A.; Surka, C.; Russell, P.; Grossman, S.R.; Chow, A.Y.; Guttman, M.; Lander, E.S. RNA-RNA interactions enable specific targeting of noncoding RNAs to nascent pre-mRNAs and chromatin sites. Cell 2014, 159, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Polymenidou, M.; Lagier-Tourenne, C.; Hutt, K.R.; Huelga, S.C.; Moran, J.; Liang, T.Y.; Ling, S.; Sun, E.; Wancewicz, E.; Mazur, C.; et al. Long pre-mRNA depletion and RNA missplicing contribute to neuronal vulnerability from loss of TDP-43. Nat. Neurosci. 2011, 14, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Änkö, M.-L.; Müller-McNicoll, M.; Brandl, H.; Curk, T.; Gorup, C.; Henry, I.; Ule, J.; Neugebauer, K.M. The RNA-binding landscapes of two SR proteins reveal unique functions and binding to diverse RNA classes. Genome Biol. 2012, 13, R17. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Xu, D.; Sha, W.; Shen, L.; Lu, G. Long non-coding RNA MALAT1 interacts with transcription factor Foxo1 to regulate SIRT1 transcription in high glucose-induced HK-2 cells injury. Biochem. Biophys. Res. Commun. 2018, 503, 849–855. [Google Scholar] [CrossRef]
- Wei, L.; Li, J.; Han, Z.; Chen, Z.; Zhang, Q. Silencing of lncRNA MALAT1 Prevents Inflammatory Injury after Lung Transplant Ischemia-Reperfusion by Downregulation of IL-8 via p300. Mol. Ther. Nucleic Acids 2019, 18, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Chen, N.; Zhou, L.; Wang, C.; Wen, X.; Jia, L.; Cui, J.; Hoffman, A.R.; Hu, J.-F.; Li, W. Genome-wide target interactome profiling reveals a novel EEF1A1 epigenetic pathway for oncogenic lncRNA MALAT1 in breast cancer. Am. J. Cancer Res. 2019, 9, 714–729. [Google Scholar]
- Amodio, N.; Stamato, M.A.; Juli, G.; Morelli, E.; Fulciniti, M.; Manzoni, M.; Taiana, E.; Agnelli, L.; Cantafio, M.E.G.; Romeo, E.; et al. Drugging the lncRNA MALAT1 via LNA gapmeR ASO inhibits gene expression of proteasome subunits and triggers anti-multiple myeloma activity. Leukemia 2018, 32, 1948–1957. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Dang, Y.; Wei, D.; Chen, P.; Tang, R.; Huang, Q.; Liu, J.; Luo, D. Clinical significance and prospective molecular mechanism of MALAT1 in pancreatic cancer exploration: A comprehensive study based on the GeneChip, GEO, Oncomine, and TCGA databases. Oncol. Targets Ther. 2017, 10, 3991–4005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalik, K.M.; You, X.; Manavski, Y.; Doddaballapur, A.; Zörnig, M.; Braun, T.; John, D.; Ponomareva, Y.; Chen, W.; Uchida, S.; et al. Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth. Circ. Res. 2014, 114, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Piao, H.-L.; Kim, B.-J.; Yao, F.; Han, Z.; Wang, Y.; Xiao, Z.; Siverly, A.N.; Lawhon, S.E.; Ton, B.N.; et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat. Genet. 2018, 50, 1705–1715. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.; Shen, Z.; Chakraborty, A.; Giri, S.; Freier, S.M.; Wu, X.; Zhang, Y.; Gorospe, M.; Prasanth, S.G.; Lal, A.; et al. Long noncoding RNA MALAT1 controls cell cycle progression by regulating the expression of oncogenic transcription factor B-MYB. PLoS Genet. 2013, 9, e1003368. [Google Scholar] [CrossRef] [PubMed]
- Herzel, L.; Ottoz, D.S.M.; Alpert, T.; Neugebauer, K.M. Splicing and transcription touch base: Co-transcriptional spliceosome assembly and function. Nat. Rev. Mol. Cell Biol. 2017, 18, 637–650. [Google Scholar] [CrossRef]
- Nojima, T.; Rebelo, K.; Gomes, T.; Grosso, A.R.; Proudfoot, N.J.; Carmo-Fonseca, M. RNA Polymerase II Phosphorylated on CTD Serine 5 Interacts with the Spliceosome during Co-transcriptional Splicing. Mol. Cell 2018, 72, 369–379.e4. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Liu, Y.; Zhang, H.; Wang, T.; Diao, R.; Jiang, Z.; Gui, Y.; Cai, Z. Hsa-miR-125b suppresses bladder cancer development by down-regulating oncogene SIRT7 and oncogenic long non-coding RNA MALAT1. FEBS Lett. 2013, 587, 3875–3882. [Google Scholar] [CrossRef]
- Yu, F.; Lu, Z.; Cai, J.; Huang, K.; Chen, B.; Li, G.; Dong, P.; Zheng, J. Malat1 functions as a competing endogenous RNA to mediate Rac1 expression by sequestering miR-101b in liver fibrosis. Cell Cycle 2015, 14, 3885–3896. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; He, H.; Xiao, W.; Liu, Q.; Deng, Z.; Lu, Y.; Wang, Q.; Zheng, Q.; Li, Y. MALAT1 promotes osteosarcoma development by targeting TGFA via MIR376A. Oncotarget 2016, 7, 54733–54743. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Zhang, Z.; Xiong, W.; Li, N.; Liu, H.; He, H.; Li, Q.; Liu, Y.; Zhang, L. Estradiol promotes EMT in endometriosis via MALAT1/ miR200s sponge function. Reproduction 2019, 157, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.; Tang, K.; Liu, P.; Chen, K.; Hu, J.; Zeng, J.; Xiao, W.; Yu, G.; Yao, W.; Zhou, H.; et al. LncRNA MALAT1 functions as a competing endogenous RNA to regulate ZEB2 expression by sponging miR-200s in clear cell kidney carcinoma. Oncotarget 2015, 6, 38005–38015. [Google Scholar] [CrossRef] [PubMed]
- Eißmann, M.; Gutschner, T.; Hämmerle, M.; Günther, S.; Caudron-Herger, M.; Groß, M.; Schirmacher, P.; Rippe, K.; Braun, T.; Zörnig, M.; et al. Loss of the abundant nuclear non-coding RNA MALAT1 is compatible with life and development. RNA Biol. 2012, 9, 1076–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, S.; Ip, J.Y.; Shioi, G.; Tripathi, V.; Zong, X.; Hirose, T.; Prasanth, K.V. Malat1 is not an essential component of nuclear speckles in mice. RNA 2012, 18, 1487–1499. [Google Scholar] [CrossRef] [Green Version]
- Barbaric, I.; Miller, G.; Dear, T.N. Appearances can be deceiving: Phenotypes of knockout mice. Brief. Funct. Genom. Proteom. 2007, 6, 91–103. [Google Scholar] [CrossRef] [Green Version]
- El-Brolosy, M.A.; Stainier, D.Y.R. Genetic compensation: A phenomenon in search of mechanisms. PLoS Genet. 2017, 13, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Liu, Y.; Li, Y.; Li, G. Inhibition of ADP-ribosylation factor-like 6 interacting protein 1 suppresses proliferation and reduces tumor cell invasion in CaSki human cervical cancer cells. Mol. Biol. Rep. 2010, 37, 3819–3825. [Google Scholar] [CrossRef]
- Wu, X.-S.; Wang, X.-A.; Wu, W.-G.; Hu, Y.-P.; Li, M.-L.; Ding, Q.; Weng, H.; Shu, Y.-J.; Liu, T.-Y.; Jiang, L.; et al. MALAT1 promotes the proliferation and metastasis of gallbladder cancer cells by activating the ERK/MAPK pathway. Cancer Biol. Ther. 2014, 15, 806–814. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, M.; Wang, Z.; Han, S.; Tang, X.; Ge, Y.; Zhou, L.; Zhou, C.; Yuan, Q.; Yang, M. Silencing of long noncoding RNA MALAT1 by miR-101 and miR-217 inhibits proliferation, migration, and invasion of esophageal squamous cell carcinoma cells. J. Biol. Chem. 2015, 290, 3925–3935. [Google Scholar] [CrossRef] [Green Version]
- Kan, J.Y.; Wu, D.C.; Yu, F.J.; Wu, C.Y.; Ho, Y.W.; Chiu, Y.J.; Jian, S.F.; Hung, J.Y.; Wang, J.Y.; Kuo, P.L. Chemokine (C-C Motif) ligand 5 is involved in tumor-associated dendritic cell-mediated colon cancer progression through non-coding RNA MALAT-1. J. Cell. Physiol. 2015, 230, 1883–1894. [Google Scholar] [CrossRef]
- Ying, L.; Chen, Q.; Wang, Y.; Zhou, Z.; Huang, Y.; Qiu, F. Upregulated MALAT-1 contributes to bladder cancer cell migration by inducing epithelial-to-mesenchymal transition. Mol. Biosyst. 2012, 8, 2289–2294. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Chen, L.; Wang, Y.; Jiang, X.; Xia, H.; Zhuang, Z. Long noncoding RNA MALAT1 promotes brain metastasis by inducing epithelial-mesenchymal transition in lung cancer. J. Neurooncol. 2015, 121, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Katsaros, D.; Biglia, N.; Shen, Y.; Fu, Y.; Loo, L.W.M.; Jia, W.; Obata, Y.; Yu, H. High expression of long non-coding RNA MALAT1 in breast cancer is associated with poor relapse-free survival. Breast Cancer Res. Treat. 2018, 171, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Tano, K.; Mizuno, R.; Okada, T.; Rakwal, R.; Shibato, J.; Masuo, Y.; Ijiri, K.; Akimitsu, N. MALAT-1 enhances cell motility of lung adenocarcinoma cells by influencing the expression of motility-related genes. FEBS Lett. 2010, 584, 4575–4580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amodio, N.; Raimondi, L.; Juli, G.; Stamato, M.A.; Caracciolo, D.; Tagliaferri, P.; Tassone, P. MALAT1: A druggable long non-coding RNA for targeted anti-cancer approaches. J. Hematol. Oncol. 2018, 11, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ronchetti, D.; Agnelli, L.; Taiana, E.; Galletti, S.; Manzoni, M.; Todoerti, K.; Musto, P.; Strozzi, F.; Neri, A. Distinct lncRNA transcriptional fingerprints characterize progressive stages of multiple myeloma. Oncotarget 2016, 7, 14814–14830. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Chen, H.; Gao, Y.; Wang, Y.W.; Zhang, G.Q.; Pan, S.H.; Ji, L.; Kong, R.; Wang, G.; Jia, Y.H.; et al. Long noncoding RNA MALAT1 promotes aggressive pancreatic cancer proliferation and metastasis via the stimulation of autophagy. Mol. Cancer Ther. 2016, 15, 2232–2243. [Google Scholar] [CrossRef] [Green Version]
- Okugawa, Y.; Toiyama, Y.; Hur, K.; Toden, S.; Saigusa, S.; Tanaka, K.; Inoue, Y.; Mohri, Y.; Kusunoki, M.; Richard Boland, C.; et al. Metastasis-associated long non-coding RNA drives gastric cancer development and promotes peritoneal metastasis. Carcinogenesis 2014, 35, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Tee, A.E.; Ling, D.; Nelson, C.; Atmadibrata, B.; Dinger, M.E.; Xu, N.; Mizukami, T.; Liu, P.Y.; Liu, B.; Cheung, B.; et al. The histone demethylase JMJD1A induces cell migration and invasion by up-regulating the expression of the long noncoding RNA MALAT1. Oncotarget 2014, 5, 1793–1804. [Google Scholar] [CrossRef] [Green Version]
- Guffanti, A.; Iacono, M.; Pelucchi, P.; Kim, N.; Soldà, G.; Croft, L.J.; Taft, R.J.; Rizzi, E.; Askarian-Amiri, M.; Bonnal, R.J.; et al. A transcriptional sketch of a primary human breast cancer by 454 deep sequencing. BMC Genom. 2009, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Gutschner, T.; Hammerle, M.; Eissmann, M.; Hsu, J.; Kim, Y.; Hung, G.; Revenko, A.; Arun, G.; Stentrup, M.; Gross, M.; et al. The Noncoding RNA MALAT1 Is a Critical Regulator of the Metastasis Phenotype of Lung Cancer Cells. Cancer Res. 2013, 73, 1180–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadaliha, M.; Zong, X.; Malakar, P.; Ray, T.; Singh, D.K.; Freier, S.M.; Jensen, T.; Prasanth, S.G.; Karni, R.; Ray, P.S.; et al. Functional and prognostic significance of long non-coding RNA MALAT1 as a metastasis driver in ER negative lymph node negative breast cancer. Oncotarget 2016, 7, 40418–40436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, L.H.; Spieker, T.; Koschmieder, S.; Humberg, J.; Jungen, D.; Bulk, E.; Hascher, A.; Wittmer, D.; Marra, A.; Hillejan, L.; et al. The long noncoding MALAT-1 RNA indicates a poor prognosis in non-small cell lung cancer and induces migration and tumor growth. J. Thorac. Oncol. 2011, 6, 1984–1992. [Google Scholar] [CrossRef] [Green Version]
- Voce, D.J.; Bernal, G.M.; Wu, L.; Crawley, C.D.; Zhang, W.; Mansour, N.M.; Cahill, K.E.; Szymura, S.J.; Uppal, A.; Raleigh, D.R.; et al. Temozolomide treatment induces lncRNA MALAT1 in an NF-κB and p53 codependent manner in Glioblastoma. Cancer Res. 2019, 79, 2536–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ching, T.; Peplowska, K.; Huang, S.; Zhu, X.; Shen, Y.; Molnar, J.; Yu, H.; Tiirikainen, M.; Fogelgren, B.; Fan, R.; et al. Pan-Cancer Analyses Reveal Long Intergenic Non-Coding RNAs Relevant to Tumor Diagnosis, Subtyping and Prognosis. EBioMedicine 2016, 7, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Wu, Y.; Tan, D.; Meng, H.; Wang, K.; Bai, Y.; Yang, K. Up-regulation of long noncoding RNA MALAT1 contributes to proliferation and metastasis in esophageal squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2015, 34, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhang, X.; Hu, X.; Zhou, W.; Zhang, P.; Zhang, J.; Yang, S.; Liu, Y. The effects of lncRNA MALAT1 on proliferation, invasion and migration in colorectal cancer through regulating SOX9. Mol. Med. 2018, 24, 1–15. [Google Scholar] [CrossRef]
- Yoshimoto, R.; Mayeda, A.; Yoshida, M.; Nakagawa, S. MALAT1 long non-coding RNA in cancer. Biochim. Biophys. Acta Gene Regul. Mech. 2016, 1859, 192–199. [Google Scholar] [CrossRef]
- Ren, S.; Liu, Y.; Xu, W.; Sun, Y.; Lu, J.; Wang, F.; Wei, M.; Shen, J.; Hou, J.; Gao, X.; et al. Long noncoding RNA MALAT-1 is a new potential therapeutic target for castration resistant prostate cancer. J. Urol. 2013, 190, 2278–2287. [Google Scholar] [CrossRef]
- Li, L.; Feng, T.; Lian, Y.; Zhang, G.; Garen, A.; Song, X. Role of human noncoding RNAs in the control of tumorigenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 12956–12961. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Chakraborty, G.; Lee-Lim, A.P.; Mavrakis, K.J.; Wendel, H.G.; Giancotti, F.G. Forward genetic screens in mice uncover mediators and suppressors of metastatic reactivation. Proc. Natl. Acad. Sci. USA 2014, 111, 16532–16537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Liu, X.; Xu, Y.; Zhang, K.; Huang, J.; Pan, B.; Chen, D.; Cui, S.; Song, H.; Wang, R.; et al. TFAP2C-Activated MALAT1 Modulates the Chemoresistance of Docetaxel-Resistant Lung Adenocarcinoma Cells. Mol. Ther. Nucleic Acids 2019, 14, 567–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Sun, Y.; Li, L.; Niu, Y.; Lin, W.; Lin, C.; Antonarakis, E.S.; Luo, J.; Yeh, S.; Chang, C. Preclinical Study using Malat1 Small Interfering RNA or Androgen Receptor Splicing Variant 7 Degradation Enhancer ASC-J9® to Suppress Enzalutamide-resistant Prostate Cancer Progression. Eur. Urol. 2017, 72, 835–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abildgaard, C.; Do Canto, L.M.; Steffensen, K.D.; Rogatto, S.R. Long Non-coding RNAs Involved in Resistance to Chemotherapy in Ovarian Cancer. Front. Oncol. 2020, 9, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwok, Z.H.; Roche, V.; Chew, X.H.; Fadieieva, A.; Tay, Y. A non-canonical tumor suppressive role for the long non-coding RNA MALAT1 in colon and breast cancers. Int. J. Cancer 2018, 143, 668–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastlack, S.C.; Dong, S.; Mo, Y.Y.; Alahari, S.K. Expression of long noncoding RNA MALAT1 correlates with increased levels of Nischarin and inhibits oncogenic cell functions in breast cancer. PLoS ONE 2018, 13, e0198945. [Google Scholar] [CrossRef]
- Arun, G.; Spector, D.L. MALAT1 long non-coding RNA and breast cancer. RNA Biol. 2019, 16, 860–863. [Google Scholar] [CrossRef]
- Davis, I.J.; Hsi, B.-L.; Arroyo, J.D.; Vargas, S.O.; Yeh, Y.A.; Motyckova, G.; Valencia, P.; Perez-Atayde, A.R.; Argani, P.; Ladanyi, M.; et al. Cloning of an Alpha-TFEB fusion in renal tumors harboring the t(6;11)(p21;q13) chromosome translocation. Proc. Natl. Acad. Sci. USA 2003, 100, 6051–6056. [Google Scholar] [CrossRef] [Green Version]
- Graham, R.P.; Nair, A.A.; Davila, J.I.; Jin, L.; Jen, J.; Sukov, W.R.; Wu, T.T.; Appelman, H.D.; Torres-Mora, J.; Perry, K.D.; et al. Gastroblastoma harbors a recurrent somatic MALAT1-GLI1 fusion gene. Mod. Pathol. 2017, 30, 1443–1452. [Google Scholar] [CrossRef]
- Menghi, F.; Barthel, F.P.; Yadav, V.; Tang, M.; Ji, B.; Tang, Z.; Carter, G.W.; Ruan, Y.; Scully, R.; Verhaak, R.G.W.; et al. The Tandem Duplicator Phenotype Is a Prevalent Genome-Wide Cancer Configuration Driven by Distinct Gene Mutations. Cancer Cell 2018, 34, 197–210.e5. [Google Scholar] [CrossRef]
- Nik-Zainal, S.; Davies, H.; Staaf, J.; Ramakrishna, M.; Glodzik, D.; Zou, X.; Martincorena, I.; Alexandrov, L.B.; Martin, S.; Wedge, D.C.; et al. Landscape of somatic mutations in 560 breast cancer whole-genome sequences. Nature 2016, 534, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.J.; Ding, L.; Shen, D.; Luo, J.; Suman, V.J.; Wallis, J.W.; Van Tine, B.A.; Hoog, J.; Goiffon, R.J.; Goldstein, T.C.; et al. Whole-genome analysis informs breast cancer response to aromatase inhibition. Nature 2012, 486, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Rheinbay, E.; Nielsen, M.M.; Abascal, F.; Wala, J.A.; Shapira, O.; Tiao, G.; Hornshøj, H.; Hess, J.M.; Juul, R.I.; Lin, Z.; et al. Analyses of non-coding somatic drivers in 2,658 cancer whole genomes. Nature 2020, 578, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, C.; Davidson, N.R.; Demircioğlu, D.; Fonseca, N.A.; He, Y.; Kahles, A.; Lehmann, K.-V.; Liu, F.; Shiraishi, Y.; Soulette, C.M.; et al. Genomic basis for RNA alterations in cancer. Nature 2020, 578, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Cui, Z.; Li, H.; Lv, X.; Gao, M.; Yang, Z.; Bi, Y.; Zhang, Z.; Wang, S.; Zhou, B.; et al. Clinicopathological and prognostic significance of long noncoding RNA MALAT1 in human cancers: A review and meta-analysis. Cancer Cell Int. 2018, 18, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Xia, Y.; Wang, Z.; Zheng, J.; Chen, Y.; Li, X.; Wang, Y.; Ming, H. Serum long non coding RNA MALAT-1 protected by exosomes is up-regulated and promotes cell proliferation and migration in non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2017, 490, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Su, Y.; He, X.; Zhao, W.; Wu, C.; Zhang, W.; Si, X.; Dong, B.; Zhao, L.; Gao, Y.; et al. Plasma long non-coding RNA MALAT1 is associated with distant metastasis in patients with epithelial ovarian cancer. Oncol. Lett. 2016, 12, 1361–1366. [Google Scholar] [CrossRef]
- Qiu, J.-J.; Lin, X.-J.; Tang, X.-Y.; Zheng, T.-T.; Lin, Y.-Y.; Hua, K.-Q. Exosomal Metastasis-Associated Lung Adenocarcinoma Transcript 1 Promotes Angiogenesis and Predicts Poor Prognosis in Epithelial Ovarian Cancer. Int. J. Biol. Sci. 2018, 14, 1960–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Ren, S.; Chen, R.; Lu, J.; Shi, X.; Zhu, Y.; Zhang, W.; Jing, T.; Zhang, C.; Shen, J.; et al. Development and prospective multicenter evaluation of the long noncoding RNA MALAT-1 as a diagnostic urinary biomarker for prostate cancer. Oncotarget 2014, 5, 11091–11102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Y.; Du, L.; Wang, L.; Jiang, X.; Zhang, S.; Li, J.; Yan, K.; Duan, W.; Zhao, Y.; Wang, L.; et al. Expression signatures of exosomal long non-coding RNAs in urine serve as novel non-invasive biomarkers for diagnosis and recurrence prediction of bladder cancer 11 Medical and Health Sciences 1112 Oncology and Carcinogenesis. Mol. Cancer 2018, 17, 1–5. [Google Scholar]
- Stone, J.K.; Kim, J.H.; Vukadin, L.; Richard, A.; Giannini, H.K.; Lim, S.T.S.; Tan, M.; Ahn, E.Y.E. Hypoxia induces cancer cell-specific chromatin interactions and increases MALAT1 expression in breast cancer cells. J. Biol. Chem. 2019, 294, 11213–11224. [Google Scholar] [CrossRef]
- Malakar, P.; Stein, I.; Saragovi, A.; Winkler, R.; Stern-Ginossar, N.; Berger, M.; Pikarsky, E.; Karni, R. Long Noncoding RNA MALAT1 Regulates Cancer Glucose Metabolism by Enhancing mTOR-Mediated Translation of TCF7L2. Cancer Res. 2019, 79, 2480–2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kölling, M.; Genschel, C.; Kaucsar, T.; Hübner, A.; Rong, S.; Schmitt, R.; Sörensen-Zender, I.; Haddad, G.; Kistler, A.; Seeger, H.; et al. Hypoxia-induced long non-coding RNA Malat1 is dispensable for renal ischemia/reperfusion-injury. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhry, H.; Scho’del, J.; Oikonomopoulos, S.; Camps, C.; Grampp, S.; Harris, A.L.; Ratcliffe, P.J.; Ragoussis, J.; Mole, D.R. Extensive regulation of the non-coding transcriptome by hypoxia: Role of HIF in releasing paused RNApol2. EMBO Rep. 2014, 15, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Sallé-Lefort, S.; Miard, S.; Nolin, M.A.; Boivin, L.; Paré, M.È.; Debigaré, R.; Picard, F. Hypoxia upregulates Malat1 expression through a CaMKK/AMPK/HIF-1α axis. Int. J. Oncol. 2016, 49, 1731–1736. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, S.; Kitadate, A.; Abe, F.; Takahashi, N.; Tagawa, H. Hypoxia-inducible KDM3A addiction in multiple myeloma. Blood Adv. 2018, 2, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Lin, J.; Fang, H.; Fang, J.; Li, C.; Chen, W.; Liu, S.; Ondrejka, S.; Gong, Z.; Reu, F.; et al. Targeting the MALAT1/PARP1/LIG3 complex induces DNA damage and apoptosis in multiple myeloma. Leukemia 2018, 32, 2250–2262. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, H.; Li, Q.; Fu, J.; Huang, J.; Zhao, Y. MALAT1 Activates the P53 signaling pathway by regulating MDM2 to promote ischemic stroke. Cell. Physiol. Biochem. 2018, 50, 2216–2228. [Google Scholar] [CrossRef]
- Handa, H.; Kuroda, Y.; Kimura, K.; Masuda, Y.; Hattori, H.; Alkebsi, L.; Matsumoto, M.; Kasamatsu, T.; Kobayashi, N.; Tahara, K.I.; et al. Long non-coding RNA MALAT1 is an inducible stress response gene associated with extramedullary spread and poor prognosis of multiple myeloma. Br. J. Haematol. 2017, 179, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Cai, T.; Liu, Y.; Xiao, J. Long noncoding RNA MALAT1 knockdown reverses chemoresistance to temozolomide via promoting microRNA-101 in glioblastoma. Cancer Med. 2018, 7, 1404–1415. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Zhang, X.; Wang, H.; Wang, L.; Liu, T.; Du, L.; Yang, Y.; Wang, C. MALAT1 is associated with poor response to oxaliplatin-based chemotherapy in colorectal cancer patients and promotes chemoresistance through EZH2. Mol. Cancer Ther. 2017, 16, 739–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, L.; Wang, A.; Zhang, Y.; Xu, X.; Zhang, X. Knockdown of MALAT1 enhances chemosensitivity of ovarian cancer cells to cisplatin through inhibiting the Notch1 signaling pathway. Exp. Cell Res. 2018, 366, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Xue, D.; Lu, H.; Xu, H.Y.; Zhou, C.X.; He, X.Z. Long noncoding RNA MALAT1 enhances the docetaxel resistance of prostate cancer cells via miR-145-5p-mediated regulation of AKAP12. J. Cell. Mol. Med. 2018, 22, 3223–3237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hamblin, M.H.; Yin, K.J. The long noncoding RNA Malat1: Its physiological and pathophysiological functions. RNA Biol. 2017, 14, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Puthanveetil, P.; Chen, S.; Feng, B.; Gautam, A.; Chakrabarti, S. Long non-coding RNA MALAT1 regulates hyperglycaemia induced inflammatory process in the endothelial cells. J. Cell. Mol. Med. 2015, 19, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ke, S.; Zhong, L.; Wu, J.; Tseng, A.; Morpurgo, B.; Golovko, A.; Wang, G.; Cai, J.J.; Ma, X.; et al. Long noncoding RNA MALAT1 regulates generation of reactive oxygen species and the insulin responses in male mice. Biochem. Pharmacol. 2018, 152, 94–103. [Google Scholar] [CrossRef]
- Biswas, S.; Thomas, A.A.; Chen, S.; Aref-Eshghi, E.; Feng, B.; Gonder, J.; Sadikovic, B.; Chakrabarti, S. MALAT1: An Epigenetic Regulator of Inflammation in Diabetic Retinopathy. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Cremer, S.; Michalik, K.M.; Fischer, A.; Pfisterer, L.; Jaé, N.; Winter, C.; Boon, R.A.; Muhly-Reinholz, M.; John, D.; Uchida, S.; et al. Hematopoietic deficiency of the long noncoding RNA Malat1 promotes atherosclerosis and plaque inflammation. Circulation 2019, 139, 1320–1334. [Google Scholar] [CrossRef]
- Crooke, S.T. Molecular Mechanisms of Antisense Oligonucleotides. Nucleic Acid Ther. 2017, 27, 70–77. [Google Scholar] [CrossRef]
- Sun, Z.; Ou, C.; Liu, J.; Chen, C.; Zhou, Q.; Yang, S.; Li, G.; Wang, G.; Song, J.; Li, Z.; et al. YAP1-induced MALAT1 promotes epithelial–mesenchymal transition and angiogenesis by sponging miR-126-5p in colorectal cancer. Oncogene 2019, 38, 2627–2644. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Liu, Y.; Yang, W.; Xia, Y.; Yang, C.; Yang, S.; Liu, X. Long noncoding RNA MALAT1 as a potential therapeutic target in osteosarcoma. J. Orthop. Res. 2016, 34, 932–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Q.; Xie, L.; Li, H. Overexpressed MALAT1 promotes invasion and metastasis of gastric cancer cells via increasing EGFL7 expression. Life Sci. 2016, 157, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Hu, Y.; Zhao, J.-J. Repression of Multiple Myeloma Cell Growth In Vivo by Single-wall Carbon Nanotube (SWCNT)-delivered MALAT1 Antisense Oligos. J. Vis. Exp. 2018, 176, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Abulwerdi, F.A.; Xu, W.; Ageeli, A.A.; Yonkunas, M.J.; Arun, G.; Nam, H.; Schneekloth, J.S.; Dayie, T.K.; Spector, D.; Baird, N.; et al. Selective Small-Molecule Targeting of a Triple Helix Encoded by the Long Noncoding RNA, MALAT1. ACS Chem. Biol. 2019, 14, 223–235. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Molecular mechanisms of MALAT1 function.

Figure 2.
Summary of known disease implications of MALAT1.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arun, G.; Aggarwal, D.; Spector, D.L. MALAT1 Long Non-Coding RNA: Functional Implications. Non-Coding RNA 2020, 6, 22. https://doi.org/10.3390/ncrna6020022
AMA Style
Arun G, Aggarwal D, Spector DL. MALAT1 Long Non-Coding RNA: Functional Implications. Non-Coding RNA. 2020; 6(2):22. https://doi.org/10.3390/ncrna6020022
Chicago/Turabian StyleArun, Gayatri, Disha Aggarwal, and David L. Spector. 2020. "MALAT1 Long Non-Coding RNA: Functional Implications" Non-Coding RNA 6, no. 2: 22. https://doi.org/10.3390/ncrna6020022
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.