Determinants of Termite Assemblages’ Characteristics within Natural Habitats of a Sudano-Guinean Savanna (Comoe National Park, Côte d’Ivoire)

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Site and Visited Habitats
2.2. Sampling Design
2.2.1. Assessing Termite Diversity and Abundance
2.2.2. Environmental Variables Characterization
2.2.3. Identification of Specimens
2.3. Data Analysis
3. Results
3.1. Overall Taxonomic Structure of the Observed Species
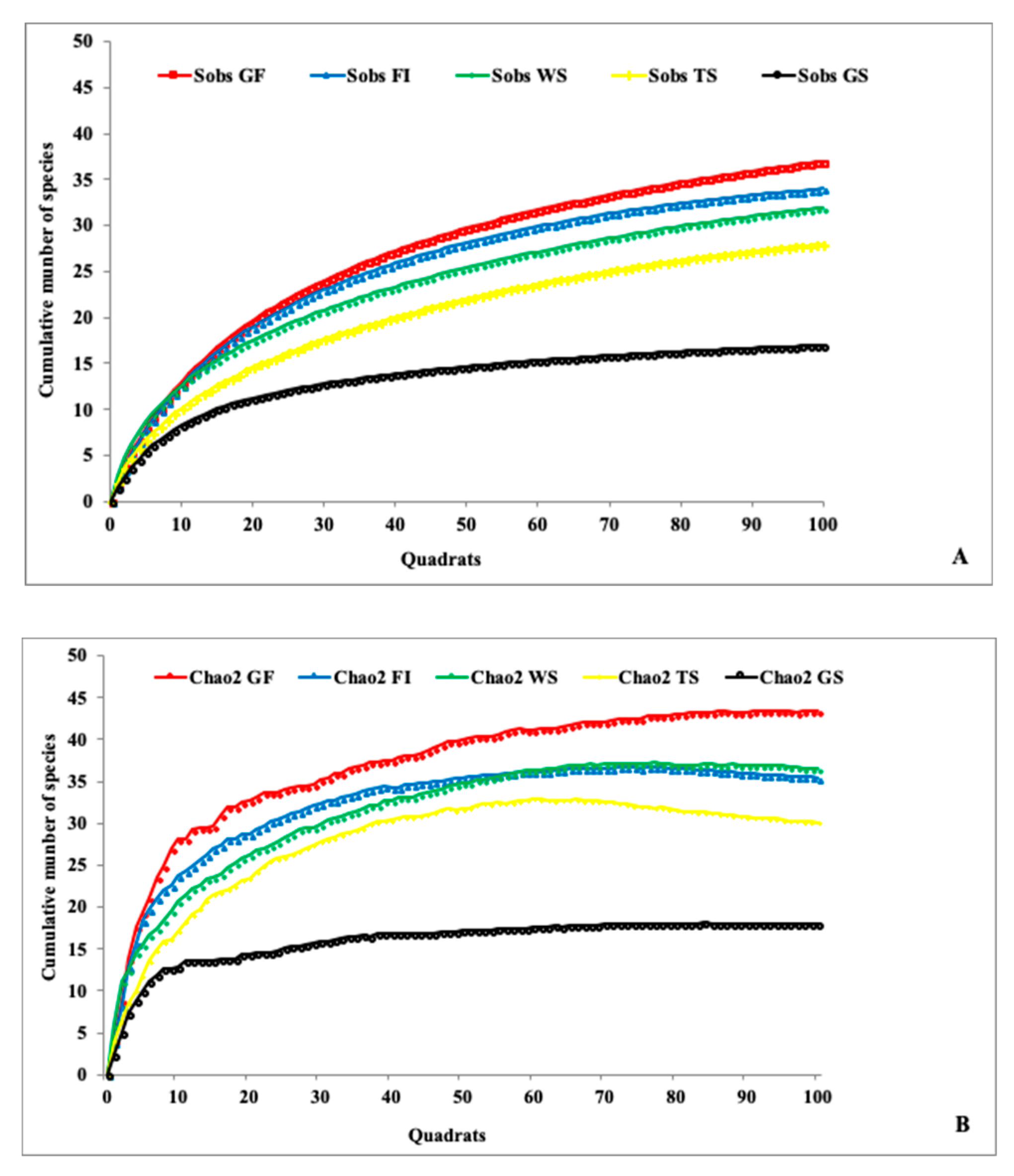
3.2. Sampling Efficiency during this Study
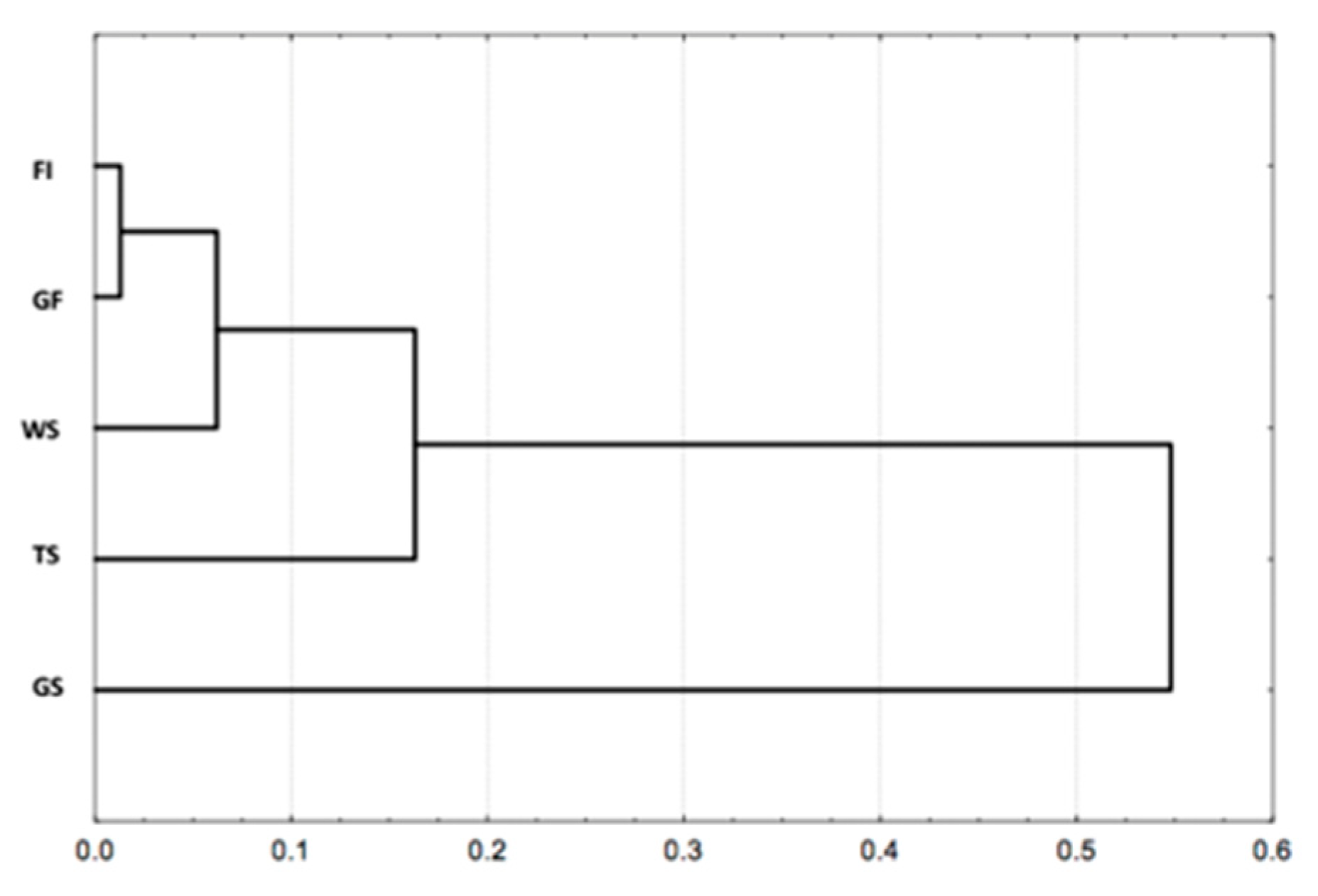
3.3. Species Richness, Species Diversity, and Spatial Distribution of the Recorded Termites
3.4. Relative Abundance of Termite’s Feeding Groups across the Visited Habitats
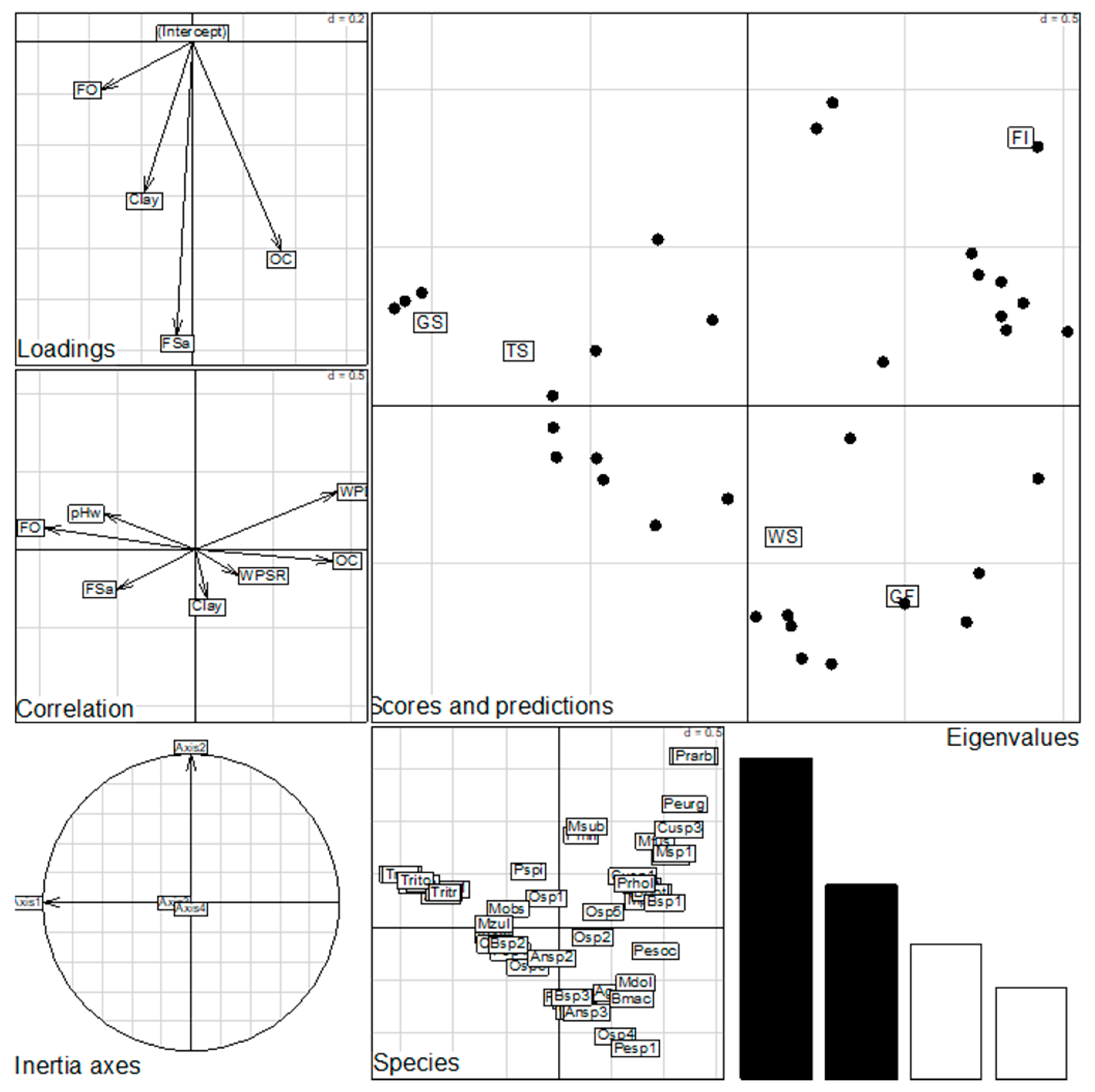
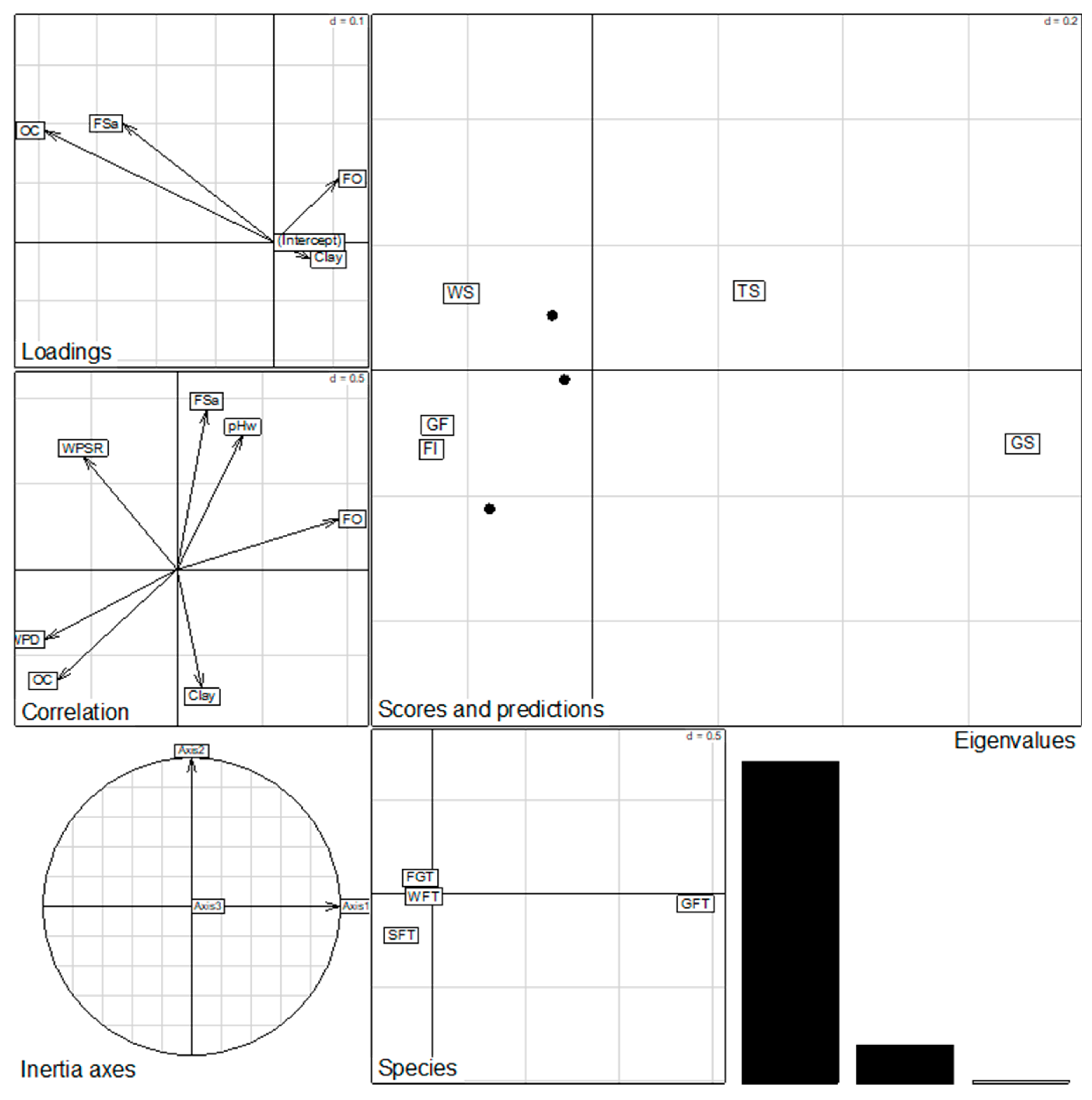
3.5. Effects of Environmental Variables on Species Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Termites Species (Feeding Group/Family/Subfamily) | Habitats | ||||
|---|---|---|---|---|---|
| FI | GF | WS | TS | GS | |
| Fungus growers—Termitidae—Macrotermitinae | |||||
| Ancistrotermes cavithorax (Sjöstedt, 1899) | 2 | 15 | 39 | 62 | 29 |
| Ancistrotermes crucifer (Sjöstedt, 1897) | 26 | 10 | 19 | 0 | 0 |
| Ancistrotermes guineensis (Silvestri, 1912) | 14 | 45 | 56 | 1 | 1 |
| Macrotermes bellicosus (Smeathman, 1781) | 7 | 4 | 2 | 1 | 0 |
| Macrotermes subhyalinus (Rambur, 1842) | 12 | 0 | 2 | 5 | 3 |
| Microtermes osborni (Emerson, 1928) | 8 | 3 | 16 | 30 | 10 |
| Microtermes toumodiensis (Grassé, 1937) | 0 | 4 | 0 | 0 | 0 |
| Pseudacanthotermes militaris (Hagen, 1858) | 11 | 0 | 2 | 9 | 0 |
| Pseudacanthotermes spiniger (Sjöstedt, 1900) | 4 | 0 | 2 | 9 | 0 |
| Pseudacanthotermes sp1 | 0 | 0 | 2 | 2 | 0 |
| Odontotermes sp1 | 10 | 9 | 4 | 2 | 15 |
| Odontotermes sp2 | 3 | 5 | 2 | 2 | 1 |
| Odontotermes sp3 | 0 | 8 | 4 | 4 | 0 |
| Odontotermes sp4 | 0 | 4 | 2 | 0 | 0 |
| Odontotermes sp5 | 2 | 1 | 2 | 1 | 0 |
| Odontotermes sp6 | 0 | 3 | 2 | 2 | 2 |
| Odontotermes sp7 | 0 | 2 | 0 | 0 | 0 |
| Odontotermes sp8 | 0 | 0 | 1 | 0 | 0 |
| Wood feeders—Rhinotermitidae—Coptotermitinae | |||||
| Coptotermes intermedius (Sylvestri, 1912) | 0 | 6 | 0 | 0 | 0 |
| Coptotermes sjoestedti (Harris, 1966) | 0 | 2 | 0 | 0 | 3 |
| Wood feeders—Termitidae—Nasutitermitinae | |||||
| Fulleritermes tenebricus (Silvestri, 1914) | 0 | 0 | 23 | 3 | 0 |
| Nasutitermes arborum (Smeathman, 1781) | 13 | 6 | 1 | 0 | 0 |
| Wood feeders—Rhinotermitidae—Rhinotermitinae | |||||
| Shedorhinotermes laminianus (Sjöstedt, 1911) | 2 | 0 | 2 | 0 | 0 |
| Wood feeders—Termitidae—Termitinae | |||||
| Amitermes evuncifer (Silvestri, 1912) | 0 | 1 | 0 | 13 | 6 |
| Amitermes lonnbergianus (Sjöstedt, 1911) | 0 | 1 | 0 | 0 | 0 |
| Amitermes sp | 0 | 0 | 1 | 0 | 0 |
| Microcerotermes choanensis (Silvestri, 1914) | 1 | 0 | 0 | 0 | 0 |
| Microcerotermes dolichognathus (Sjöstedt, 1926) | 1 | 3 | 1 | 0 | 0 |
| Microcerotermes fuscotibialis (Sjöstedt, 1896) | 2 | 0 | 1 | 0 | 0 |
| Microcerotermes parvulus (Sjöstedt, 1911) | 2 | 1 | 1 | 0 | 0 |
| Microcerotermes parvus (Haviland, 1898) | 17 | 11 | 8 | 1 | 0 |
| Microcerotermes zuluensis (Sjöstedt, 1926) | 3 | 7 | 14 | 24 | 17 |
| Microcerotermes sp1 | 2 | 1 | 0 | 0 | 0 |
| Microcerotermes sp2 | 0 | 0 | 0 | 0 | 1 |
| Termes sp1 | 0 | 1 | 0 | 0 | 0 |
| Soil feeders—Termitidae—Apicotermitinae | |||||
| Adaiphrotermes sp | 1 | 0 | 2 | 10 | 9 |
| Aderitotermes sp | 7 | 21 | 28 | 11 | 15 |
| Astratotermes sp | 0 | 7 | 6 | 2 | 0 |
| Soil feeders—Termitidae—Cubitermitinae | |||||
| Basidentitermes potens (Silvestri, 1914) | 2 | 1 | 1 | 0 | 0 |
| Basidentitermes mactus (Silvestri, 1914) | 2 | 9 | 3 | 0 | 0 |
| Basidentitermes sp1 | 1 | 1 | 0 | 0 | 0 |
| Basidentitermes sp2 | 0 | 1 | 0 | 2 | 0 |
| Basidentitermes sp3 | 0 | 4 | 0 | 2 | 0 |
| Cubitermes sp1 | 3 | 2 | 0 | 1 | 0 |
| Cubitermes sp2 | 3 | 0 | 0 | 0 | 0 |
| Cubitermes sp3 | 6 | 2 | 0 | 0 | 0 |
| Soil feeders—Termitidae—Termitinae | |||||
| Pericapritermes appelans (Silvestri, 1914) | 0 | 2 | 0 | 0 | 0 |
| Pericapritermes silvestrianus (Emerson, 1928) | 1 | 0 | 0 | 0 | 0 |
| Pericapritermes socialis (Sjöstedt 1965) | 1 | 2 | 0 | 0 | 0 |
| Pericapritermes urgens (Silvestri 1914) | 5 | 1 | 0 | 0 | 0 |
| Pericapritermes sp1 | 0 | 3 | 0 | 0 | 0 |
| Procubitermes aburiensis (Sjöstedt, 1926) | 5 | 0 | 0 | 0 | 0 |
| Promirotermes holmgreni (Silvestri, 1912) | 2 | 0 | 2 | 0 | 0 |
| Procubitermes sjoestedti (Rosen, 1912) | 6 | 0 | 0 | 0 | 0 |
| Grass feeders—Termitidae—Nasutitermitinae | |||||
| Trinervitermes geminatus (Wasmann, 1897) | 0 | 0 | 1 | 1 | 12 |
| Trinervitermes occidentalis (Sjöstedt, 1904) | 0 | 0 | 0 | 0 | 11 |
| Trinervitermes oeconomus (Trägårdh, 1904) | 0 | 0 | 0 | 4 | 0 |
| Trinervitermes togoensis (Sjöstedt, 1899) | 0 | 0 | 0 | 2 | 4 |
| Trinervitermes rhodesiensis (Sjöstedt, 1911) | 0 | 0 | 0 | 2 | 2 |
| Trinervitermes sp1 | 0 | 0 | 0 | 1 | 0 |
References
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Lavelle, P. Faunal Activities and Soil Processes: Adaptive Strategies That Determine Ecosystem Function. Adv. Ecol. Res. 1997, 27, 93–132. [Google Scholar] [Green Version]
- Dangerfield, J.M.; McCarthy, T.S.; Ellery, W.N. The mound-building termite Macrotermes michaelseni as an ecosystem engineer. J. Trop. Ecol. 1998, 14, 507–520. [Google Scholar] [CrossRef]
- Bignell, D.E.; Eggleton, P. Termites in ecosystems. In Termites: Evolution, Sociality, Symbiose, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 363–387. [Google Scholar]
- Jouquet, P.; Blanchart, E.; Capowiezc, Y. Utilization of earthworms and termites for the restoration of ecosystem functioning. Appl. Soil Ecol. 2014, 73, 34–40. [Google Scholar] [CrossRef]
- Lobry de Bruyn, L.A.; Cornacher, A.J. The role of termites and ants in soil modification: A review. Aust. J. Soil Res. 1990, 28, 55–93. [Google Scholar]
- Sileshi, G.W.; Arshad, M.A.; Konaté, S.; Nkunika, P.O.Y. Termite-induced heterogeneity in African savanna vegetation: Mechanisms and patterns. J. Veg. Sci. 2010, 21, 923–937. [Google Scholar] [CrossRef]
- Collins, N.M. The role termites in the decomposition of wood and leaf litter in the southern Guinea Savanna of Nigeria. Oecologia 1981, 51, 389–399. [Google Scholar]
- Davies, R.G. Termite species richness in fire-prone and fire protected dry deciduous dipterocarp forest in Doi Sthep-Pui National Park, Northern Thailand. J. Trop. Ecol. 1997, 13, 153–160. [Google Scholar] [CrossRef]
- Eggleton, P.; Bignell, D.E.; Hauser, S.; Dibog, L.; Norgrove, L.; Madong, B. Termite diversity across an anthropogenic gradient in the humid forest zone of west Africa. Agric. Ecosyst. Environ. 2002, 90, 189–202. [Google Scholar] [CrossRef]
- Konaté, S.; Yéo, K.; Yeboue, L.; Alonso, L.F.; Kouassi, K. Evaluation rapide de la diversité des insectes des forêts classées de la Haute Dodo et du Cavally (Côte-d’Ivoire). In RAP Bulletin of Biological Assessment; Conservation International (CI): Washington, DC, USA, 2005; 27p. [Google Scholar]
- Venter, F.J.; Scholes, R.J.; Eckhardt, H.C. The abiotic template and its associated vegetation pattern. In The Kruger Experience Ecology and Management of Savanna Heterogeneity; du Toit, J., Rogers, K., Biggs, H., Eds.; Island Press: Washington, DC, USA, 2003; pp. 83–129. [Google Scholar]
- Baldeck, C.A.; Harms, K.E.; Yavitt, J.B.; John, R.; Turner, B.L.; Valencia, R.; Navarrete, H.; Davies, S.J.; Chuyong, G.B.; Kenfack, D.; et al. Soil resources and topography shape local tree community structure in tropical forests. Proc. R. Soc. Lond. B Biol. Sci. 2013, 280, 1753. [Google Scholar] [CrossRef]
- Sankaran, M.; Hanan, N.P.; Scholes, R.J.; Ratnam, J.; Augustine, D.J.; Cade, B.S.; Gignoux, J.; Higgins, S.I.; Le Roux, X.; Ludwig, F.; et al. Determinants of woody cover in African Savannas. Nature 2005, 438, 846–849. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.E.; Wood, T.G. Abundance of mounds and competition among colonies of some Australian termite species. Pedobiologia 1971, 11, 341–366. [Google Scholar]
- Dosso, K.; Konaté, S.; Aidara, D.; Linsenmair, K.E. Termite diversity and Abundance across fire-induced habitat variability in a tropical moist savanna (Lamto, Central Côte d’Ivoire). J. Trop. Ecol. 2010, 26, 323–334. [Google Scholar] [CrossRef]
- Koffi, P. Le parc National de la Comoé. In Atlas de la Biodiversité de L’Afrique de L’Ouest—Tome III: Côte d’Ivoire; Konaté, S., Kampmann, D., Eds.; BIOTA: Abidjan, Côte d’Ivoire & Frankfort/Main, Germany, 2010; pp. 272–274. ISBN 978-3-9813933-2-3. [Google Scholar]
- Eldin, M. Le climat de la CÔte d’Ivoire. In Le Milieu Naturel de Côte d’Ivoire; Avenard, J.-M., Eldin, M., Girard, G., Sircoulon, J., Touchebeuf, P., Guillaumet, J.-L., Adjanohoun, E., Eds.; Mémoires Orstom No. 50; ORSTOM: Paris, France, 1971; pp. 73–108. [Google Scholar]
- Guillaumet, J.-L.; Adjanohoun, E. La végétation. In Le Milieu Naturel de Côte d’Ivoire; Avenard, J.-M., Eldin, M., Girard, G., Sircoulon, J., Touchebeuf, P., Guillaumet, J.-L., Adjanohoun, E., Eds.; Mémoires Orstom No. 50; ORSTOM: Paris, France, 1971; pp. 161–263. [Google Scholar]
- Hennenberg, K.J.; Goetze, D.; Kouamé, L.; Orthmann, B.; Porembski, S. Border and ecotone detection by vegetation composition along forest-savanna transects in Ivory Coast. J. Veg. Sci. 2005, 16, 301–310. [Google Scholar] [CrossRef]
- Poilecot, P.; Bonfou, K.; Dosso, H.; Laugenie, F.; N’dri, K.; Nicole, M.; Sangaré, Y. Un Écosystème de Savane Soudanienne: Le Parc National de la Comoé (Côte d’Ivoire); UNESCO: Paris, France, 1991; 346p. [Google Scholar]
- Gautier, L. Contact forêt-savane en Côte d’Ivoire centrale: Evolution du recouvrement ligneux des savanes de la réserve de Lamto (sud du V Baoulé). Candollea 1990, 45, 627–641. [Google Scholar]
- Jones, D.T.; Eggleton, P. Sampling termite assemblage in tropical forest: Testing a rapid biodiversity assessment protocol. J. Appl. Ecol. 2000, 37, 191–203. [Google Scholar] [CrossRef]
- Hausberger, B.; Kimpel, D.; van Neer, A.; Korb, J. Uncovering cryptic species diversity of a community in a West African savanna. Mol. Phylogenet. Evol. 2011, 61, 964–969. [Google Scholar] [CrossRef]
- Koné, N.A.; Dosso, K.; Konaté, S.; Kouadio, Y.J; Linsenmair, K.E. Environmental and biological determinants of Termitomyces species seasonal fructification in central and southern Côte d’Ivoire. Insectes Sociaux 2011, 58, 371–382. [Google Scholar] [CrossRef]
- Bouillon, A.; Mathot, G. Quel est ce Termite Africain? Université de Leopoldville: Leopoldville, Congo, 1965; 115p. [Google Scholar]
- Bouillon, A.; Mathot, G. Quel est ce Termite Africain? Supplement No. 1; Université Lovanium: Kinshassa, Congo, 1966; 23p. [Google Scholar]
- Bouillon, A.; Mathot, G. Quel est ce Termite Africain? Supplement No. 2; Université Nationale du Zaıre: Kinshassa, Congo, 1971; 48p. [Google Scholar]
- Webb, G.C. Keys of the Genera of the African Termites Adapted from Revision Der Termiten Afrikas of Sjoestedt; Ibadan University Press: Ibadan, Nigeria, 1961; 35p. [Google Scholar]
- Josens, G. Etudes Biologiques et Écologiques des Termites (Isoptera) de la Savane de Lamto. Ph.D. Thesis, L’Universite Libre de Bruxelles, Bruxelles, Belgium, 1972; 262p. [Google Scholar]
- Grassé, P.P. Termitologia, Tome III: Comportement, Socialite, Ecologie, Evolution, Systematique; Masson: Paris, France, 1986; 676p. [Google Scholar]
- Grassé, P.P. Termitologia, Tome II: Fondation des Sociétés et Construction; Masson: Paris, France, 1984; 613p. [Google Scholar]
- Konaté, S. Structure Dynamique et Rôle des Buttes Termitiques Dans le Fonctionnement D’une Savane Préforestière (Lamto, Côte d’Ivoire): Le Termite Champignonniste Odontotermes Comme Ingénieur de L’écosystème. Ph.D. Thesis, L’Université Paris 6, Paris, France, 1998; 252p. [Google Scholar]
- Cao, Y.; Williams, D.D.; Larsen, P.D. Comparison of ecological communities: The problem of sample representativeness. Ecol. Monogr. 2002, 72, 313–318. [Google Scholar] [CrossRef]
- Colwell, R.K.; Coddington, J.A. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. Lond. B 1994, 345, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Morin, A.; Findlay, S. Biodiversité: Tendance et Processus—Biologie de la Conservation des Espèces; Université d’Ottawa: Ottawa, ON, Canada, 2001; 25p. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science Ltd.: Oxford, UK, 2004; 256p. [Google Scholar]
- Vane-Wright, R.I.; Humphries, C.J.; Williams, P.H. What to protect? Systematics and the agony of choice. Biol. Conserv. 1991, 55, 235–254. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Palmer, M.W. Putting things in even better order: The advantages of canonical correspondence analysis. Ecology 1983, 74, 2215–2230. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F. Canonical correspondence analysis. A new eigenvector technique formultivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F. The analysis of vegetation—Environment relationships by canonical correspondence analysis. Vegetation 1987, 69, 69–77. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F. Canoco—A Fortran Program for Canonical Community Ordination by (Partial) Detrended (Canonical) Correspondence Analysis Principal Components Analysis and Redundancy Analysis (Version 2.1); Agricultural Mathematics Group: Wageningen, The Netherlands, 1988. [Google Scholar]
- Lebreton, J.D.; Chessel, D.; Richardot-Coolet, M.; Yoccoz, N. L’analyse des relations espèces—Milieu par l’analyse canonique des correspondences. I. variable de milieu quantitative. Acta Oecol. 1988, 9, 53–67. [Google Scholar]
- Jongman, R.H.G.; Ter Braak, C.J.K.; van Tongeren, O.F.R. Data Analysis in Community and Landscape Ecology, 2nd ed.; Cambridge University Press: New York, NY, USA, 1995; 299p. [Google Scholar]
- Farji-Brener, A.G.; Corley, J.C.; Bettinelli, J. The effects of fire on ant communities in north-western Patagonia: The importance of habitat structure and regional context. Divers. Distrib. 2002, 8, 235–243. [Google Scholar] [CrossRef]
- Parr, C.L.; Bond, W.L.; Robertson, H. A preliminary study of the effect of fire on ants (Hymenoptera: Formicidae) in a South African savanna. Afr. Entomol. 2002, 10, 101–111. [Google Scholar]
- Hoffmann, B.D. Responses of ant communities to experimental fire regimes on rangelands in the Victoria River District of the Northern Territory. Austral Ecol. 2003, 28, 182–195. [Google Scholar] [CrossRef]
- Williams, R.J.; Gill, A.M.; Moore, P.H.R. Seasonal changes in fire behaviour in a tropical savanna in Northern Australia. Int. J. Wildland Fire 1998, 8, 227–239. [Google Scholar] [CrossRef]
- Savadogo, D.D.; Savadogo, P.; Zida, D.; Sawadogo, L.; Tiveau, D.; Oden, P.C. Fuel and fire characteristics in savanna-woodland of west Africa in relation to grazing and dominant grass type. Int. J. Wildland Fire 2007, 16, 531–539. [Google Scholar] [CrossRef]
- Fontaine, J.B.; Vanessa, C.W.; Enright, N.J.; Lade, J.C.; Miller, B.P. Fire behaviour in South-western Austrlian shrublands: Evuluating the influence of fuel age and fire weather. Int. J. Wildland Fire 2012, 21, 385–395. [Google Scholar] [CrossRef]
- Trollope, W.S.W.; Trollope, L.A.; Harnett, D.C. Fire behaviour a key factor in the ecology of African grassland and savannas. In Forest Fire Research and Wildland Fire Safety, Proceedings of the IV International Conference on Forest Fire Research, Wildland Fire Safety Summit, Coimbra, Portugal, 18–23 November 2002; Viega, D.X., Ed.; Millpress: Rotterdam, The Netherlands, 2002. [Google Scholar]
- Josens, G. Etude fonctionnelle de quelques animaux: Les termites. Bulletin de Liaison des Chercheurs de Lamto 1974, 5, 91–131. [Google Scholar]
- Josens, G. Recherches Sur la Structure et le Fonctionnement Des Nids Hypogés de Quatres Espèces de Macrotermitinae (Termitidae) Communes Dans Les Savannes de Lamto (Côte d’Ivoire); Mémoire de la Classe Des Sciences, Université Libre de Bruxelle: Bruxelles, Belgium, 1977; 122p. [Google Scholar]
- Wood, T.G.; Johnson, R.A. Abundance and vertical distribution in sail of Microtermes (Isoptera, Termitidae) in savanna woodland and agricultural ecosystems at Mokwa, Nigeria. Memorobiola Zoologica 1978, 29, 203–213. [Google Scholar]
- Koné, N.A.; Silué, S.K.; Niango, N.S.A.; Konaté, S.; Linsenmair, K.E. Variation saisonnière des biomasses de meules à champignon des termites Macrotermitinae dans différents types d’habitats d’une savane guinéenne (Lamto, Côte d’Ivoire). Revue Environnement et Biodiversité PASRES 2017, 2, 56–69. [Google Scholar]
- Frost, P.G.; Medina, E.; Menaut, J.C.; Solbrig, O.; Swift, M.; Walker, B.H. (Eds.) Response of Savannas to Stress and Disturbance; Biology International Special Issue 10; IUBS: Paris, France, 1986. [Google Scholar]
- Black, H.I.J.; Okwakol, M.J.N. Agricultural intensification, soil biodiversity and agroecosystem function in the tropics: The role of termites. Appl. Soil Ecol. 1997, 6, 37–53. [Google Scholar] [CrossRef]
- Davies, A.B.; Eggleton, P.; van Rensburg, B.J.; Parr, C.L. The pyrodiversity–biodiversity hypothesis: A test with savanna termite assemblages. J. Appl. Ecol. 2012, 49, 422–430. [Google Scholar] [CrossRef]




| Habitat Types | Total Species Richness | Sampled Coverage | Similarity between Transects | Simpson’s Index | Evenness’s Index | Abundance | Unique Species per Habitat |
|---|---|---|---|---|---|---|---|
| Gallery Forest | 37 | 85.33 | 0.43 | 0.48 | 0.78 | 209 | 10 |
| Forest Island | 34 | 95.8 | 0.43 | 0.46 | 0.84 | 187 | 6 |
| Woodland Savanna | 32 | 87.77 | 0.52 | 0.56 | 0.66 | 252 | 8 |
| Tree Savanna | 28 | 93.09 | 0.44 | 0.48 | 0.81 | 209 | 7 |
| Grassy Savanna | 17 | 94.5 | 0.69 | 0.51 | 0.89 | 141 | 3 |
| Habitat Types | GF | FI | WS | TS | GS |
|---|---|---|---|---|---|
| Gallery Forest | 0 | ||||
| Forest Island | 22/27/55% | 0 | |||
| Woodland Savanna | 20/29/59% | 23/20/50% | 0 | ||
| Tree Savanna | 17/31/65% | 15/32/68% | 21/19/46% | 0 | |
| Grassy Savanna | 10/34/77% | 9/33/79% | 11/27/71% | 14/17/54% | 0 |
| Habitats | |||||||
|---|---|---|---|---|---|---|---|
| Feeding Groups | GF | FI | WS | TS | GS | Total | p Values |
| Fungus growers | 113 (54.06) | 99 (52.94) | 157 (64.68) | 130 (62.2) | 61 (42.95) | 560 (56.05) | 0.001 |
| Wood feeders | 40 (19.13) | 43 (23) | 52 (19.33) | 41 (19.61) | 27 (19.01) | 203 (20.32) | 0.156 |
| Soil feeders | 56 (26.8) | 45 (24.06) | 42 (15.61) | 28 (13.39) | 24 (16.9) | 195 (19.51) | 0.014 |
| Grass feeders | 0 | 0 | 1 (0.37) | 10 (4.78) | 30 (21.12) | 41 (4.1) | 0.001 |
| p Values | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | - | - |
| Summary Statistic for CCA Ordination | Habitat-Specific Diversity and Abundance of Collected Termites in Dependence of Environmental Variables | Habitat-Specific Diversity of Collected Termite’s Feeding Groups in Dependence of Environmental Variables | Habitat-Specific Abundance of Collected Termite’s Feeding Groups in Dependence of Environmental Variables | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Axes | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 |
| Eigenvalues | 0.45 | 0.27 | 0.19 | 0.13 | 0.15 | 0.02 | 0 | - | 0.11 | 0.01 | 0 | - |
| Projected inertia (%) | 43.35 | 26.24 | 18.13 | 12.29 | 84.74 | 12.05 | 3.2 | - | 88.6 | 10.57 | 0.82 | - |
| Cumulative projected inertia (%) | 43.35 | 69.58 | 87.71 | 100 | 84.74 | 96.79 | 100 | - | 88.61 | 99.18 | 100 | - |
| Total unconstrained inertia | 0.10 | 0.18 | 0.13 | |||||||||
| Inertia (%) | 100 | 100 | 100 | |||||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koné, N.A.; Silué, K.S.; Konaté, S.; Linsenmair, K.E. Determinants of Termite Assemblages’ Characteristics within Natural Habitats of a Sudano-Guinean Savanna (Comoe National Park, Côte d’Ivoire). Insects 2018, 9, 189. https://doi.org/10.3390/insects9040189
Koné NA, Silué KS, Konaté S, Linsenmair KE. Determinants of Termite Assemblages’ Characteristics within Natural Habitats of a Sudano-Guinean Savanna (Comoe National Park, Côte d’Ivoire). Insects. 2018; 9(4):189. https://doi.org/10.3390/insects9040189
Chicago/Turabian StyleKoné, N’golo Abdoulaye, Kolotchèlèma Simon Silué, Souleymane Konaté, and Karl Eduard Linsenmair. 2018. "Determinants of Termite Assemblages’ Characteristics within Natural Habitats of a Sudano-Guinean Savanna (Comoe National Park, Côte d’Ivoire)" Insects 9, no. 4: 189. https://doi.org/10.3390/insects9040189