Progenitor Cells Derived from Drain Waste Product of Open-Heart Surgery in Children

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
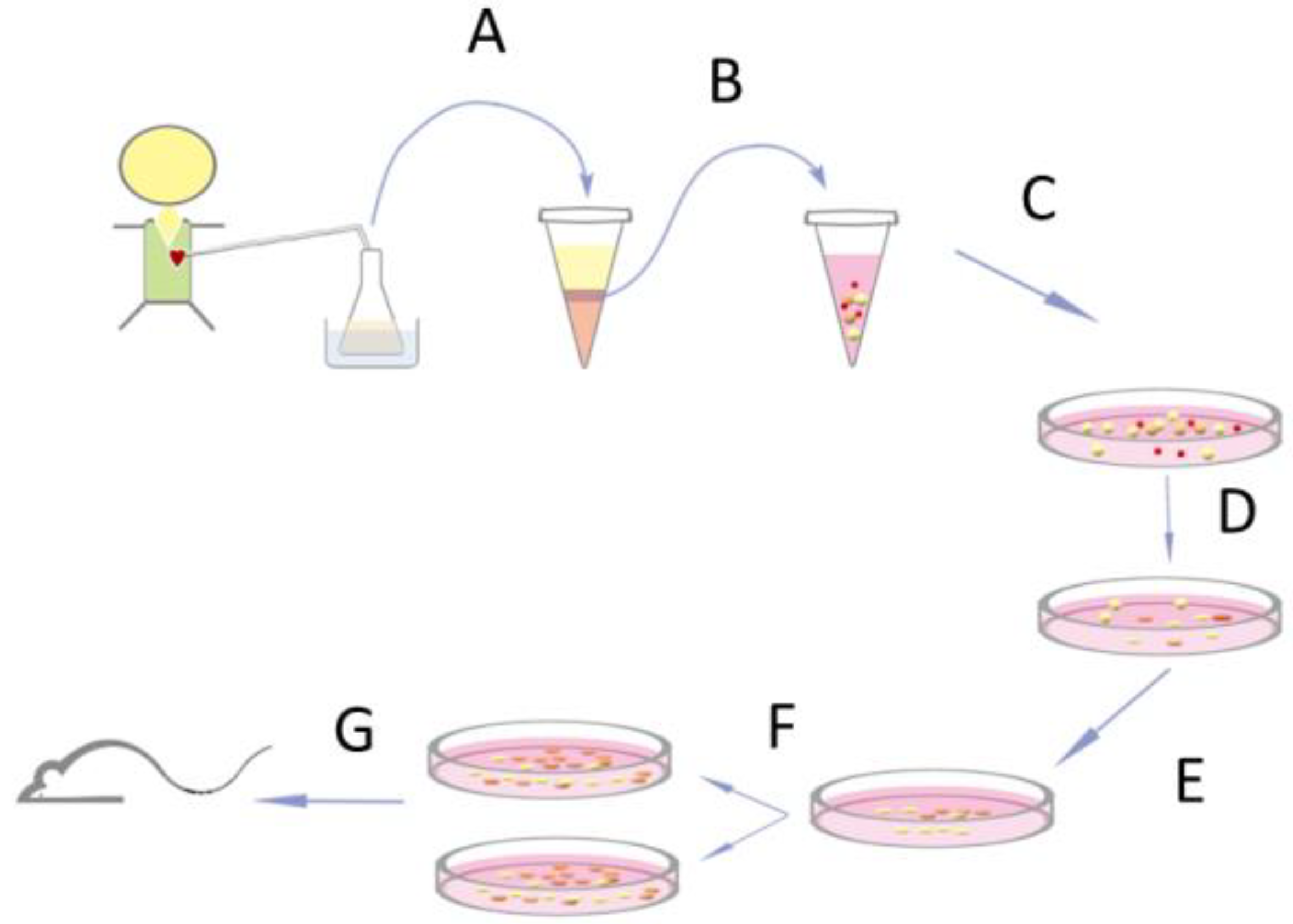
2.1. Isolation and Expansion of Progenitor Cells from Open-Heart Surgery Drain Fluids
2.2. Flow Cytometry
2.3. Cell Culture and Conditioned Medium for Angiogenesis Assay
2.4. Cardiac Infarction in Mice
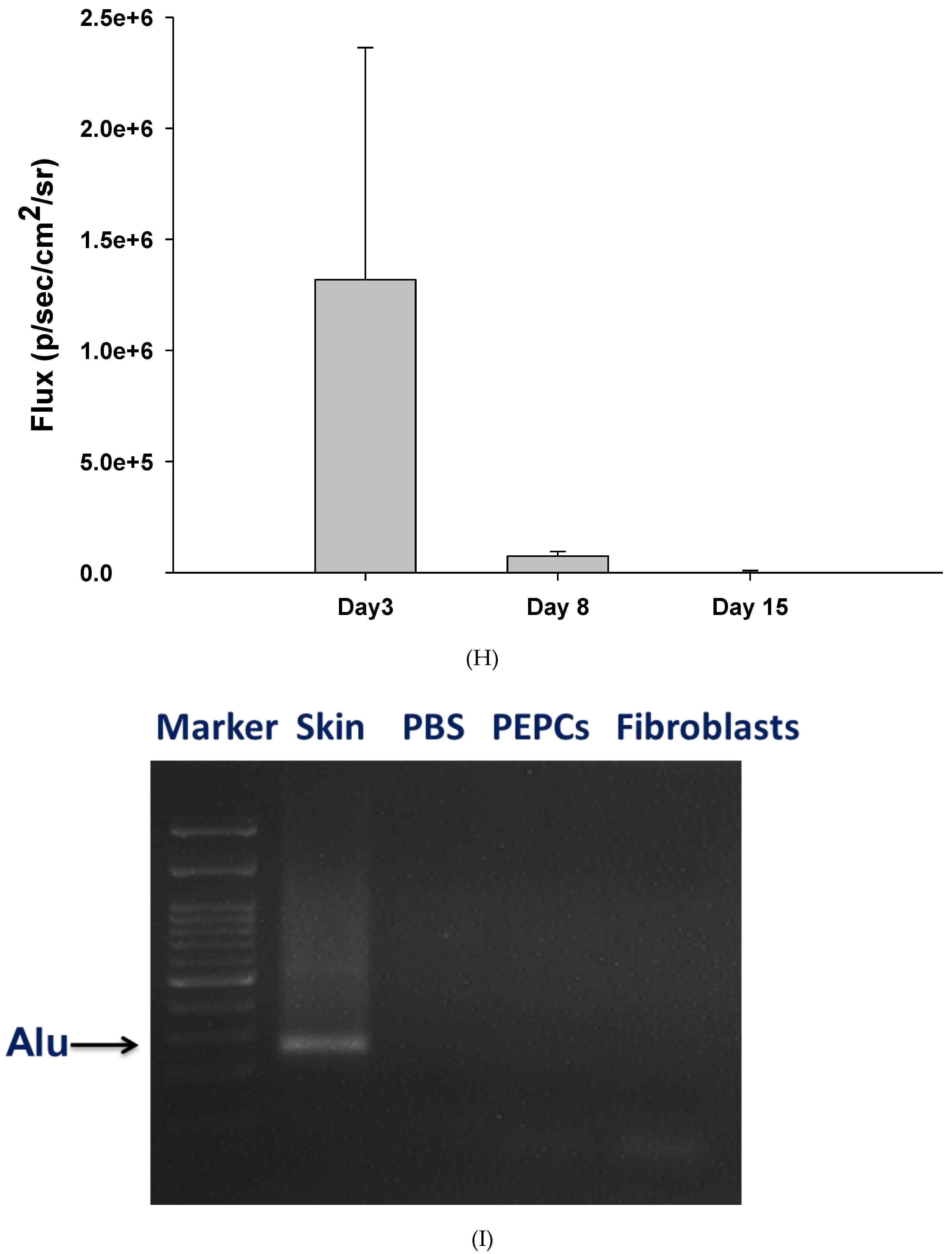
2.5. Transplanted Cell Tracing with Optical Bioluminescence Imaging and Alu Sequence
2.6. Virus Production and Cell Transduction
2.7. Immunocytochemistry and Histological Analysis
2.8. Gene Expression in Mouse Heart after Cell Transplantation
2.9. Statistics Analysis
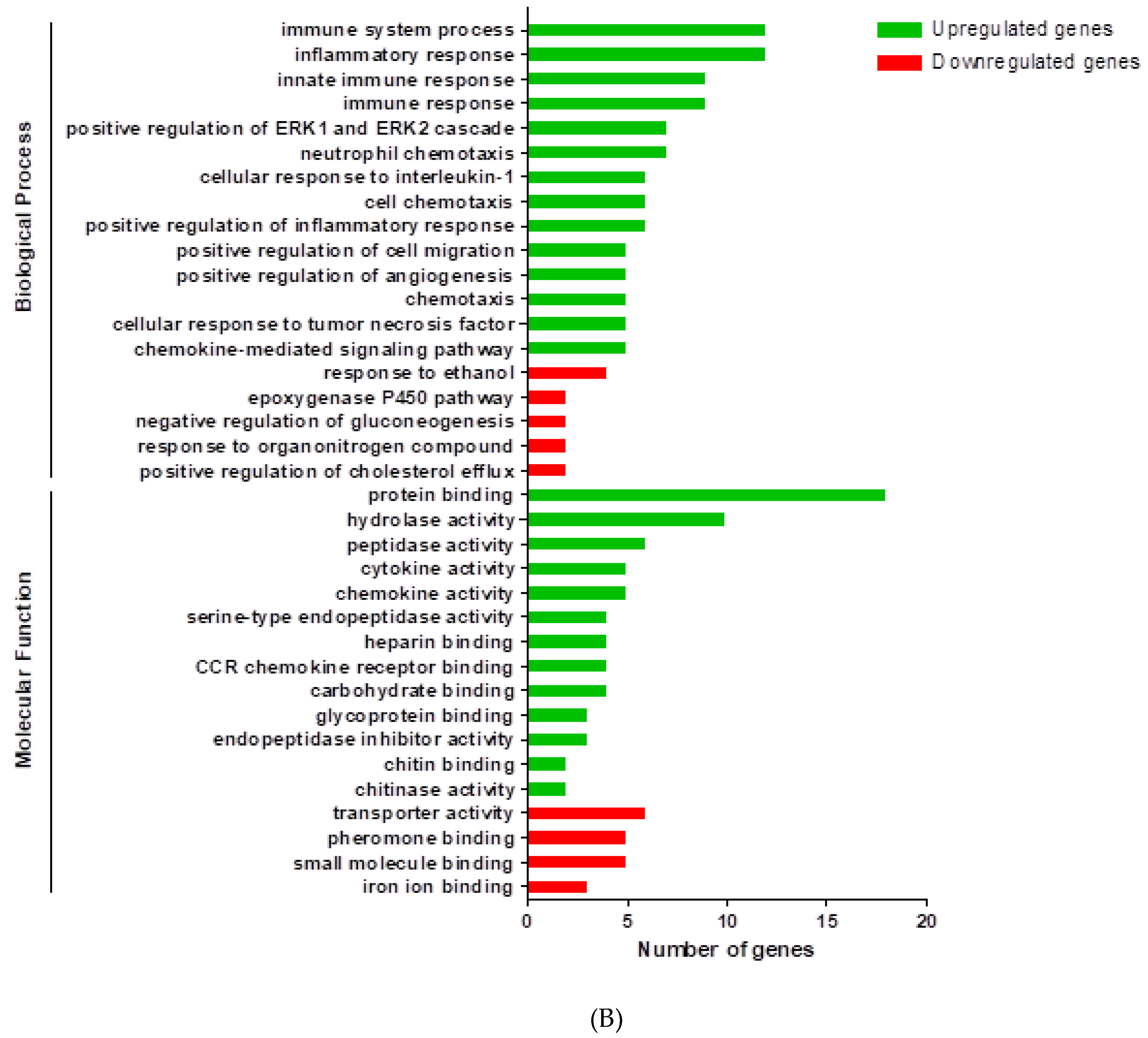
3. Results
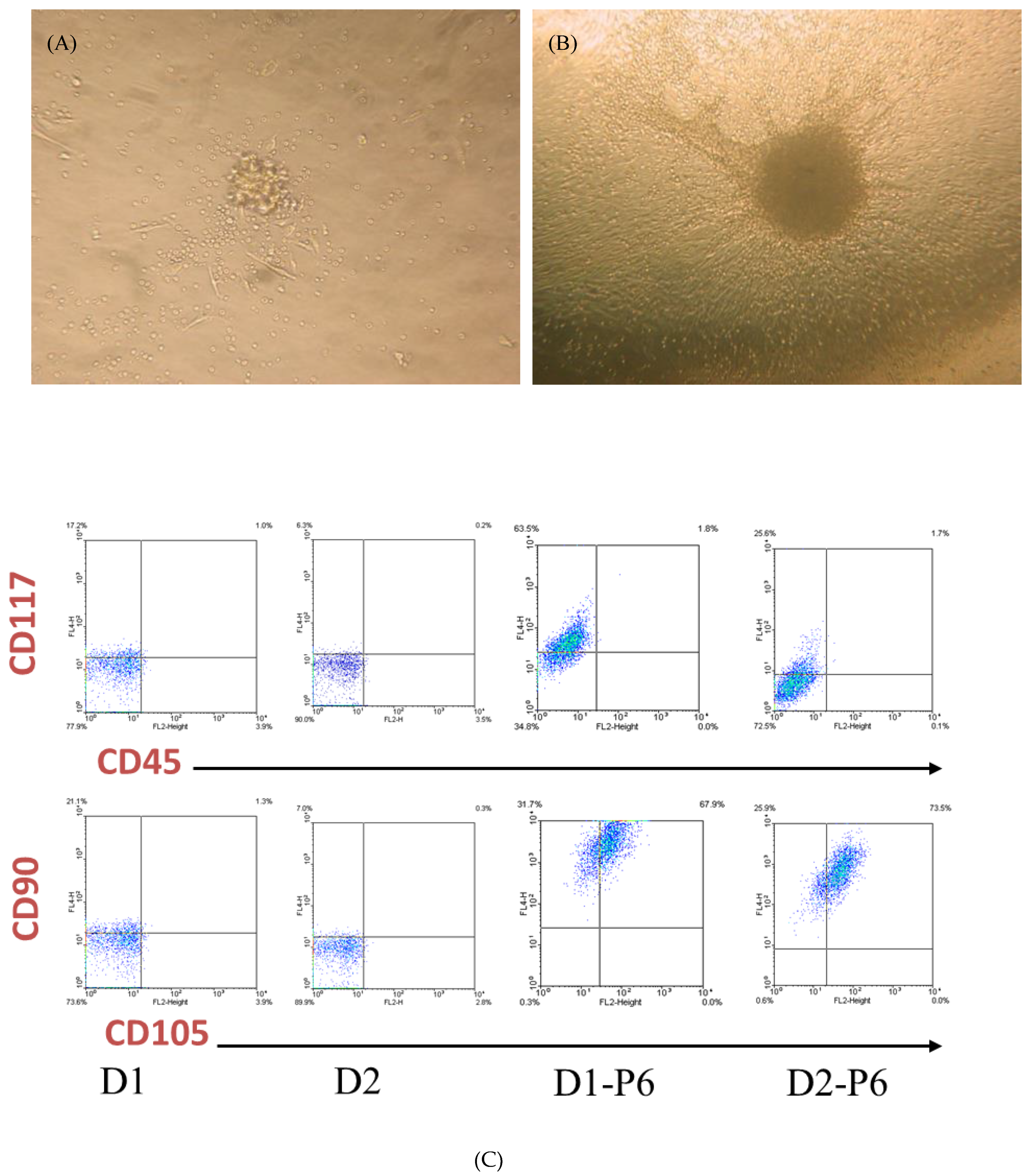
3.1. Isolation of PEPCs from Drain Fluid after Open-Heart Surgery
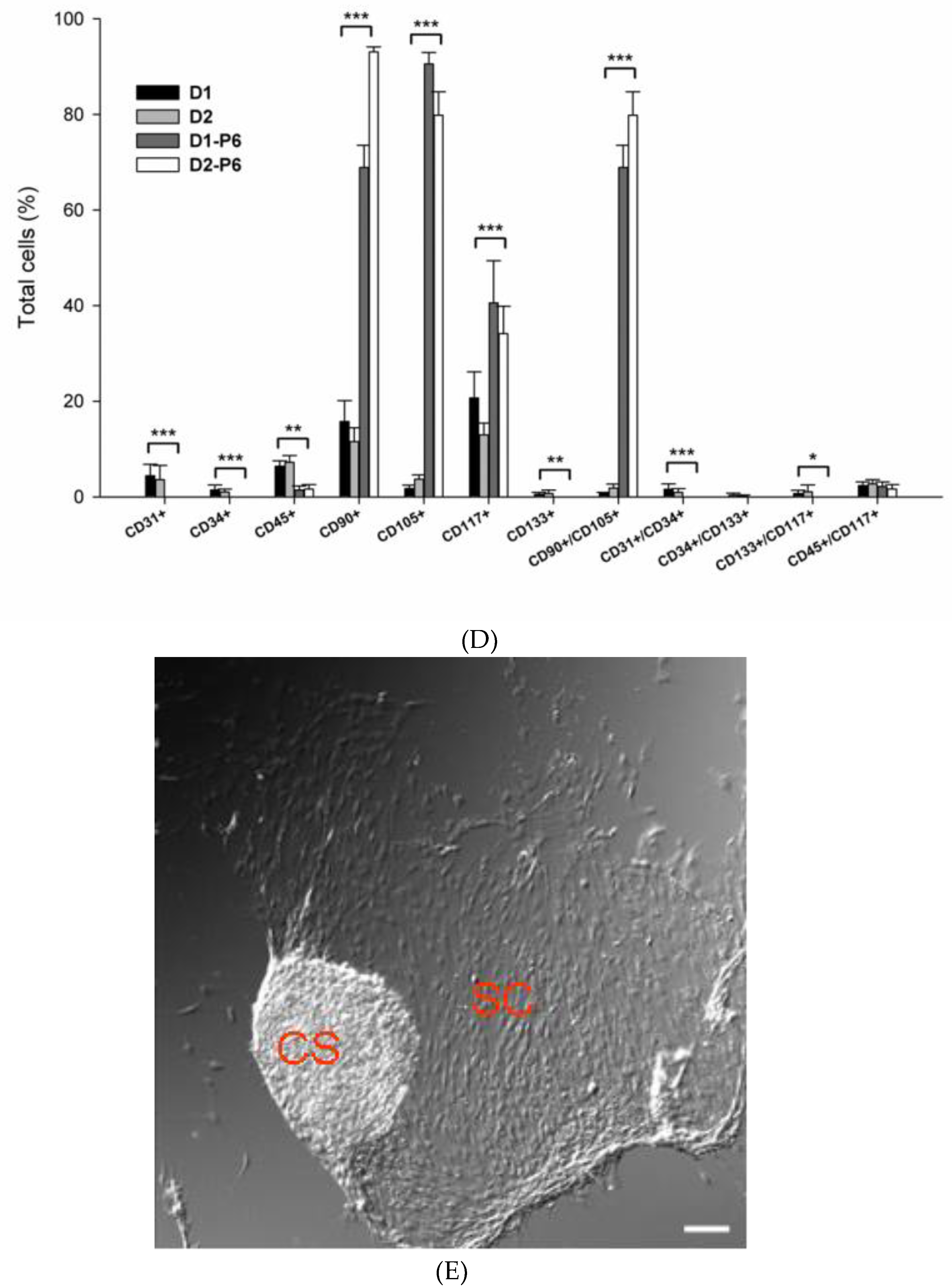
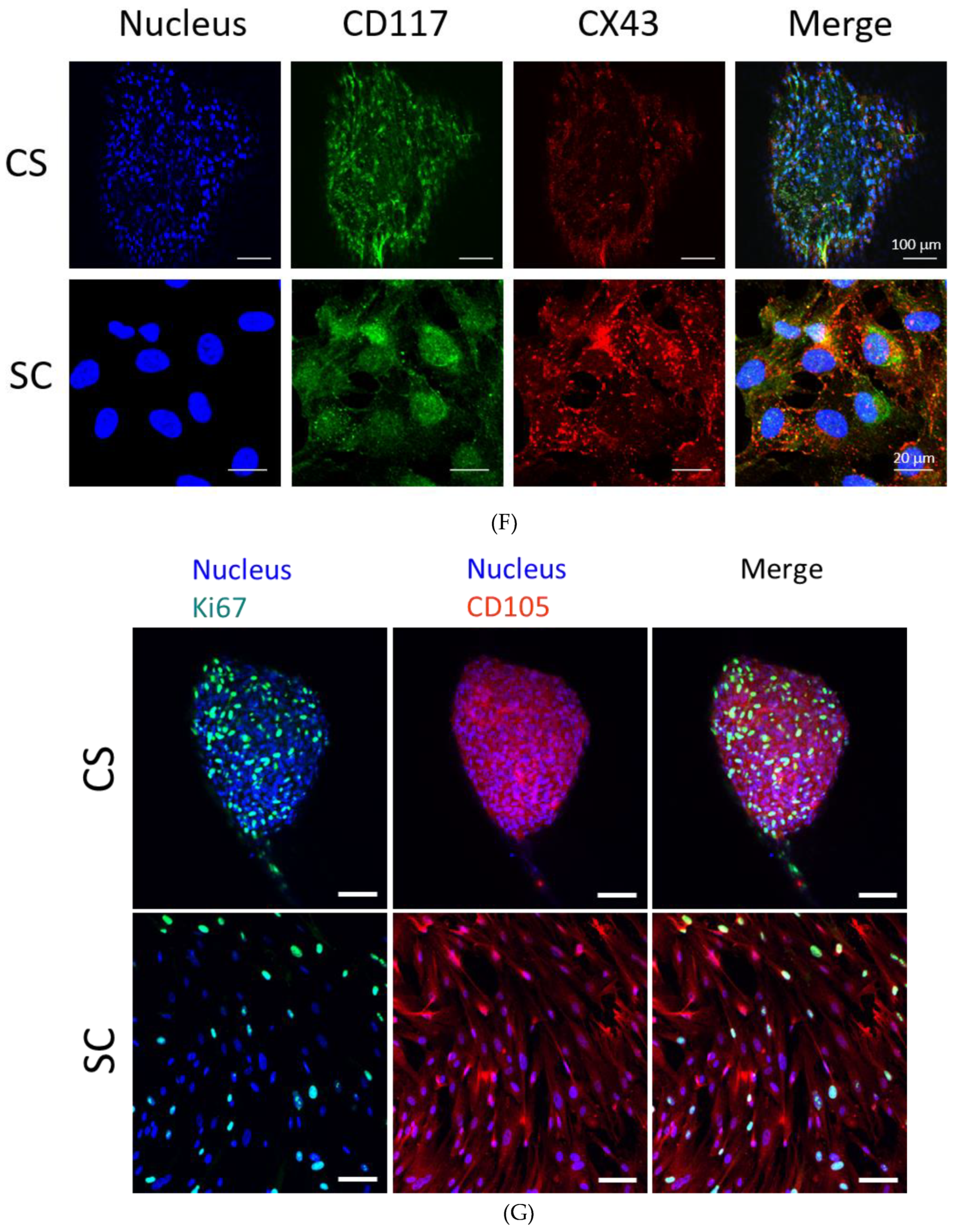
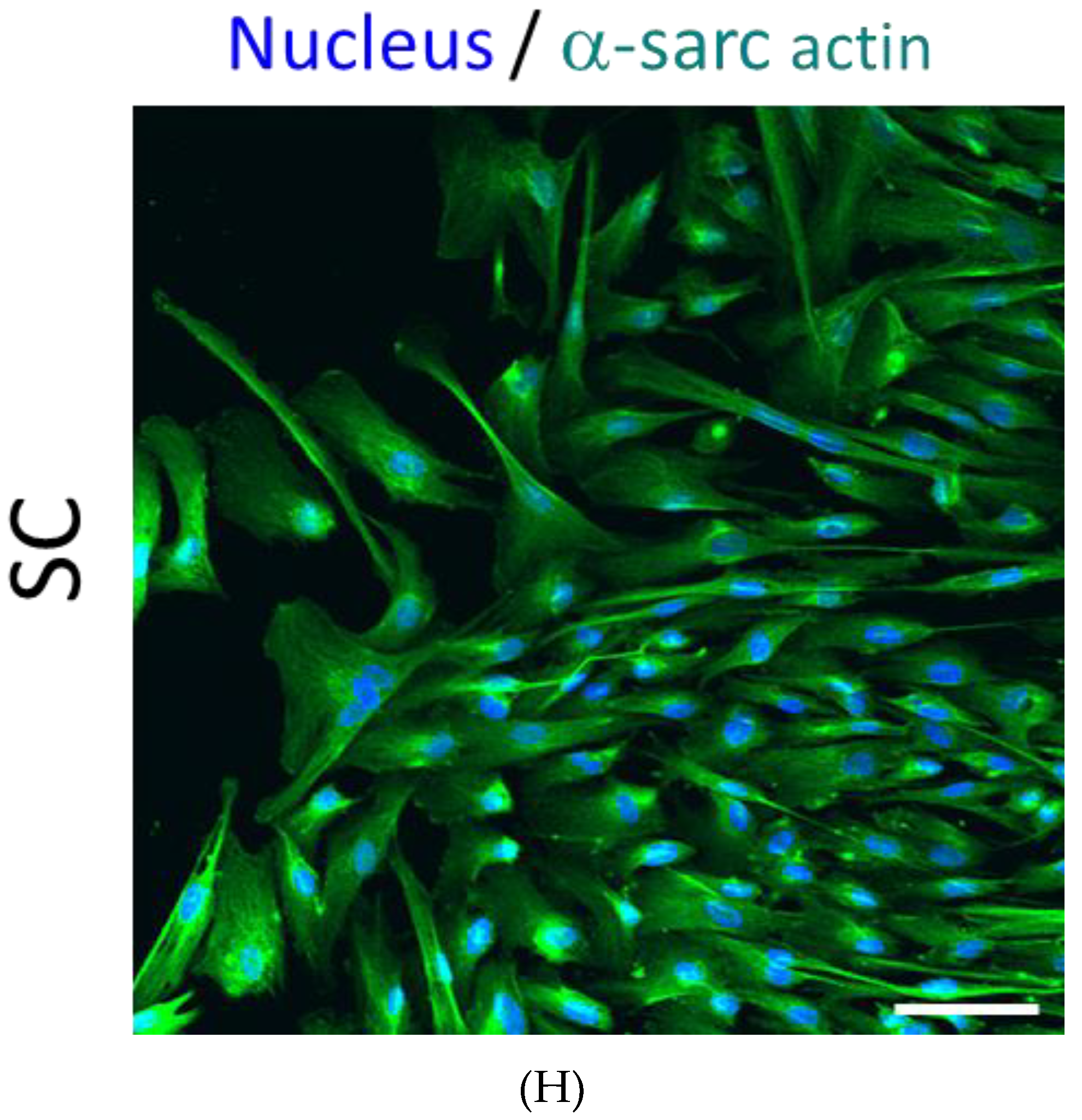
3.2. PEPCs Differentiate toward Cardiovascular Lineage
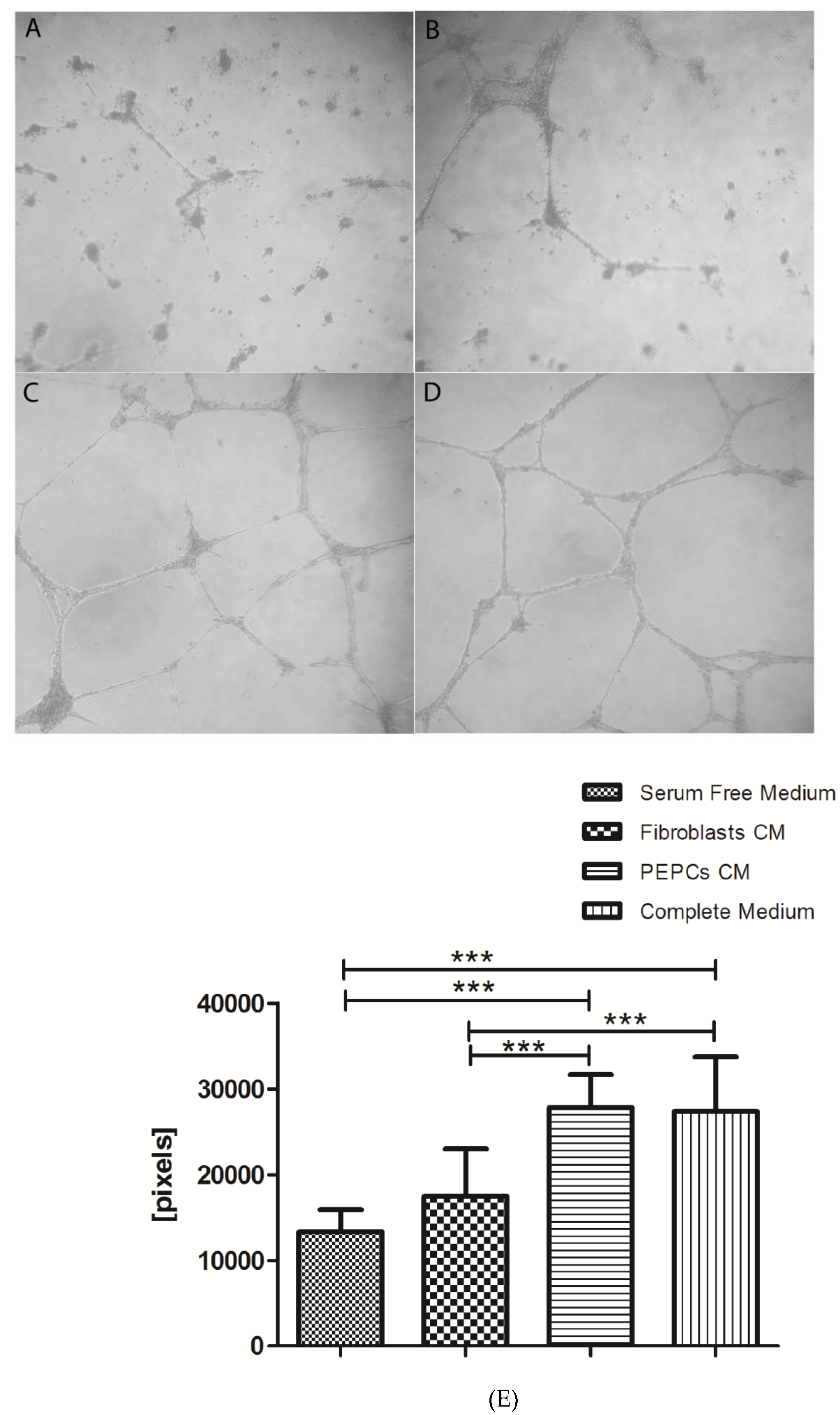
3.3. In Vitro Angiogenesis of PEPCs
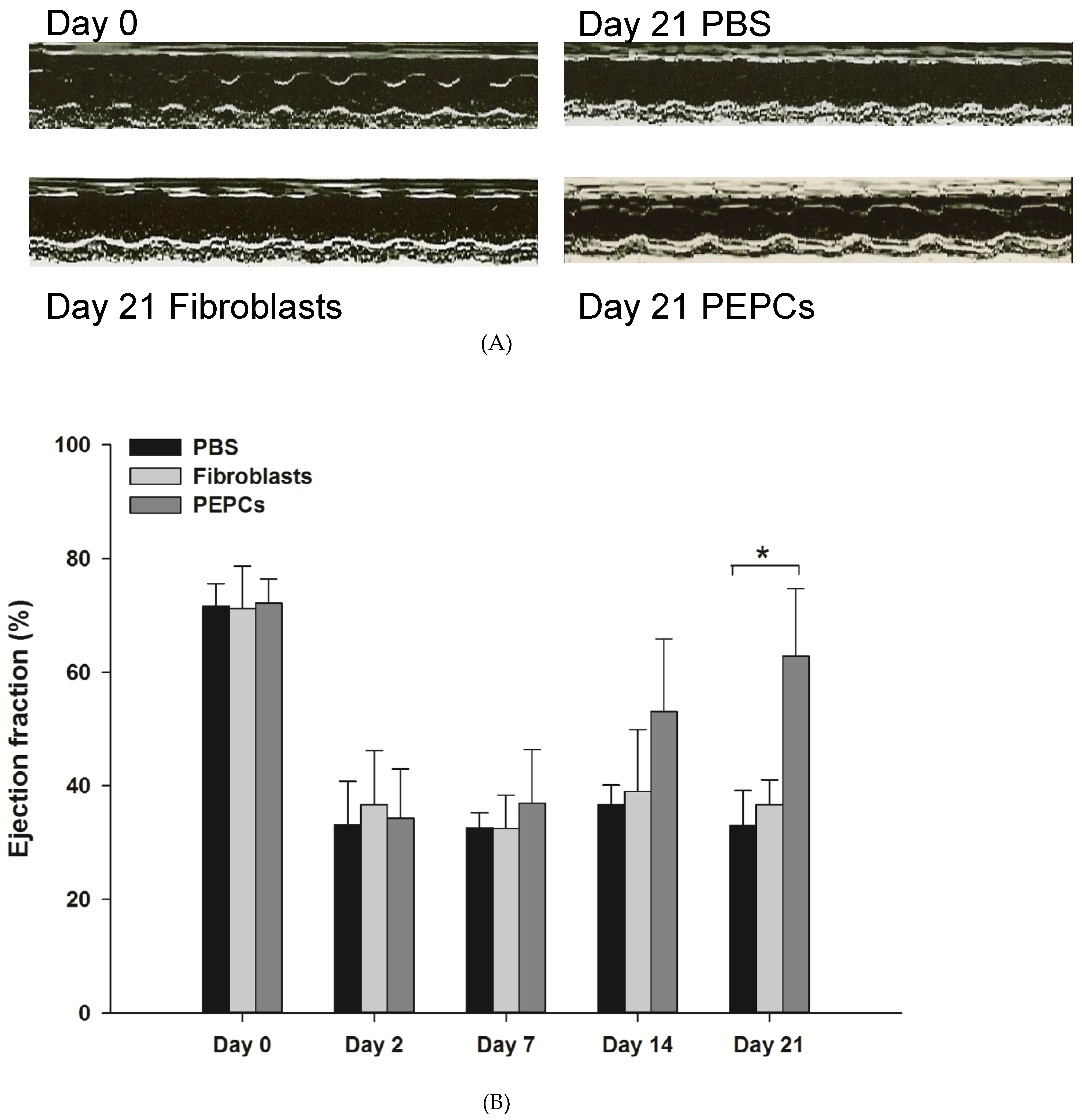
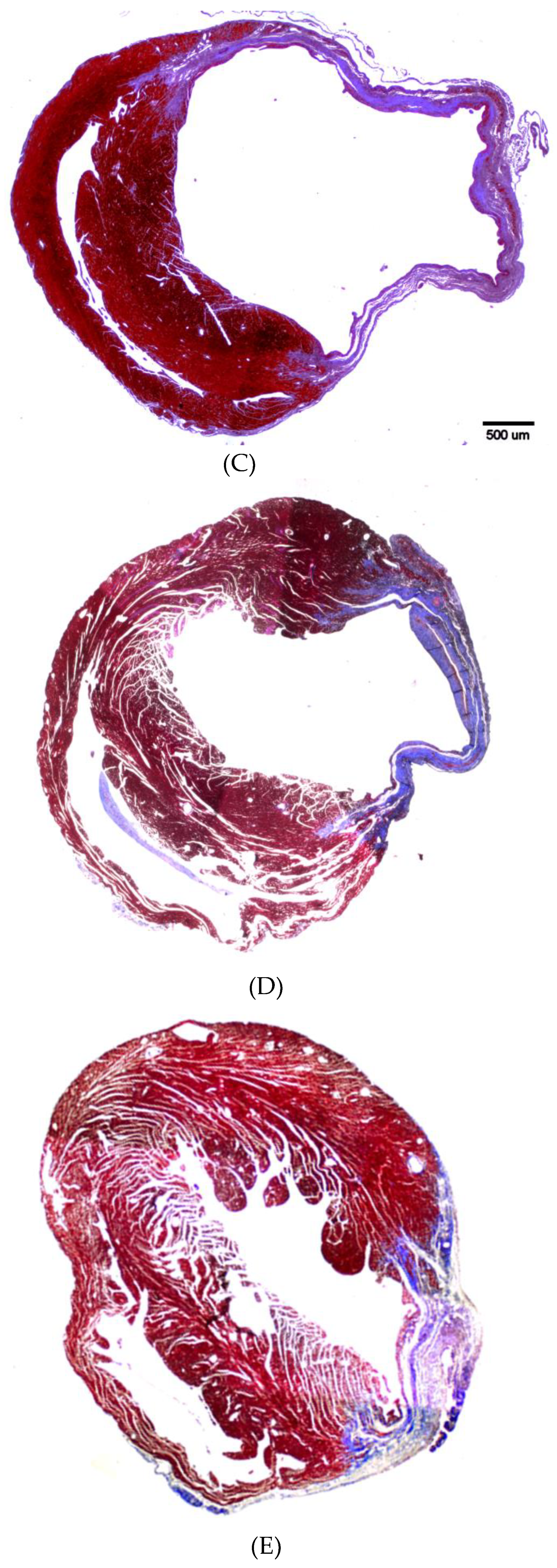
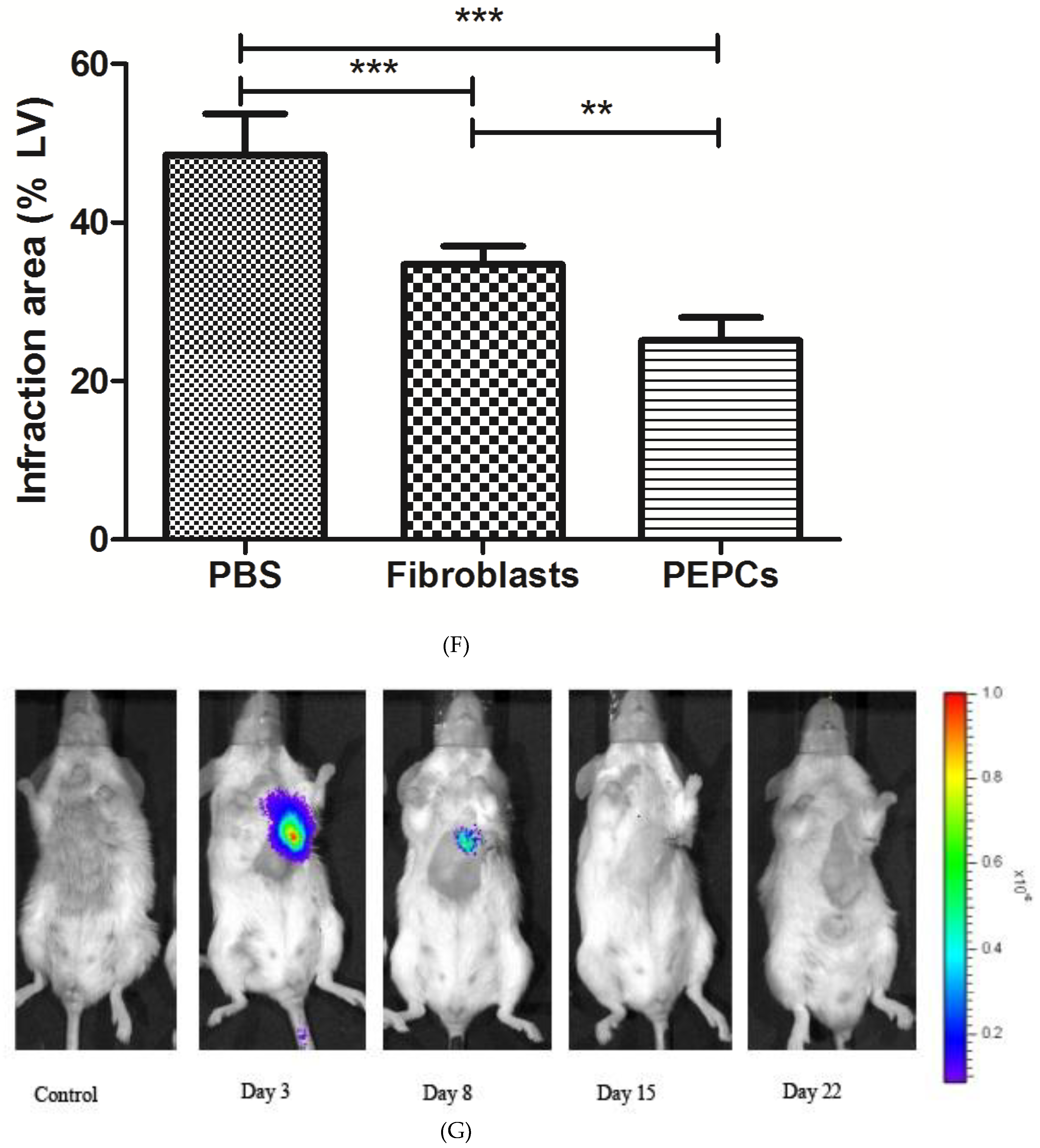
3.4. Therapeutic Potential of PEPCs and Cell Fate in Ischemic Heart Repair
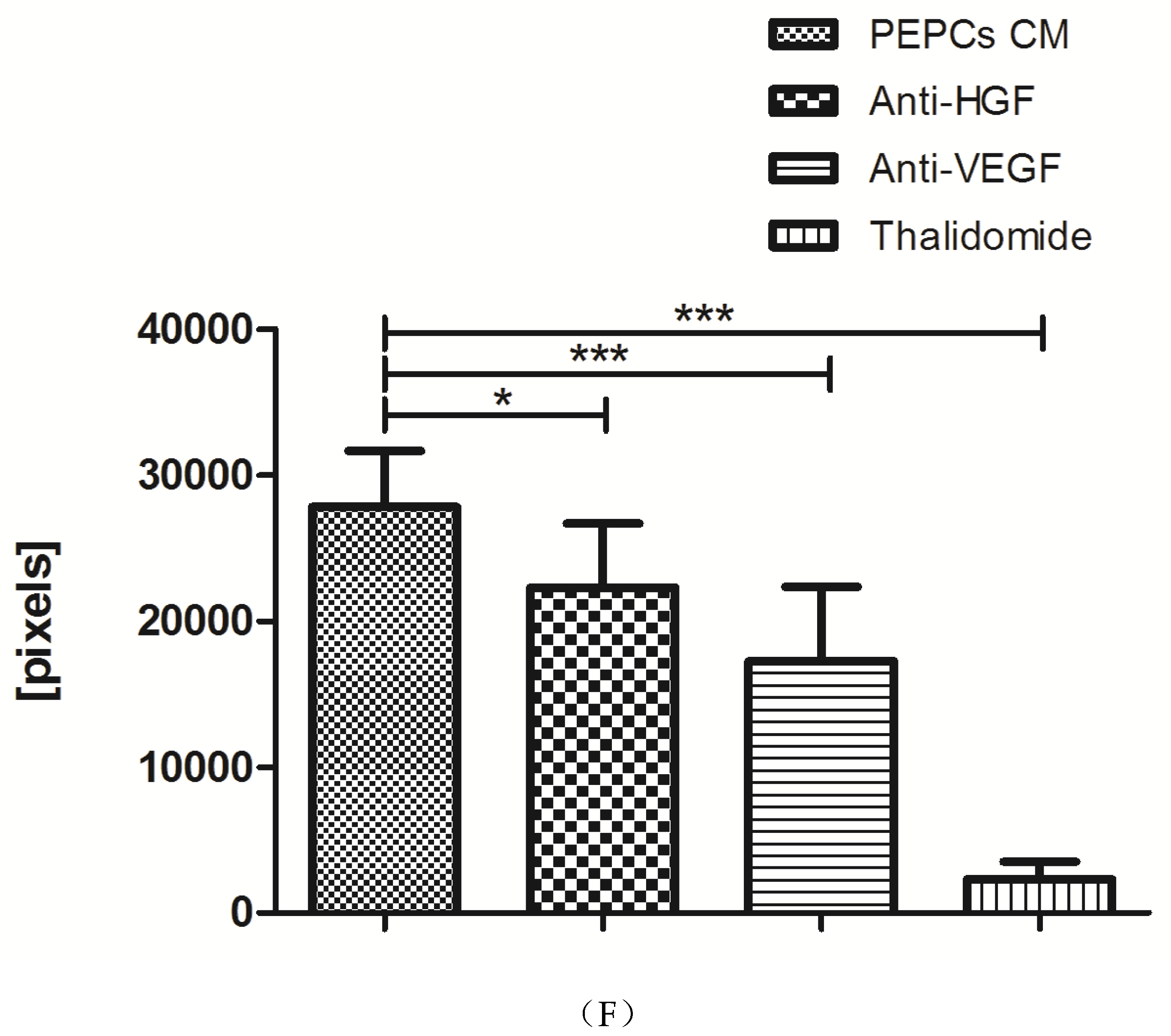
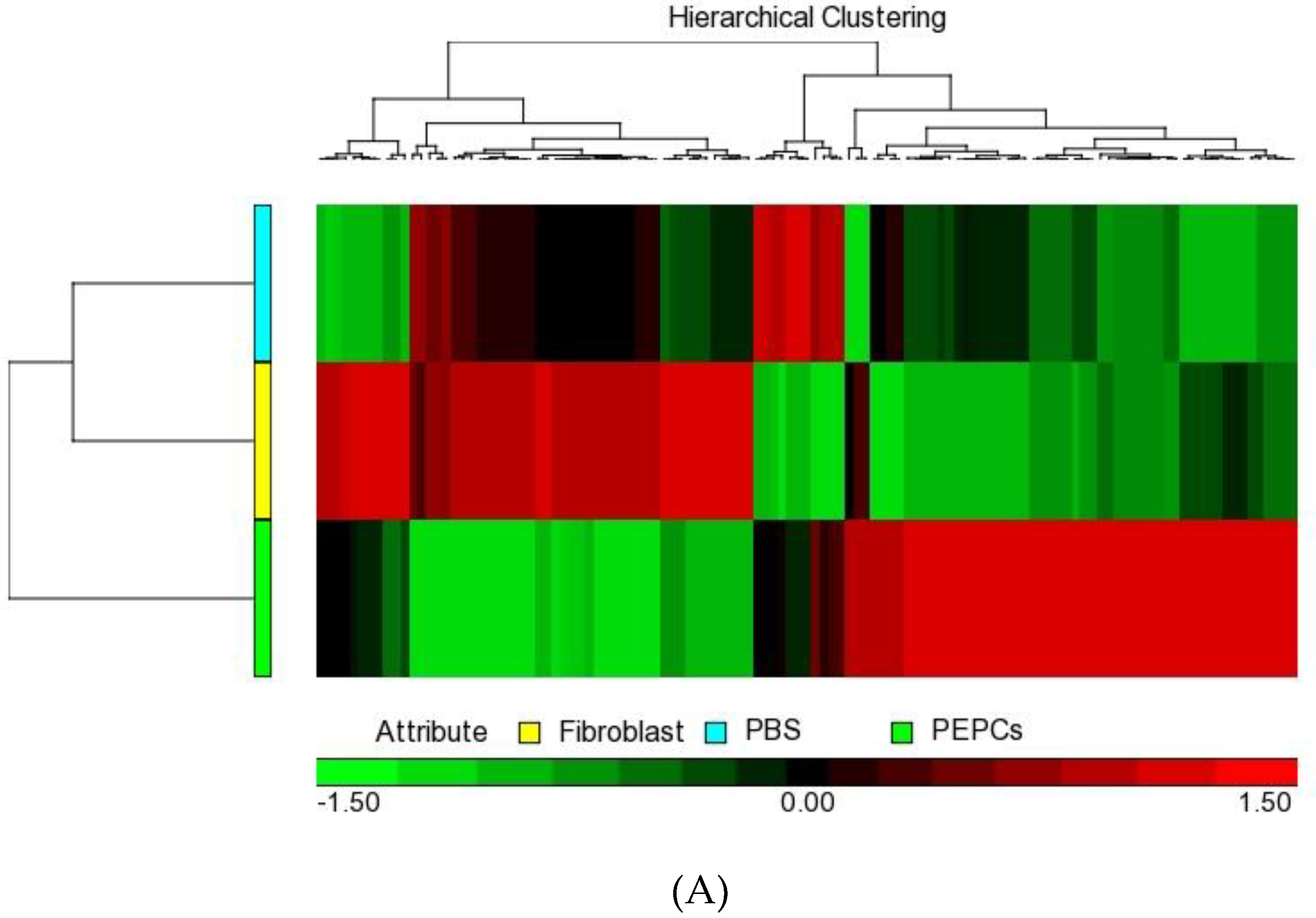
3.5. Paracrine Effects of PEPCs on Ischemic Heart Repair
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; de Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics-2017 Update: A Report from the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef] [PubMed]
- Zheleva, B.; Atwood, J.B. The invisible child: Childhood heart disease in global health. Lancet 2017, 389, 16–18. [Google Scholar] [CrossRef]
- Wren, C.; Irving, C.A.; Griffiths, J.A.; O’Sullivan, J.J.; Chaudhari, M.P.; Haynes, S.R.; Smith, J.H.; Hamilton, J.R.; Hasan, A. Mortality in infants with cardiovascular malformations. Eur. J. Pediatr. 2012, 171, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Oster, M.E.; Lee, K.A.; Honein, M.A.; Riehle-Colarusso, T.; Shin, M.; Correa, A. Temporal trends in survival among infants with critical congenital heart defects. Pediatrics 2013, 131, e1502–e1508. [Google Scholar] [CrossRef] [PubMed]
- Erikssen, G.; Liestol, K.; Seem, E.; Birkeland, S.; Saatvedt, K.J.; Hoel, T.N.; Dohlen, G.; Skulstad, H.; Svennevig, J.L.; Thaulow, E.; et al. Achievements in congenital heart defect surgery: A prospective, 40-year study of 7038 patients. Circulation 2015, 131, 337–346; discussion 346. [Google Scholar] [CrossRef] [PubMed]
- Lui, G.K.; Fernandes, S.; McElhinney, D.B. Management of cardiovascular risk factors in adults with congenital heart disease. J. Am. Heart Assoc. 2014, 3, e001076. [Google Scholar] [CrossRef] [PubMed]
- Behfar, A.; Crespo-Diaz, R.; Terzic, A.; Gersh, B.J. Cell therapy for cardiac repair—Lessons from clinical trials. Nat. Rev. Cardiol. 2014, 11, 232–246. [Google Scholar] [CrossRef] [PubMed]
- Mathur, A.; Fernandez-Aviles, F.; Dimmeler, S.; Hauskeller, C.; Janssens, S.; Menasche, P.; Wojakowski, W.; Martin, J.F.; Zeiher, A.; Investigators, B. The consensus of the Task Force of the European Society of Cardiology concerning the clinical investigation of the use of autologous adult stem cells for the treatment of acute myocardial infarction and heart failure: Update 2016. Eur. Heart J. 2017. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.J.; Yang, X.; Don, C.W.; Minami, E.; Liu, Y.W.; Weyers, J.J.; Mahoney, W.M.; Van Biber, B.; Cook, S.M.; Palpant, N.J.; et al. Human embryonic-stem-cell-derived cardiomyocytes regenerate non-human primate hearts. Nature 2014, 510, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Fisher, S.A.; Brunskill, S.J.; Doree, C.; Mathur, A.; Taggart, D.P.; Martin-Rendon, E. Stem cell therapy for chronic ischaemic heart disease and congestive heart failure. Cochrane Database Syst. Rev. 2014, CD007888. [Google Scholar] [CrossRef]
- Nowbar, A.N.; Mielewczik, M.; Karavassilis, M.; Dehbi, H.M.; Shun-Shin, M.J.; Jones, S.; Howard, J.P.; Cole, G.D.; Francis, D.P.; Group, D.W. Discrepancies in autologous bone marrow stem cell trials and enhancement of ejection fraction (DAMASCENE): Weighted regression and meta-analysis. BMJ 2014, 348, g2688. [Google Scholar] [CrossRef] [PubMed]
- Abbott, A. Doubts over heart stem-cell therapy. Nature 2014, 509, 15–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dykstra, J.A.; Facile, T.; Patrick, R.J.; Francis, K.R.; Milanovich, S.; Weimer, J.M.; Kota, D.J. Concise Review: Fat and Furious: Harnessing the Full Potential of Adipose-Derived Stromal Vascular Fraction. Stem Cells Transl. Med. 2017, 6, 1096–1108. [Google Scholar] [CrossRef] [PubMed]
- Roura, S.; Pujal, J.M.; Galvez-Monton, C.; Bayes-Genis, A. The role and potential of umbilical cord blood in an era of new therapies: A review. Stem Cell Res. Ther. 2015, 6, 123. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.R.; Barile, L.; Cho, H.C.; Leppo, M.K.; Hare, J.M.; Messina, E.; Giacomello, A.; Abraham, M.R.; Marban, E. Regenerative potential of cardiosphere-derived cells expanded from percutaneous endomyocardial biopsy specimens. Circulation 2007, 115, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Cambria, E.; Pasqualini, F.S.; Wolint, P.; Gunter, J.; Steiger, J.; Bopp, A.; Hoerstrup, S.P.; Emmert, M.Y. Translational cardiac stem cell therapy: Advancing from first-generation to next-generation cell types. NPJ Regen. Med. 2017, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Cheung, E.W.; Ho, S.A.; Tang, K.K.; Chau, A.K.; Chiu, C.S.; Cheung, Y.F. Pericardial effusion after open-heart surgery for congenital heart disease. Heart 2003, 89, 780–783. [Google Scholar] [CrossRef]
- Zudaire, E.; Gambardella, L.; Kurcz, C.; Vermeren, S. A computational tool for quantitative analysis of vascular networks. PLoS ONE 2011, 6, e27385. [Google Scholar] [CrossRef]
- Li, Z.; Wu, J.C.; Sheikh, A.Y.; Kraft, D.; Cao, F.; Xie, X.; Patel, M.; Gambhir, S.S.; Robbins, R.C.; Cooke, J.P. Differentiation, survival, and function of embryonic stem cell derived endothelial cells for ischemic heart disease. Circulation 2007, 116, I46–I54. [Google Scholar] [CrossRef]
- Nelson, D.L.; Ledbetter, S.A.; Corbo, L.; Victoria, M.F.; Ramirez-Solis, R.; Webster, T.D.; Ledbetter, D.H.; Caskey, C.T. Alu polymerase chain reaction: A method for rapid isolation of human-specific sequences from complex DNA sources. Proc. Natl. Acad. Sci. USA 1989, 86, 6686–6690. [Google Scholar] [CrossRef]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukho, P.; Kirpensteijn, J.; Hesselink, J.W.; van Osch, G.J.; Verseijden, F.; Bastiaansen-Jenniskens, Y.M. Effect of Cell Seeding Density and Inflammatory Cytokines on Adipose Tissue-Derived Stem Cells: An in Vitro Study. Stem Cell Rev. 2017, 13, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Gude, N.A.; Sussman, M.A. Chasing c-Kit through the heart: Taking a broader view. Pharmacol. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Czapla, J.; Matuszczak, S.; Wisniewska, E.; Jarosz-Biej, M.; Smolarczyk, R.; Cichon, T.; Glowala-Kosinska, M.; Sliwka, J.; Garbacz, M.; Szczypior, M.; et al. Human Cardiac Mesenchymal Stromal Cells with CD105+CD34- Phenotype Enhance the Function of Post-Infarction Heart in Mice. PLoS ONE 2016, 11, e0158745. [Google Scholar] [CrossRef] [PubMed]
- Britz-Cunningham, S.H.; Shah, M.M.; Zuppan, C.W.; Fletcher, W.H. Mutations of the Connexin43 gap-junction gene in patients with heart malformations and defects of laterality. N. Engl. J. Med. 1995, 332, 1323–1329. [Google Scholar] [CrossRef]
- Keck, M.; van Dijk, R.M.; Deeg, C.A.; Kistler, K.; Walker, A.; von Ruden, E.L.; Russmann, V.; Hauck, S.M.; Potschka, H. Proteomic profiling of epileptogenesis in a rat model: Focus on cell stress, extracellular matrix and angiogenesis. Neurobiol. Dis. 2018, 112, 119–135. [Google Scholar] [CrossRef]
- Yagi, H.; Soto-Gutierrez, A.; Parekkadan, B.; Kitagawa, Y.; Tompkins, R.G.; Kobayashi, N.; Yarmush, M.L. Mesenchymal stem cells: Mechanisms of immunomodulation and homing. Cell Transplant. 2010, 19, 667–679. [Google Scholar] [CrossRef]
- Agarwal, U.; Smith, A.W.; French, K.M.; Boopathy, A.V.; George, A.; Trac, D.; Brown, M.E.; Shen, M.; Jiang, R.; Fernandez, J.D.; et al. Age-Dependent Effect of Pediatric Cardiac Progenitor Cells After Juvenile Heart Failure. Stem Cells Transl. Med. 2016, 5, 883–892. [Google Scholar] [CrossRef]
- Marban, E. Breakthroughs in cell therapy for heart disease: Focus on cardiosphere-derived cells. Mayo Clin. Proc. 2014, 89, 850–858. [Google Scholar] [CrossRef]
- Privratsky, J.R.; Newman, D.K.; Newman, P.J. PECAM-1: Conflicts of interest in inflammation. Life Sci. 2010, 87, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Sidney, L.E.; Branch, M.J.; Dunphy, S.E.; Dua, H.S.; Hopkinson, A. Concise review: Evidence for CD34 as a common marker for diverse progenitors. Stem Cells 2014, 32, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, A.; Sen, D. Mesenchymal stem cells in cardiac regeneration: A detailed progress report of the last 6 years (2010–2015). Stem Cell Res. Ther. 2016, 7, 82. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.J.; Chandrakanthan, V.; Xaymardan, M.; Asli, N.S.; Li, J.; Ahmed, I.; Heffernan, C.; Menon, M.K.; Scarlett, C.J.; Rashidianfar, A.; et al. Adult cardiac-resident MSC-like stem cells with a proepicardial origin. Cell Stem Cell 2011, 9, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.E.; Garitaonandia, I.; Loring, J.F. The tumorigenic potential of pluripotent stem cells: What can we do to minimize it? Bioessays 2016, 38 (Suppl. 1), S86–S95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.S.; Tang, C.; Rao, M.S.; Weissman, I.L.; Wu, J.C. Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat. Med. 2013, 19, 998–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, K.; Ibrahim, A.; Hensley, M.T.; Shen, D.; Sun, B.; Middleton, R.; Liu, W.; Smith, R.R.; Marban, E. Relative roles of CD90 and c-kit to the regenerative efficacy of cardiosphere-derived cells in humans and in a mouse model of myocardial infarction. J. Am. Heart Assoc. 2014, 3, e001260. [Google Scholar] [CrossRef] [PubMed]
- Smits, A.M.; van Vliet, P.; Metz, C.H.; Korfage, T.; Sluijter, J.P.; Doevendans, P.A.; Goumans, M.J. Human cardiomyocyte progenitor cells differentiate into functional mature cardiomyocytes: An in vitro model for studying human cardiac physiology and pathophysiology. Nat. Protoc. 2009, 4, 232–243. [Google Scholar] [CrossRef]
- Rupp, S.; Badorff, C.; Koyanagi, M.; Urbich, C.; Fichtlscherer, S.; Aicher, A.; Zeiher, A.M.; Dimmeler, S. Statin therapy in patients with coronary artery disease improves the impaired endothelial progenitor cell differentiation into cardiomyogenic cells. Basic Res. Cardiol. 2004, 99, 61–68. [Google Scholar] [CrossRef]
- Gnecchi, M.; Zhang, Z.; Ni, A.; Dzau, V.J. Paracrine mechanisms in adult stem cell signaling and therapy. Circ. Res. 2008, 103, 1204–1219. [Google Scholar] [CrossRef]
- Korf-Klingebiel, M.; Kempf, T.; Sauer, T.; Brinkmann, E.; Fischer, P.; Meyer, G.P.; Ganser, A.; Drexler, H.; Wollert, K.C. Bone marrow cells are a rich source of growth factors and cytokines: Implications for cell therapy trials after myocardial infarction. Eur. Heart J. 2008, 29, 2851–2858. [Google Scholar] [CrossRef]
- Gallet, R.; de Couto, G.; Simsolo, E.; Valle, J.; Sun, B.; Liu, W.; Tseliou, E.; Zile, M.R.; Marban, E. Cardiosphere-derived cells reverse heart failure with preserved ejection fraction (HFpEF) in rats by decreasing fibrosis and inflammation. JACC Basic Transl. Sci. 2016, 1, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Marban, E.; Cheng, K. Heart to heart: The elusive mechanism of cell therapy. Circulation 2010, 121, 1981–1984. [Google Scholar] [CrossRef] [PubMed]
- Gallet, R.; Dawkins, J.; Valle, J.; Simsolo, E.; de Couto, G.; Middleton, R.; Tseliou, E.; Luthringer, D.; Kreke, M.; Smith, R.R.; et al. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur. Heart J. 2017, 38, 201–211. [Google Scholar] [CrossRef] [PubMed]













© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, T.-W.; Kan, C.-D.; Chiu, W.-T.; Fok, K.L.; Ruan, Y.C.; Jiang, X.; Chen, J.; Kao, C.-C.; Chen, I.-Y.; Lin, H.-C.; et al. Progenitor Cells Derived from Drain Waste Product of Open-Heart Surgery in Children. J. Clin. Med. 2019, 8, 1028. https://doi.org/10.3390/jcm8071028
Wong T-W, Kan C-D, Chiu W-T, Fok KL, Ruan YC, Jiang X, Chen J, Kao C-C, Chen I-Y, Lin H-C, et al. Progenitor Cells Derived from Drain Waste Product of Open-Heart Surgery in Children. Journal of Clinical Medicine. 2019; 8(7):1028. https://doi.org/10.3390/jcm8071028
Chicago/Turabian StyleWong, Tak-Wah, Chung-Dann Kan, Wen-Tai Chiu, Kin Lam Fok, Ye Chun Ruan, Xiaohua Jiang, Junjiang Chen, Chiu-Ching Kao, I-Yu Chen, Hui-Chun Lin, and et al. 2019. "Progenitor Cells Derived from Drain Waste Product of Open-Heart Surgery in Children" Journal of Clinical Medicine 8, no. 7: 1028. https://doi.org/10.3390/jcm8071028