Tryptophan Metabolism via the Kynurenine Pathway: Implications for Graft Optimization during Machine Perfusion

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Donor Livers
2.2. Procurement of Grafts
2.3. Machine Perfusion
2.4. Energy Cofactor Analysis
2.5. Untargeted Metabolomic Analysis
2.6. Statistical Analysis
3. Results
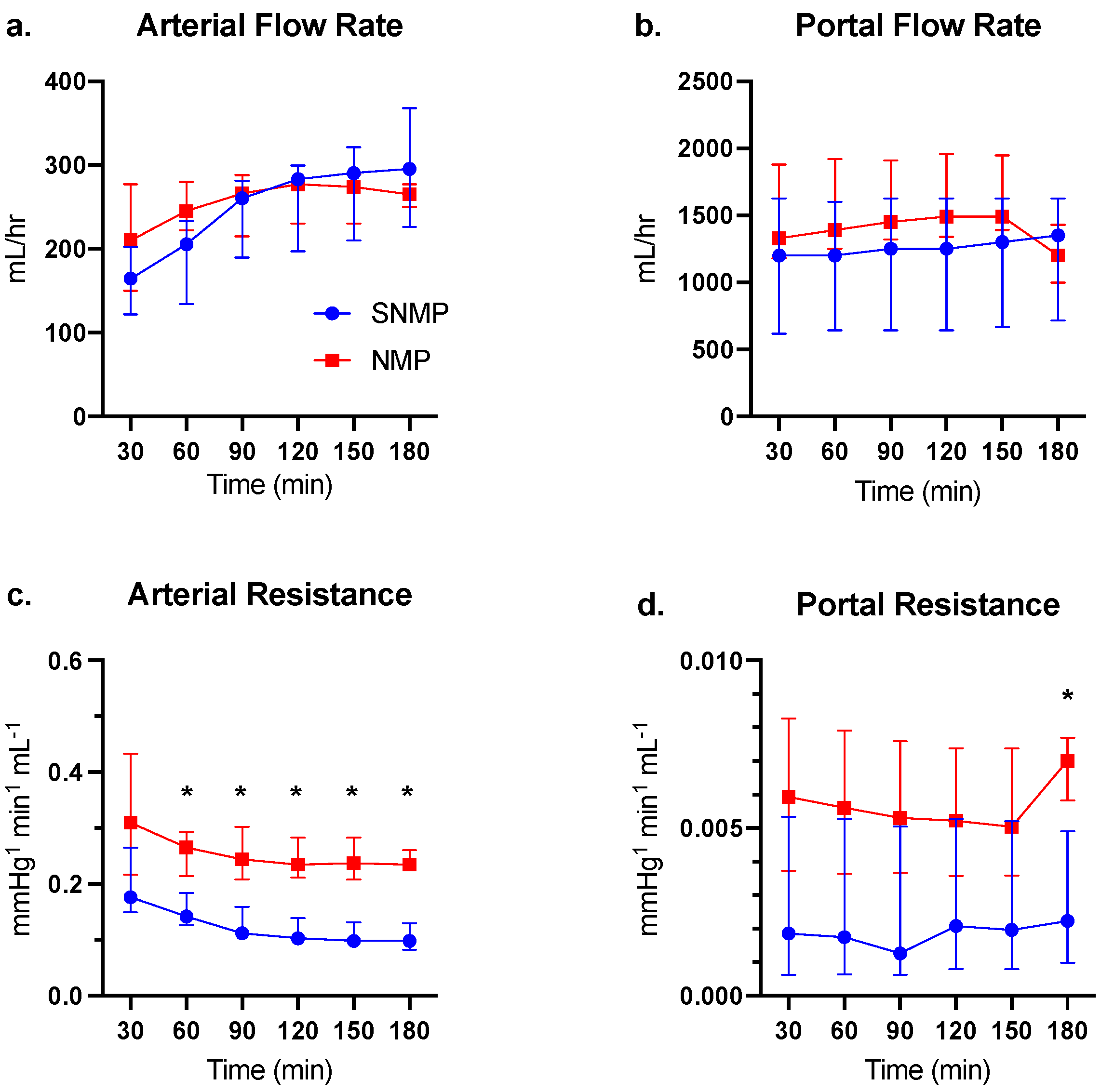
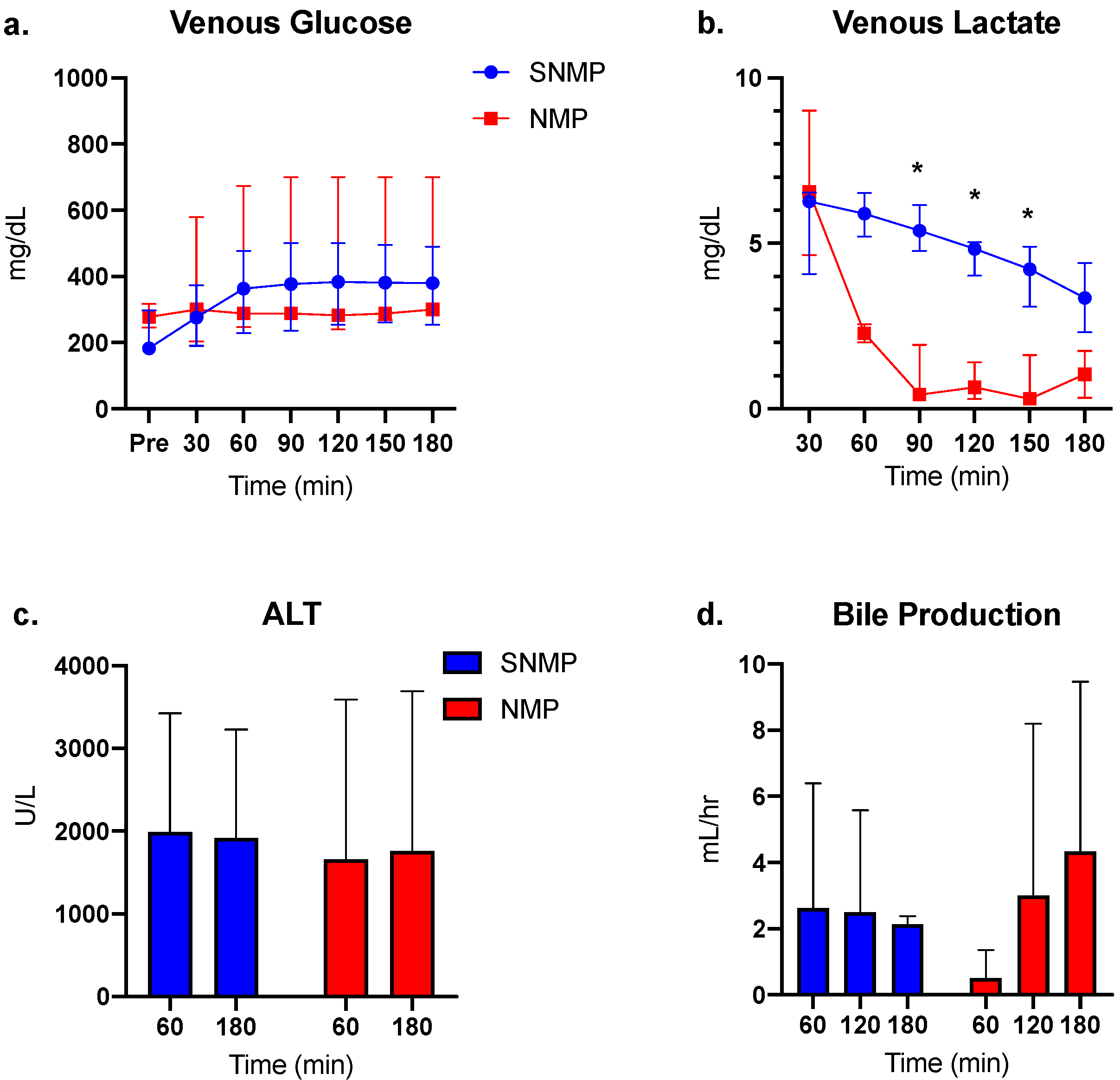
3.1. Perfusion and Functional Parameters
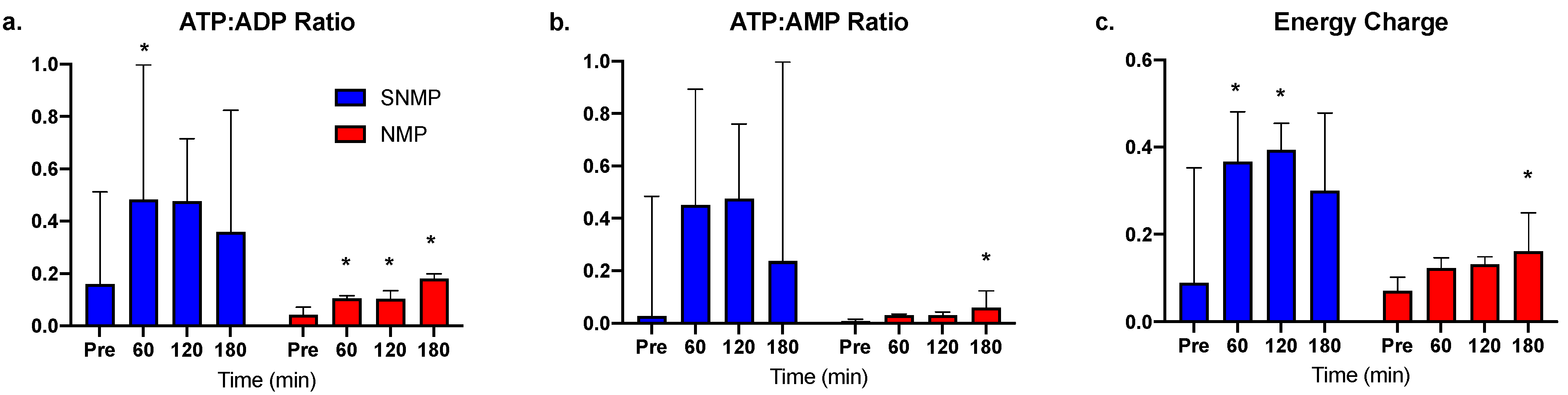
3.2. Greater ATP Conservation and Energy Charge Ratios during SNMP
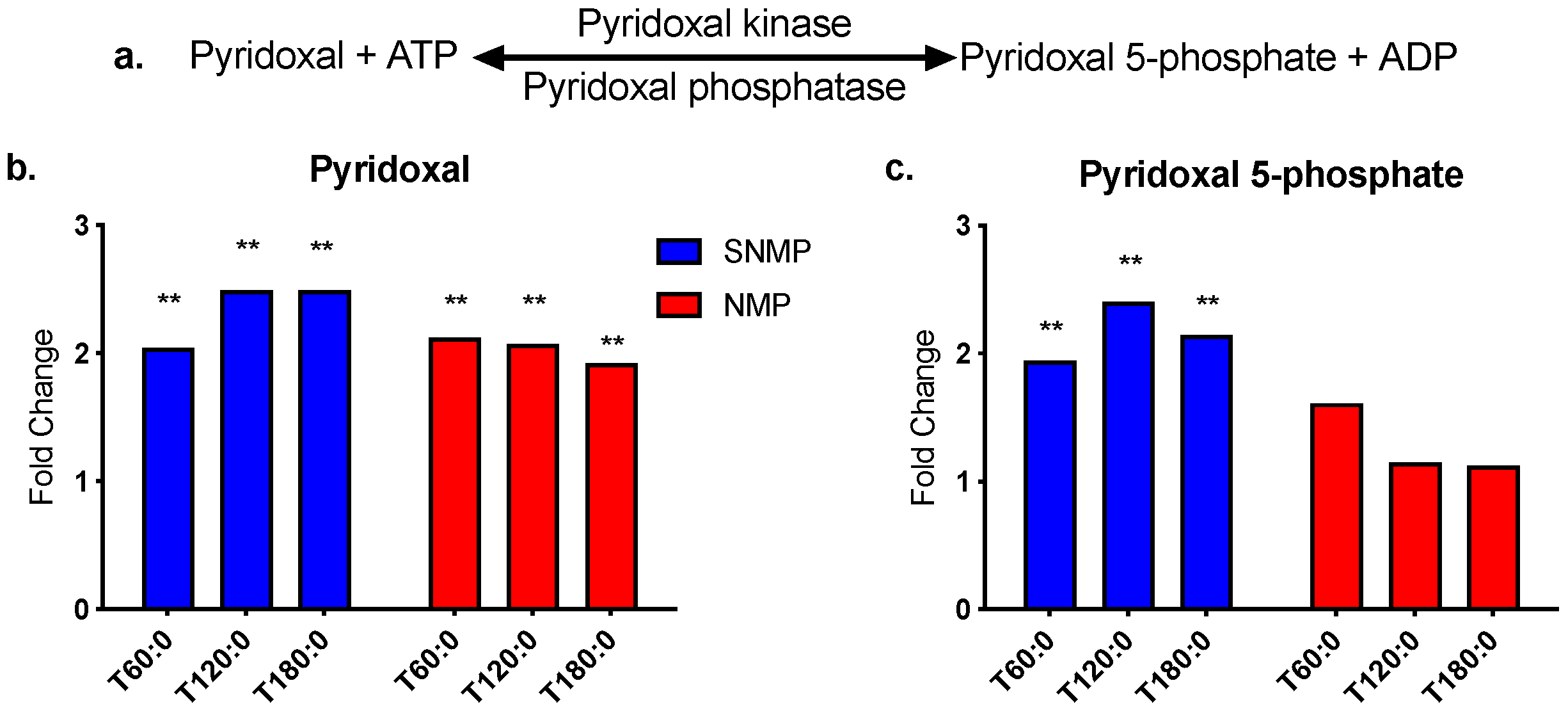
3.3. Greater Tryptophan Metabolism in Livers during NMP
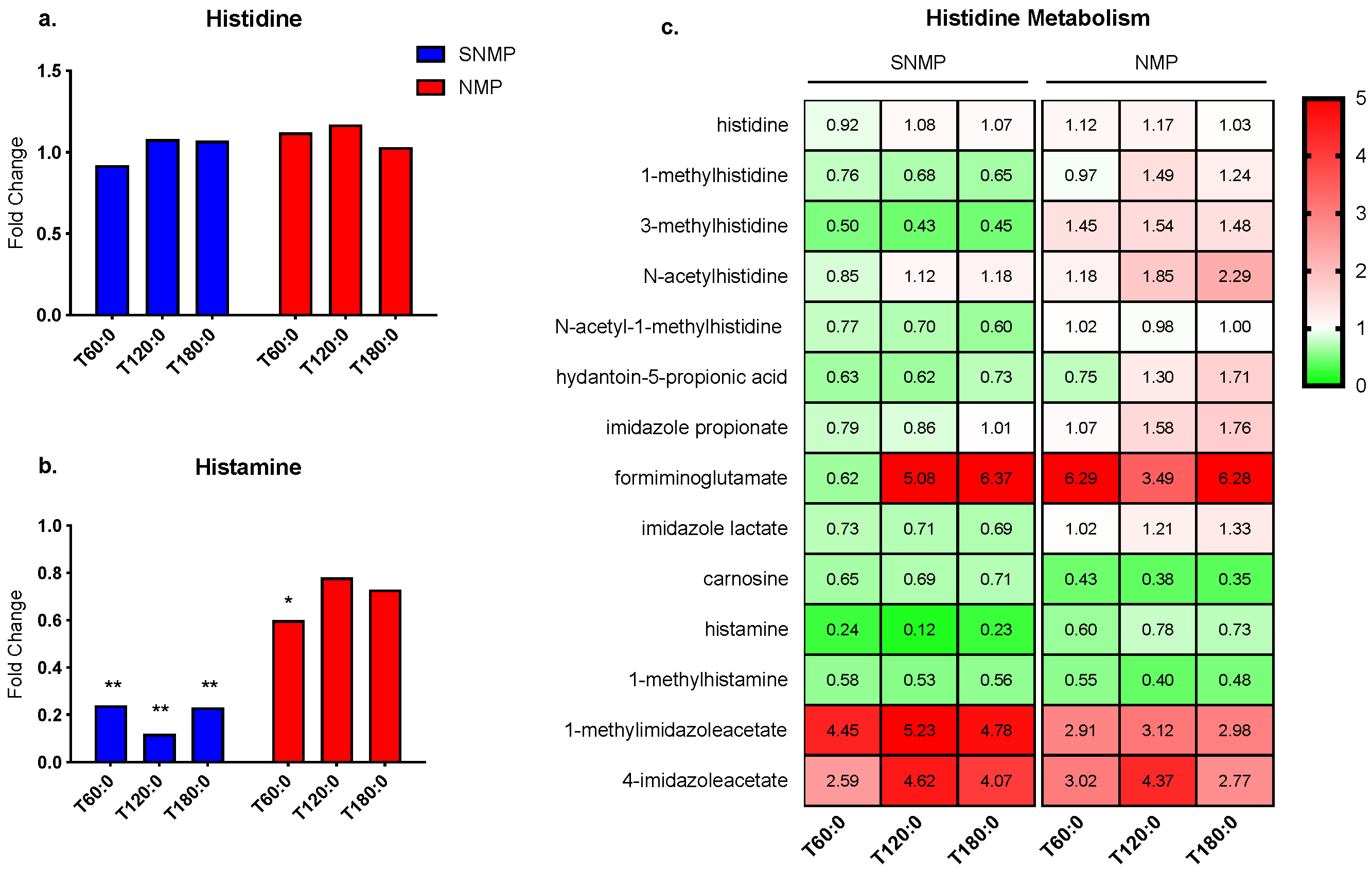
3.4. Greater Histamine Reduction in Livers during SNMP
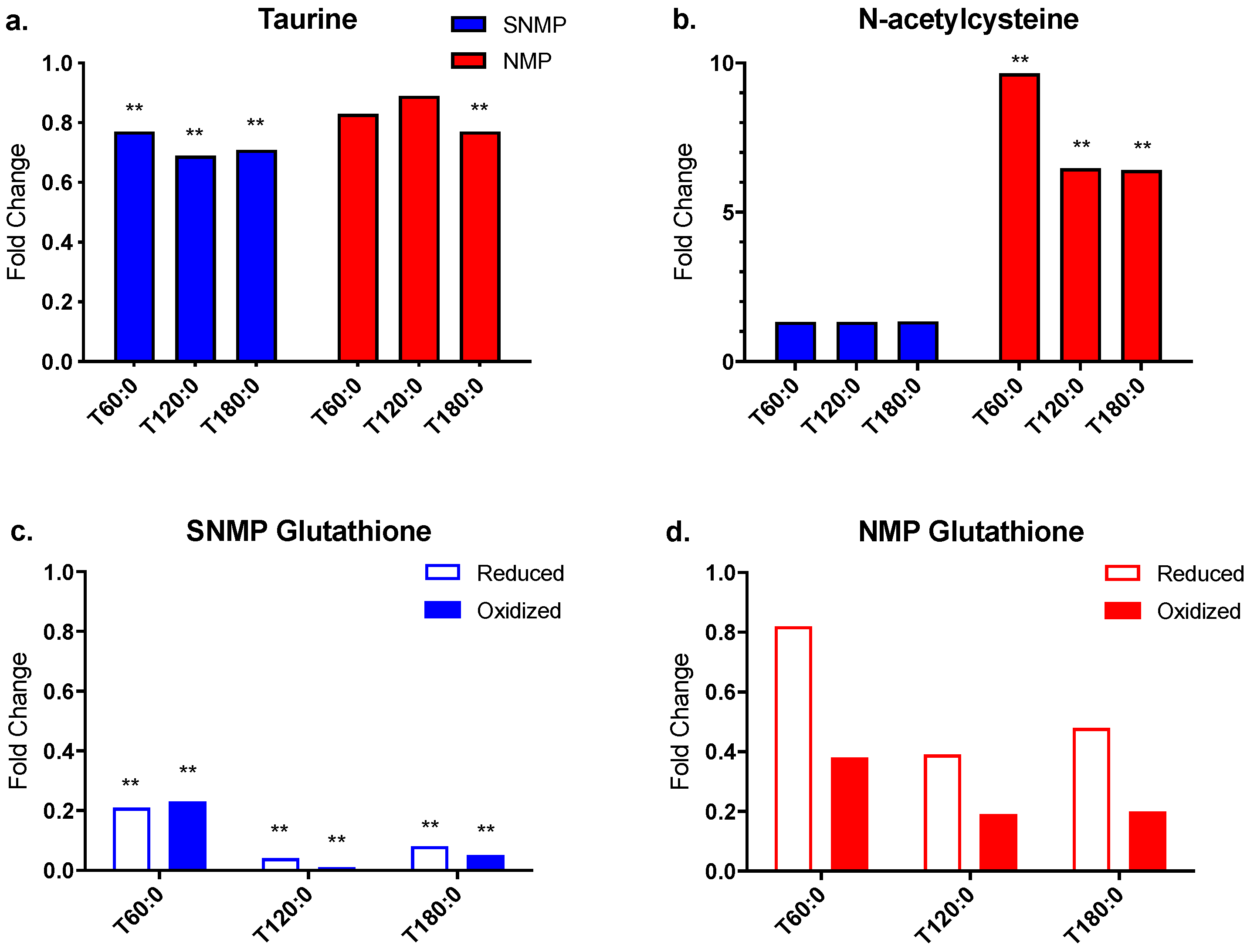
3.5. Decreased Antioxidant Capacity in Livers during SNMP
3.6. Bile Acid Metabolism in Livers during SNMP and NMP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldberg, D.S.; Reese, P.P.; Amaral, S.; Abt, P.L. Reframing the impact of combined heart-liver allocation on liver transplant wait-list candidates. Liver Transpl. 2014, 20, 1356–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raigani, S.; De Vries, R.J.; Uygun, K.; Yeh, H. Pumping new life into old ideas: Preservation and rehabilitation of the liver using ex situ machine perfusion. Artif. Organs 2020, 44, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Karimian, N.; Matton, A.P.; Westerkamp, A.C.; Burlage, L.C.; Op den Dries, S.; Leuvenink, H.G.; Lisman, T.; Uygun, K.; Markmann, J.F.; Porte, R.J. Ex Situ Normothermic Machine Perfusion of Donor Livers. J. Vis. Exp. 2015, e52688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruinsma, B.G.; Sridharan, G.V.; Weeder, P.D.; Avruch, J.H.; Saeidi, N.; Ozer, S.; Geerts, S.; Porte, R.J.; Heger, M.; van Gulik, T.M.; et al. Metabolic profiling during ex vivo machine perfusion of the human liver. Sci. Rep. 2016, 6, 22415. [Google Scholar] [CrossRef] [Green Version]
- Kron, P.; Schlegel, A.; Mancina, L.; Clavien, P.A.; Dutkowski, P. Hypothermic oxygenated perfusion (HOPE) for fatty liver grafts in rats and humans. J. Hepatol. 2017. [Google Scholar] [CrossRef]
- Hoyer, D.P.; Mathe, Z.; Gallinat, A.; Canbay, A.C.; Treckmann, J.W.; Rauen, U.; Paul, A.; Minor, T. Controlled Oxygenated Rewarming of Cold Stored Livers Prior to Transplantation: First Clinical Application of a New Concept. Transplantation 2016, 100, 147–152. [Google Scholar] [CrossRef]
- Minor, T.; Efferz, P.; Fox, M.; Wohlschlaeger, J.; Luer, B. Controlled oxygenated rewarming of cold stored liver grafts by thermally graduated machine perfusion prior to reperfusion. Am. J. Transplant. 2013, 13, 1450–1460. [Google Scholar] [CrossRef]
- Von Horn, C.; Baba, H.A.; Hannaert, P.; Hauet, T.; Leuvenink, H.; Paul, A.; Minor, T.; COPE Consortium Partners. Controlled oxygenated rewarming up to normothermia for pretransplant reconditioning of liver grafts. Clin. Transplant. 2017, 31. [Google Scholar] [CrossRef] [Green Version]
- Bruinsma, B.G.; Yeh, H.; Ozer, S.; Martins, P.N.; Farmer, A.; Wu, W.; Saeidi, N.; Op den Dries, S.; Berendsen, T.A.; Smith, R.N.; et al. Subnormothermic machine perfusion for ex vivo preservation and recovery of the human liver for transplantation. Am. J. Transplant. 2014, 14, 1400–1409. [Google Scholar] [CrossRef] [Green Version]
- Karimian, N.; Yeh, H. Opportunities for Therapeutic Intervention During Machine Perfusion. Curr. Transplant. Rep. 2017, 4, 141–148. [Google Scholar] [CrossRef]
- Martins, P.N.; Buchwald, J.E.; Mergental, H.; Vargas, L.; Quintini, C. The Role of Normothermic Machine Perfusion in Liver Transplantation. Int. J. Surg. 2020. [Google Scholar] [CrossRef] [PubMed]
- Faitot, F.; Besch, C.; Battini, S.; Ruhland, E.; Onea, M.; Addeo, P.; Woehl-Jaegle, M.L.; Ellero, B.; Bachellier, P.; Namer, I.J. Impact of real-time metabolomics in liver transplantation: Graft evaluation and donor-recipient matching. J. Hepatol. 2018, 68, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Chen, J.; Jin, M.; Zhang, Z.; Chen, W.H.; Xu, F. Dynamic metabolomic analysis of intestinal ischemia-reperfusion injury in rats. IUBMB Life 2020, 72, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Andrianova, N.V.; Popkov, V.A.; Klimenko, N.S.; Tyakht, A.V.; Baydakova, G.V.; Frolova, O.Y.; Zorova, L.D.; Pevzner, I.B.; Zorov, D.B.; Plotnikov, E.Y. Microbiome-Metabolome Signature of Acute Kidney Injury. Metabolites 2020, 10, 142. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, R.N.; Patel, S.V.B.; Sun, Q.; Jiang, L.; Richard-Mohamed, M.; Ruthirakanthan, A.; Aquil, S.; Al-Ogaili, R.; Juriasingani, S.; Sener, A.; et al. Renal Protection Against Ischemia Reperfusion Injury: Hemoglobin-based Oxygen Carrier-201 Versus Blood as an Oxygen Carrier in Ex Vivo Subnormothermic Machine Perfusion. Transplantation 2020, 104, 482–489. [Google Scholar] [CrossRef]
- Bakhta, O.; Pascaud, A.; Dieu, X.; Beaumont, J.; Kouassi Nzoughet, J.; Kamel, R.; Croyal, M.; Tamareille, S.; Simard, G.; Chao de la Barca, J.M.; et al. Tryptophane-kynurenine pathway in the remote ischemic conditioning mechanism. Basic. Res. Cardiol. 2020, 115, 1–8. [Google Scholar] [CrossRef]
- Hrydziuszko, O.; Perera, M.T.; Laing, R.; Kirwan, J.; Silva, M.A.; Richards, D.A.; Murphy, N.; Mirza, D.F.; Viant, M.R. Mass Spectrometry Based Metabolomics Comparison of Liver Grafts from Donors after Circulatory Death (DCD) and Donors after Brain Death (DBD) Used in Human Orthotopic Liver Transplantation. PLoS ONE 2016, 11, e0165884. [Google Scholar] [CrossRef]
- Cortes, M.; Pareja, E.; Garcia-Canaveras, J.C.; Donato, M.T.; Montero, S.; Mir, J.; Castell, J.V.; Lahoz, A. Metabolomics discloses donor liver biomarkers associated with early allograft dysfunction. J. Hepatol. 2014, 61, 564–574. [Google Scholar] [CrossRef]
- Xu, J.; Hassan-Ally, M.; Casas-Ferreira, A.M.; Suvitaival, T.; Ma, Y.; Vilca-Melendez, H.; Rela, M.; Heaton, N.; Wayel, J.; Legido-Quigley, C. Deregulation of the Purine Pathway in Pre-Transplant Liver Biopsies Is Associated with Graft Function and Survival after Transplantation. J. Clin. Med. 2020, 9, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, M.T.; Higdon, R.; Richards, D.A.; Silva, M.A.; Murphy, N.; Kolker, E.; Mirza, D.F. Biomarker differences between cadaveric grafts used in human orthotopic liver transplantation as identified by coulometric electrochemical array detection (CEAD) metabolomics. OMICS 2014, 18, 767–777. [Google Scholar] [CrossRef]
- Karimian, N.; Raigani, S.; Huang, V.; Nagpal, S.; Hafiz, E.O.A.; Beijert, I.; Mahboub, P.; Porte, R.J.; Uygun, K.; Yarmush, M.; et al. Subnormothermic Machine Perfusion of Steatotic Livers Results in Increased Energy Charge at the Cost of Anti-Oxidant Capacity Compared to Normothermic Perfusion. Metabolites 2019, 9, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Op den Dries, S.; Karimian, N.; Sutton, M.E.; Westerkamp, A.C.; Nijsten, M.W.; Gouw, A.S.; Wiersema-Buist, J.; Lisman, T.; Leuvenink, H.G.; Porte, R.J. Ex vivo normothermic machine perfusion and viability testing of discarded human donor livers. Am. J. Transplant. 2013, 13, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.J.E.; Kosmoliaptsis, V.; Pley, C.; Randle, L.; Fear, C.; Crick, K.; Gimson, A.E.; Allison, M.; Upponi, S.; Brais, R.; et al. Observations on the ex situ perfusion of livers for transplantation. Am. J. Transplant. 2018, 18, 2005–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laing, R.W.; Mergental, H.; Yap, C.; Kirkham, A.; Whilku, M.; Barton, D.; Curbishley, S.; Boteon, Y.L.; Neil, D.A.; Hubscher, S.G.; et al. Viability testing and transplantation of marginal livers (VITTAL) using normothermic machine perfusion: Study protocol for an open-label, non-randomised, prospective, single-arm trial. BMJ Open 2017, 7, e017733. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Breitkopf, S.B.; Yang, X.; Asara, J.M. A positive/negative ion-switching, targeted mass spectrometry-based metabolomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat. Protoc. 2012, 7, 872–881. [Google Scholar] [CrossRef] [Green Version]
- Kanai, M.; Funakoshi, H.; Takahashi, H.; Hayakawa, T.; Mizuno, S.; Matsumoto, K.; Nakamura, T. Tryptophan 2,3-dioxygenase is a key modulator of physiological neurogenesis and anxiety-related behavior in mice. Mol. Brain 2009, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Ueland, P.M.; Ulvik, A.; Rios-Avila, L.; Midttun, O.; Gregory, J.F. Direct and Functional Biomarkers of Vitamin B6 Status. Annu. Rev. Nutr. 2015, 35, 33–70. [Google Scholar] [CrossRef]
- Branco, A.; Yoshikawa, F.S.Y.; Pietrobon, A.J.; Sato, M.N. Role of Histamine in Modulating the Immune Response and Inflammation. Mediat. Inflamm. 2018, 2018, 9524075. [Google Scholar] [CrossRef]
- Mitchell, R.G.; Butt, H.R.; Code, C.F. Histamine metabolism in diseases of the liver. J. Clin. Investig. 1954, 33, 1199–1203. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.M.; Dey, A. Regulation of Glutathione in Health and Disease with Special Emphasis on Chronic Alcoholism and Hyperglycaemia Mediated Liver Injury: A Brief Perspective. Springer Sci. Rev. 2014, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cacciatore, I.; Cornacchia, C.; Pinnen, F.; Mollica, A.; Di Stefano, A. Prodrug approach for increasing cellular glutathione levels. Molecules 2010, 15, 1242–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, F.; Miggiano, R.; Ferraris, D.M.; Rizzi, M. The Synthesis of Kynurenic Acid in Mammals: An Updated Kynurenine Aminotransferase Structural KATalogue. Front. Mol. Biosci. 2019, 6, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.; Cai, T.; Tagle, D.A.; Li, J. Thermal stability, pH dependence and inhibition of four murine kynurenine aminotransferases. BMC Biochem. 2010, 11, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.; Cai, T.; Tagle, D.A.; Robinson, H.; Li, J. Substrate specificity and structure of human aminoadipate aminotransferase/kynurenine aminotransferase II. Biosci. Rep. 2008, 28, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Liu, D.; Song, P.; Zou, M.H. Tryptophan-kynurenine pathway is dysregulated in inflammation, and immune activation. Front. Biosci. 2015, 20, 1116–1143. [Google Scholar] [CrossRef] [Green Version]
- Badawy, A.A. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan. Res. 2017, 10, 1178646917691938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, W.; Gao, L.; Zhou, Z.; Lin, H.; Chen, C.; Huang, P.; Huang, W.; Zhou, C.; Huang, S.; Nie, L.; et al. Indoleamine 2,3-dioxygenase 1 deficiency attenuates CCl4-induced fibrosis through Th17 cells down-regulation and tryptophan 2,3-dioxygenase compensation. Oncotarget 2017, 8, 40486–40500. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.F.; Yang, T.H.; Zhong, C.P.; Shen, C.; Lin, W.W.; Gu, G.X.; Xia, Q.; Xu, N. Dual Effect of Hepatic Macrophages on Liver Ischemia and Reperfusion Injury during Liver Transplantation. Immune Netw. 2018, 18, e24. [Google Scholar] [CrossRef]
- Yan, M.L.; Wang, Y.D.; Tian, Y.F.; Lai, Z.D.; Yan, L.N. Inhibition of allogeneic T-cell response by Kupffer cells expressing indoleamine 2,3-dioxygenase. World. J. Gastroenterol. 2010, 16, 636–640. [Google Scholar] [CrossRef]
- Schmidt, S.K.; Muller, A.; Heseler, K.; Woite, C.; Spekker, K.; MacKenzie, C.R.; Daubener, W. Antimicrobial and immunoregulatory properties of human tryptophan 2,3-dioxygenase. Eur. J. Immunol. 2009, 39, 2755–2764. [Google Scholar] [CrossRef]
- Giorgini, F.; Huang, S.Y.; Sathyasaikumar, K.V.; Notarangelo, F.M.; Thomas, M.A.; Tararina, M.; Wu, H.Q.; Schwarcz, R.; Muchowski, P.J. Targeted deletion of kynurenine 3-monooxygenase in mice: a new tool for studying kynurenine pathway metabolism in periphery and brain. J. Biol. Chem. 2013, 288, 36554–36566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarugi, A.; Carpenedo, R.; Molina, M.T.; Mattoli, L.; Pellicciari, R.; Moroni, F. Comparison of the neurochemical and behavioral effects resulting from the inhibition of kynurenine hydroxylase and/or kynureninase. J. Neurochem. 1995, 65, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Francis, H.; Franchitto, A.; Ueno, Y.; Glaser, S.; DeMorrow, S.; Venter, J.; Gaudio, E.; Alvaro, D.; Fava, G.; Marzioni, M.; et al. H3 histamine receptor agonist inhibits biliary growth of BDL rats by downregulation of the cAMP-dependent PKA/ERK1/2/ELK-1 pathway. Lab. Investig. 2007, 87, 473–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, H.; Glaser, S.; Demorrow, S.; Gaudio, E.; Ueno, Y.; Venter, J.; Dostal, D.; Onori, P.; Franchitto, A.; Marzioni, M.; et al. Small mouse cholangiocytes proliferate in response to H1 histamine receptor stimulation by activation of the IP3/CaMK I/CREB pathway. Am. J. Physiol. Cell. Physiol. 2008, 295, C499–C513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, H.L.; Demorrow, S.; Franchitto, A.; Venter, J.K.; Mancinelli, R.A.; White, M.A.; Meng, F.; Ueno, Y.; Carpino, G.; Renzi, A.; et al. Histamine stimulates the proliferation of small and large cholangiocytes by activation of both IP3/Ca2+ and cAMP-dependent signaling mechanisms. Lab. Investig. 2012, 92, 282–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, W.; Boeckl, O.; Struck, E.; Hell, E.; Zimmermann, G.; Reimann, H.J.; Tauber, R. Significance and causes of histamine release during orthopic homologous liver transplantation in the pig. Agents Actions 1973, 3, 2–11. [Google Scholar] [CrossRef]
- Wu, T.; Gan, X.; Zhou, S.; Ge, M.; Zhang, Z.; Hei, Z. Histamine at low concentrations aggravates rat liver BRL-3A cell injury induced by hypoxia/reoxygenation through histamine H2 receptor in vitro. Toxicol. In Vitro 2013, 27, 378–386. [Google Scholar] [CrossRef]
- Shaik, I.H.; Mehvar, R. Effects of cytochrome p450 inhibition by cimetidine on the warm hepatic ischemia-reperfusion injury in rats. J. Surg. Res. 2010, 159, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Farrell, D.J.; Hines, J.E.; Walls, A.F.; Kelly, P.J.; Bennett, M.K.; Burt, A.D. Intrahepatic mast cells in chronic liver diseases. Hepatology 1995, 22, 1175–1181. [Google Scholar] [CrossRef]
- Jarido, V.; Kennedy, L.; Hargrove, L.; Demieville, J.; Thomson, J.; Stephenson, K.; Francis, H. The emerging role of mast cells in liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G89–G101. [Google Scholar] [CrossRef]
- Koruk, S.T.; Ozardali, I.; Dincoglu, D.; Bitiren, M. Increased liver mast cells in patients with chronic hepatitis C. Indian J. Pathol. Microbiol. 2011, 54, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi-Hashjin, G.; Nijkamp, F.P.; Henricks, P.A.; Folkerts, G. Sodium cromoglycate and doxantrazole are oxygen radical scavengers. Eur. Respir. J. 2002, 20, 867–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laing, R.W.; Bhogal, R.H.; Wallace, L.; Boteon, Y.; Neil, D.A.H.; Smith, A.; Stephenson, B.T.F.; Schlegel, A.; Hubscher, S.G.; Mirza, D.F.; et al. The Use of an Acellular Oxygen Carrier in a Human Liver Model of Normothermic Machine Perfusion. Transplantation 2017, 101, 2746–2756. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Liver # | Reason for Discard |
|---|---|---|
| NMP | 1 | No appropriate recipient, maximum cold ischemic time exceeded |
| 2 | DCD with prolonged WIT | |
| 3 | DCD with prolonged WIT, history of alcohol abuse | |
| SNMP | 1 | DCD with prolonged WIT |
| 2 | DCD in donor >50 years of age | |
| 3 | DCD with prolonged WIT | |
| 4 | DCD with prolonged WIT |
| NMP (n = 3) | SNMP (n = 4) | p Value | |
|---|---|---|---|
| Age (years) | 44 (28–60) | 49 (33.5–52) | 0.64 |
| Gender (male) | 2 (67%) | 4 (100%) | 0.43 |
| BMI (kg/m2) | 24.7 (16.9–32.5) | 28.2 (26.1–32.1) | 0.64 |
| DCD Recovery | 2 (67%) | 4 (100%) | 0.43 |
| WIT (min) | 34 (33–35) | 30 (20–33) | 0.14 |
| CIT (min) | 690 (360–930) | 754 (685.5–861) | 0.48 |
| Liver weight (g) | 1350 (1300–2200) | 2139 (1646–2614) | 0.28 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, A.; Carroll, C.; Raigani, S.; Karimian, N.; Huang, V.; Nagpal, S.; Beijert, I.; Porte, R.J.; Yarmush, M.; Uygun, K.; et al. Tryptophan Metabolism via the Kynurenine Pathway: Implications for Graft Optimization during Machine Perfusion. J. Clin. Med. 2020, 9, 1864. https://doi.org/10.3390/jcm9061864
Zhang A, Carroll C, Raigani S, Karimian N, Huang V, Nagpal S, Beijert I, Porte RJ, Yarmush M, Uygun K, et al. Tryptophan Metabolism via the Kynurenine Pathway: Implications for Graft Optimization during Machine Perfusion. Journal of Clinical Medicine. 2020; 9(6):1864. https://doi.org/10.3390/jcm9061864
Chicago/Turabian StyleZhang, Anna, Cailah Carroll, Siavash Raigani, Negin Karimian, Viola Huang, Sonal Nagpal, Irene Beijert, Robert J. Porte, Martin Yarmush, Korkut Uygun, and et al. 2020. "Tryptophan Metabolism via the Kynurenine Pathway: Implications for Graft Optimization during Machine Perfusion" Journal of Clinical Medicine 9, no. 6: 1864. https://doi.org/10.3390/jcm9061864