Geographic and Ecological Dimensions of Host Plant-Associated Genetic Differentiation and Speciation in the Rhagoletis cingulata (Diptera: Tephritidae) Sibling Species Group

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
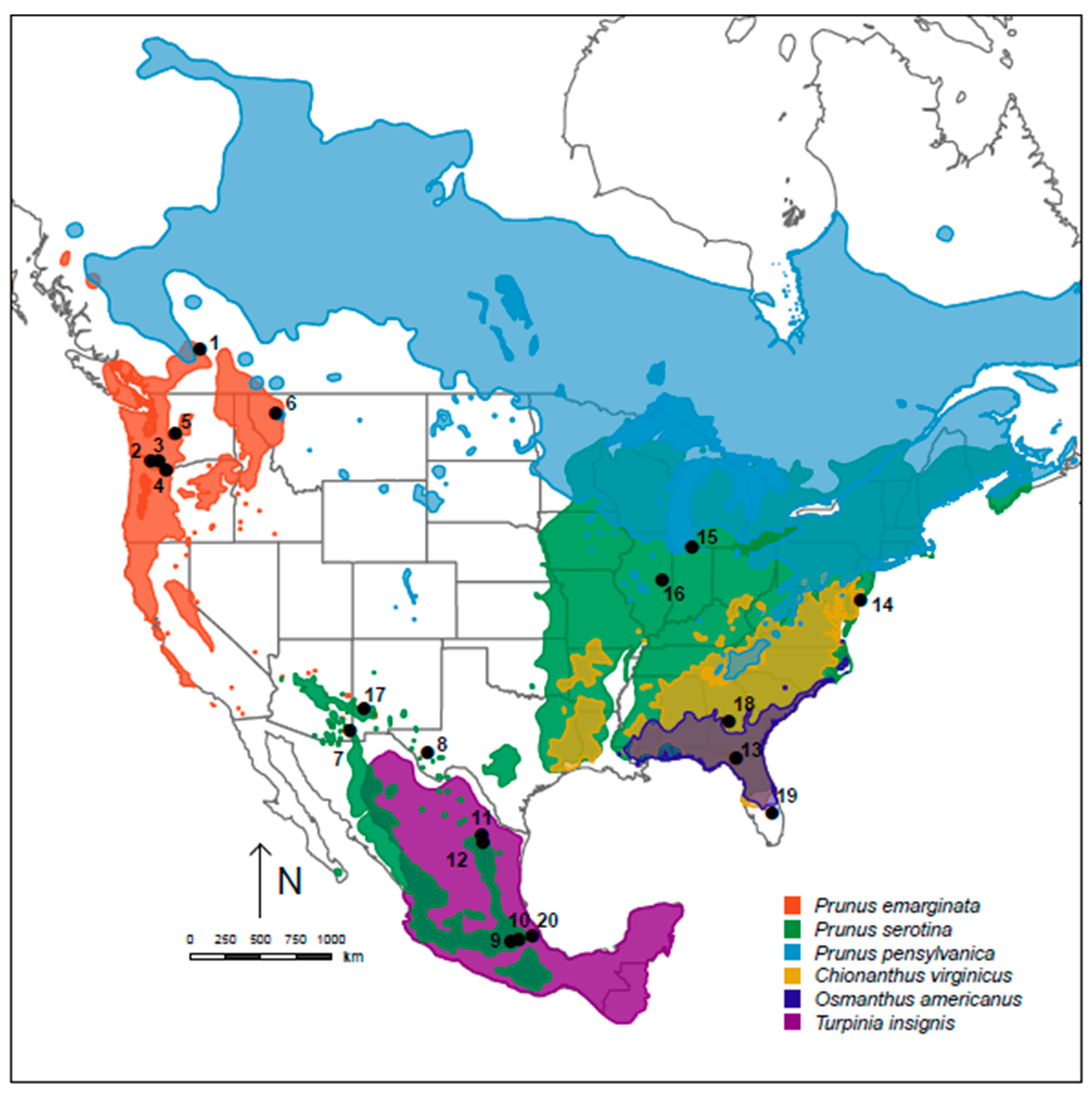
2.1. Study Sites and Collections
2.2. Microsatellite Genotyping
2.3. Genetic Analysis of Microsatellites
2.4. Mitochondrial DNA
2.5. Crosses Testing for Postzygotic RI
3. Results
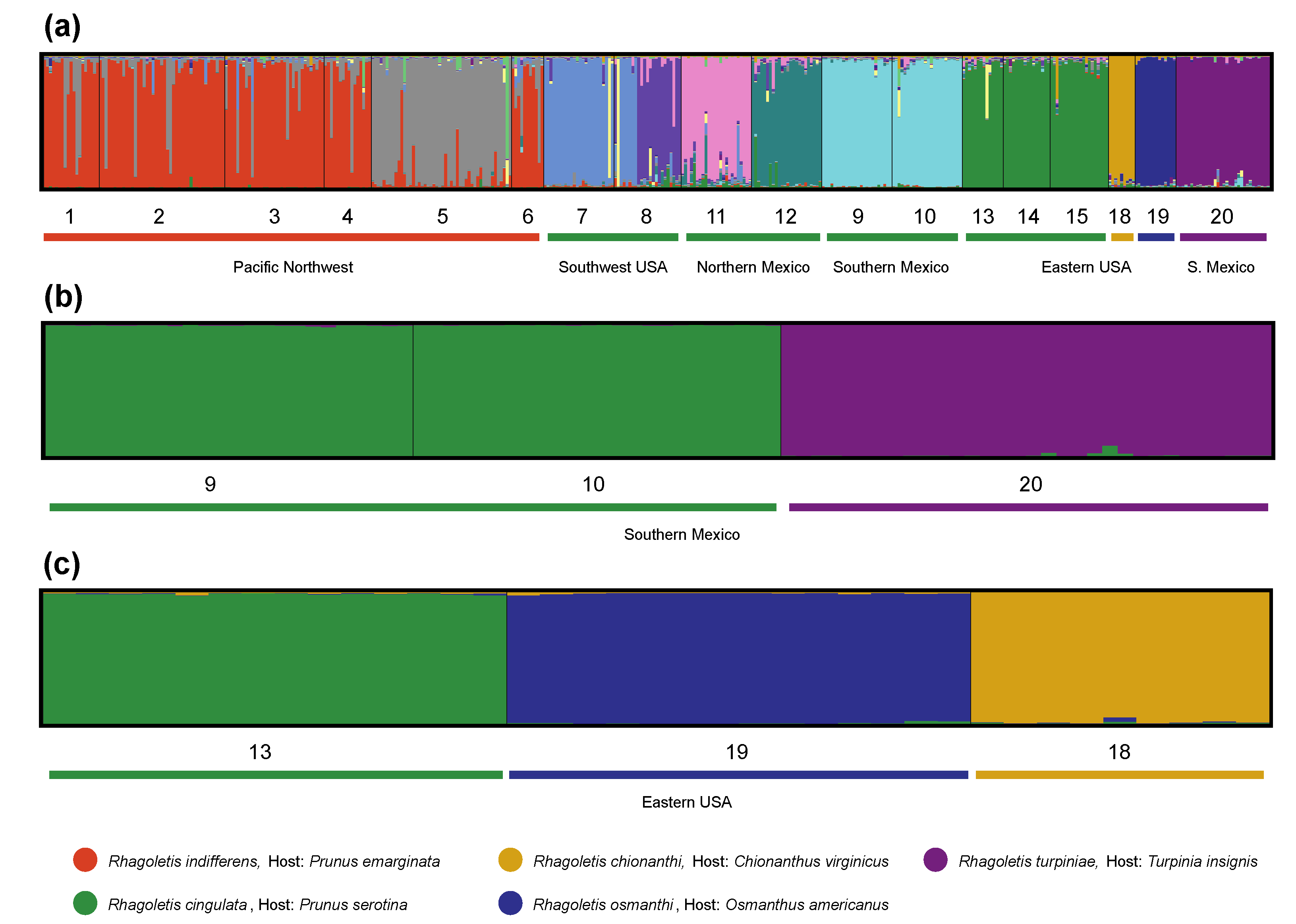
3.1. Nuclear Encoded Microsatellites
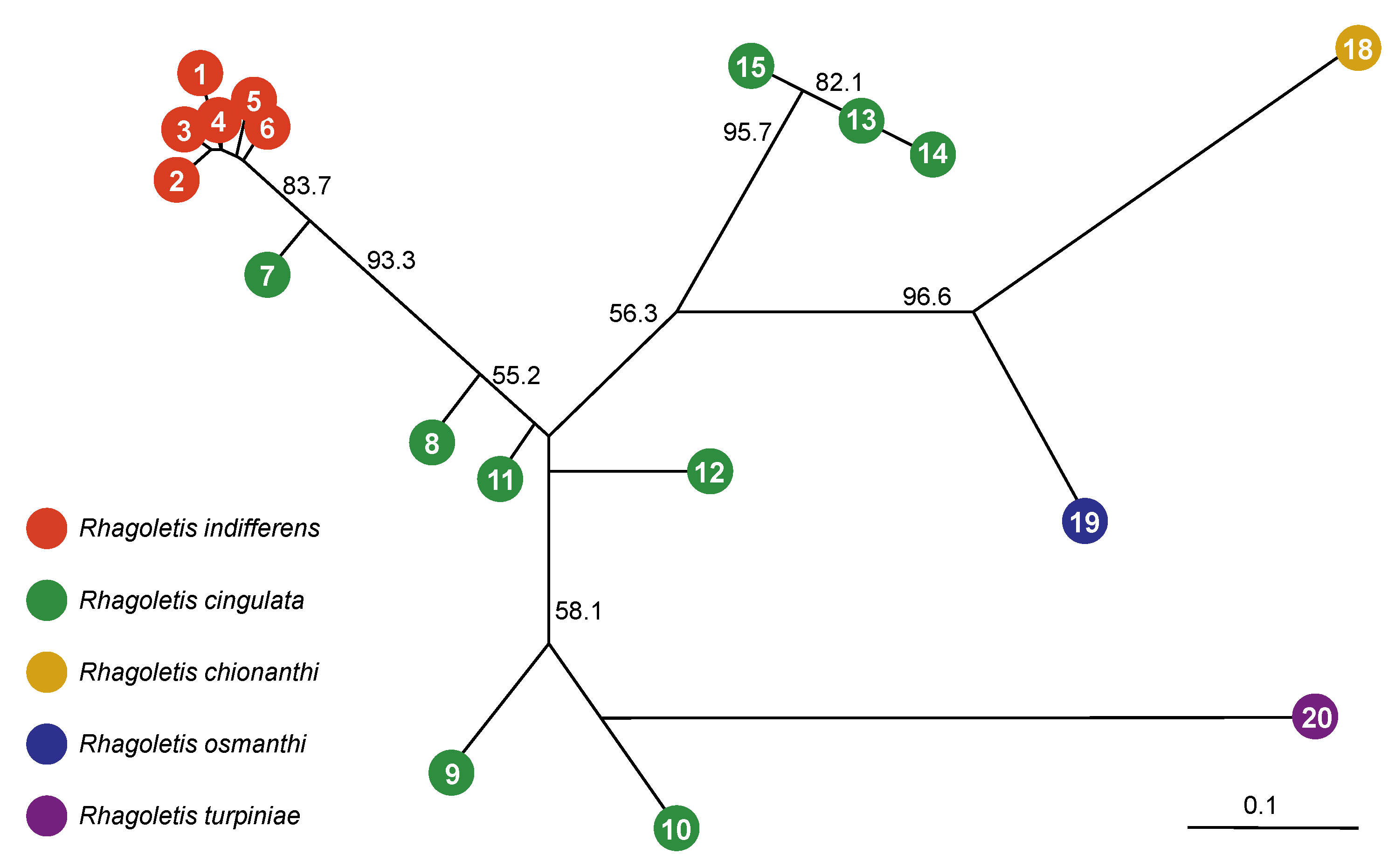
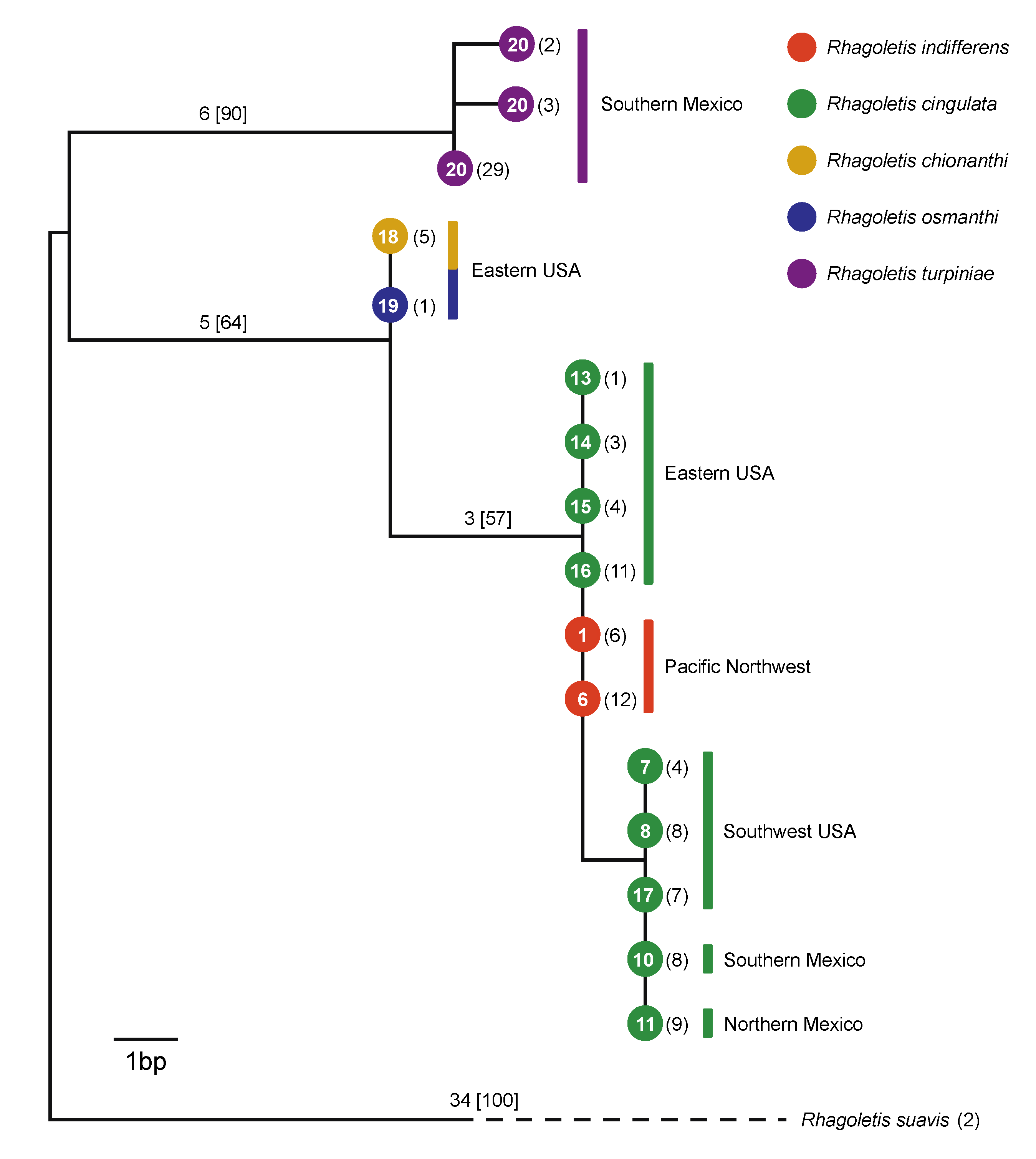
3.2. Mitochondrial DNA
3.3. Postzygotic RI
4. Discussion
4.1. Divergence Timing and Ecological Versus Geographic Isolation
4.2. Pre- and Postzygotic RI and Geographic Isolation
4.3. Cytoplasmic Incompatibility and Patterns of mtDNA Variation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Givnish, T.J.; Sytsma, K.J. Molecular Evolution and Adaptive Radiation; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Simpson, G.G. The Major Features of Evolution; Columbia University Press: New York, NY, USA, 1953. [Google Scholar]
- Stroud, J.T.; Losos, J.B. Ecological opportunity and adaptive radiation. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 507–532. [Google Scholar] [CrossRef]
- Hood, G.R.; Forbes, A.A.; Powell, T.H.Q.; Egan, S.P.; Hamerlinck, G.; Smith, J.J.; Feder, J.L. Sequential divergence and the multiplicative origin of community diversity. Proc. Natl. Acad. Sci. USA 2015, 112, E5980–E5989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rundell, R.J.; Price, T.D. Adaptive radiation, nonadaptive radiation, ecological speciation and nonecological speciation. Trends Ecol. Evol. 2009, 24, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Coyne, J.A.; Orr, H.A. Speciation; Sinauer Associates Inc.: Sunderland, MA, USA, 2004. [Google Scholar]
- Nosil, P.; Flaxman, S.M. Conditions for mutation-order speciation. Proc. R. Soc. B Biol. Sci. 2011, 278, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Pfennig, D.W.; Pfennig, K.S. Evolution’s Wedge: Competition and the Origins of Diversity; University of California Press: Oakland, CA, USA, 2012. [Google Scholar]
- Dufour, C.M.S.; Herrel, A.; Losos, J.B. Ecological character displacement between a native and an introduced species: The invasion of Anolis cristatellus in Dominica. Biol. J. Linn. Soc. 2018, 123, 43–54. [Google Scholar] [CrossRef]
- Driscoe, A.; Nice, C.; Busbee, R.; Hood, G.; Egan, S.P.; Ott, J. Host plant associations and geography interact to shape diversification in a specialist insect herbivore. Mol. Ecol. 2019, in press. [Google Scholar]
- Doellman, M.M.; Feder, J.L. Genomic transitions during host race and species formation. Curr. Opin. Insect Sci. 2019, 31, 84–92. [Google Scholar] [CrossRef]
- Bush, G.L. Sympatric host race formation and speciation in frugivorous flies of the genus Rhagoletis (Diptera, Tephritidae). Evolution 1969, 23, 237–251. [Google Scholar] [CrossRef]
- Feder, J.L.; Chilcote, C.A.; Bush, G.L. Genetic differentiation between sympatric host races of the apple maggot fly Rhagoletis pomonella. Nature 1988, 336, 61–64. [Google Scholar] [CrossRef]
- Powell, T.H.Q.; Forbes, A.A.; Hood, G.R.; Feder, J.L. Ecological adaptation and reproductive isolation in sympatry: Genetic and phenotypic evidence for native host races of Rhagoletis pomonella. Mol. Ecol. 2014, 23, 688–704. [Google Scholar] [CrossRef]
- Doellman, M.M.; Ragland, G.J.; Hood, G.R.; Meyers, P.J.; Egan, S.P.; Powell, T.H.Q.; Lazorchak, P.; Glover, M.M.; Tait, C.; Schuler, H.; et al. Genomic differentiation during speciation-with-gene-flow: Comparing geographic and host-related variation in divergent life history adaptation in Rhagoletis pomonella. Genes 2018, 9, 262. [Google Scholar] [CrossRef]
- Doellman, M.M.; Egan, S.P.; Ragland, G.J.; Meyers, P.J.; Hood, G.R.; Powell, T.H.Q.; Lazorchak, P.; Hahn, D.A.; Berlocher, S.H.; Nosil, P.; et al. Standing geographic variation in eclosion time and the genomics of host race formation in Rhagoletis pomonella fruit flies. Ecol. Evol. 2019, 9, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Bush, G.L. The taxonomy, cytology, and evolution of the genus Rhagoletis in North America (Diptera, Tephritidae). Bull. Museum Comp. Zool. Harvard Univ. 1966, 134, 431–562. [Google Scholar]
- Berlocher, S.H.; McPheron, B.A.; Feder, J.L.; Bush, G.L. Genetic differentiation at allozyme loci in the Rhagoletis pomonella (Diptera: Tephritidae) species complex. Ann. Entomol. Soc. Am. 1993, 86, 716–727. [Google Scholar] [CrossRef]
- Berlocher, S.H.; Feder, J.L. Sympatric speciation in phytophagous insects: Moving beyond controversy? Annu. Rev. Entomol. 2002, 47, 773–815. [Google Scholar] [CrossRef] [PubMed]
- McPheron, B.A.; Smith, D.C.; Berlocher, S.H. Genetic differences between host races of Rhagoletis pomonella. Nature 1988, 336, 64–66. [Google Scholar] [CrossRef]
- Glover, M.M.; Egan, S.P.; Hood, G.R.; Rull, J.; Aluja, M.; Feder, J.L. Phylogeography of walnut-infesting Rhagoletis suavis (Diptera: Tephritidae) flies. Insect Syst. Divers. 2018, 2, 1–9. [Google Scholar] [CrossRef]
- Glover, M.M. The Ecological and Evolutionary Drivers of Speciation in Walnut-Infesting Flies; University of Notre Dame: Notre Dame, IN, USA, 2018. [Google Scholar]
- Smith, J.J.; Powell, T.H.Q.; Teixeira, L.; Armstrong, W.O.; McClowry, R.J.; Isaacs, R.; Hood, G.R.; Feder, J.L.; Gut, L. Genetic structure of cherry fruit fly (Rhagoletis cingulata) populations across managed, unmanaged, and natural habitats. Entomol. Exp. Appl. 2014, 150, 157–165. [Google Scholar] [CrossRef]
- Maxwell, S.A.; Thistlewood, H.M.A.; Keyghobadi, N. Population genetic structure of the western cherry fruit fly Rhagoletis indifferens (Diptera: Tephritidae) in British Columbia, Canada. Agric. For. Entomol. 2014, 16, 33–44. [Google Scholar] [CrossRef]
- Saint Jean, G.; Hood, G.R.; Egan, S.P.; Powell, T.H.Q.; Schuler, H.; Doellman, M.M.; Glover, M.M.; Smith, J.J.; Yee, W.L.; Goughnour, R.B.; et al. Limited genetic evidence for host plant-related differentiation in the western cherry fruit fly, Rhagoletis indifferens. Entomol. Exp. Appl. 2018. [Google Scholar] [CrossRef]
- Hood, G.R.; Egan, S.P.; Feder, J.L. Evidence for sexual isolation as a prezygotic barrier to gene flow between morphologically divergent species of Rhagoletis fruit flies. Ecol. Entomol. 2012, 37, 521–528. [Google Scholar] [CrossRef]
- Little, E., Jr. Atlas of United States Trees. Volume 1. Conifers and Important Hardwoods; US Dept. of Agriculture, Forest Service: Washington, DC, USA, 1971.
- Little, E., Jr. Atlas of United States Trees. Volume 3. Minor Western Hardwoods; US Dept. of Agriculture, Forest Service: Washington, DC, USA, 1976.
- Clone of Digital Representations of Tree Species Range Maps from “Atlas of United States Trees” by Elbert, L.; Little, Jr. (and other publications). Available online: https://github.com/wpetry/USTreeAtlas (accessed on 12 July 2019).
- Kew Science, Plants of the World Online. Available online: http://www.plantsoftheworldonline.org/taxon/urn:lsid:ipni.org:names:77183581-1#source-KBD (accessed on 20 July 2019).
- Sosa, V. Flora de Verazruz, Staphyleaceae; Xalapa: Veracruz, Mexico, 1988. [Google Scholar]
- Hernandez-Ortiz, V. Description of a New Rhagoletis Species From Tropical Mexico (Diptera: Tephritidae) View Project; Entomological Society of Washington: Washington, DC, USA, 1993. [Google Scholar]
- Aluja, M.; Lozada, N.; Piñero, J.; Birke, A.; Hernández-Ortiz, V.; Díaz-Fleischer, F. Basic behavior of Rhagoletis turpiniae (Diptera: Tephritidae) with comparative notes on the sexual behavior of Rhagoletis pomonella and Rhagoletis zoqui. Ann. Entomol. Soc. Am. 2001, 94, 268–274. [Google Scholar] [CrossRef]
- SEINet, Turpinia insignis. Available online: http://swbiodiversity.org/seinet/collections/map/googlemap.php?usethes=1&taxa=134230 (accessed on 20 July 2019).
- Doellman, M.M.; Saint Jean, G.; Egan, S.P.; Powell, T.H.Q.; Hood, G.R.; Schuler, H.; Bruzzese, D.J.; Glover, M.M.; Smith, J.J.; Yee, W.L. Contrasting patterns of genetic differentiation reveal complex geographic history of divergence for North American cherry-infesting Rhagoletis flies. J. Biogeogr. 2019. in review. [Google Scholar]
- Allen, C.D.; Betancourt, J.L.; Swetnam, T.W. Landscape changes in the southwestern United States: Techniques, long-term data sets, and trends. USGS Land Use Hist. N. Am. 1998, 71–84. [Google Scholar]
- Aguirre-Planter, E.; Furnier, G.R.; Eguiarte, L.E. Low levels of genetic variation within and high levels of genetic differentiation among populations of species of Abies from southern Mexico and Guatemala. Am. J. Bot. 2000, 87, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Selem, L.; Heine, K. Late Quaternary glaciations of México. In Quaternary Glaciations—Extent and Chronology, Part III. South America, Asia, Africa, Australia, Antarctica; Ehlers, J., Gibbard, P.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 223–242. [Google Scholar]
- Jaramillo-Correa, J.P.; Aguirre-Planter, E.; Khasa, D.P.; Eguiarte, L.E.; Piñero, D.; Furnier, G.R.; Bousquet, J. Ancestry and divergence of subtropical montane forest isolates: Molecular biogeography of the genus Abies (Pinaceae) in southern México and Guatemala. Mol. Ecol. 2008, 17, 2476–2490. [Google Scholar] [CrossRef] [PubMed]
- Neilson, W.T.A.; McAllan, J.W. Effects of mating on fecundity of the apple maggot, Rhagoletis pomonella (Walsh). Can. Entomol. 1965, 97, 276–279. [Google Scholar] [CrossRef]
- Feder, J.L.; Chilcote, C.A.; Bush, G.L. The geographic pattern of genetic differentiation between host associated populations of Rhagoletis pomonella (Diptera: Tephritidae) in the Eastern United States and Canada. Evolution 1990, 44, 570–594. [Google Scholar] [CrossRef] [PubMed]
- Tadeo, E.; Feder, J.L.; Egan, S.P.; Schuler, H.; Aluja, M.; Rull, J. Divergence and evolution of reproductive barriers among three allopatric populations of Rhagoletis cingulata across eastern North America and Mexico. Entomol. Exp. Appl. 2015, 156, 301–311. [Google Scholar] [CrossRef]
- Maxwell, S.A.; Rasic, G.; Keyghobadi, N. Characterization of microsatellite loci for the western cherry fruit fly, Rhagoletis indifferens (Diptera: Tephritidae). Mol. Ecol. Resour. 2009, 9, 1025–1028. [Google Scholar] [CrossRef]
- Velez, S.; Taylor, M.S.; Noor, M.A.F.; Lobo, N.F.; Feder, J.L. Isolation and characterization of microsatellite loci from the apple maggot fly Rhagoletis pomonella (Diptera: Tephritidae). Mol. Ecol. Notes 2006, 6, 90–92. [Google Scholar] [CrossRef]
- Michel, A.P.; Sim, S.; Powell, T.H.Q.; Taylor, M.S.; Nosil, P.; Feder, J.L. Widespread genomic divergence during sympatric speciation. Proc. Natl. Acad. Sci. USA 2010, 107, 9724–9729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Hey, J.; Nielsen, R. Multilocus methods for estimating population sizes, migration rates and divergence time, with applications to the divergence of Drosophila pseudoobscura and D. persimilis. Genetics 2004, 167, 747–760. [Google Scholar] [CrossRef]
- Hey, J. Isolation with migration models for more than two populations. Mol. Biol. Evol. 2010, 27, 905–920. [Google Scholar] [CrossRef]
- Sethuraman, A.; Hey, J. IMa2p-parallel MCMC and inference of ancient demography under the Isolation with migration (IM) model. Mol. Ecol. Resour. 2016, 16, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Estoup, A.; Angers, B. Microsatellites and minisatellites for molecular ecology: Theoretical and empirical considerations. In Advances in Molecular Ecology; Carvalho, G., Ed.; NATO press: Amsterdam, The Netherlands, 1998; pp. 55–86. [Google Scholar]
- Schug, M.D.; Mackay, T.F.C.; Aquadro, C.F. Low mutation rates of microsatellite loci in Drosophila melanogaster. Nat. Genet. 1997, 15, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.F.; Pérez, T.; Albornoz, J.; Domínguez, A. Estimation of microsatellite mutation rates in Drosophila melanogaster. Genet. Res. 2000, 76, 323–326. [Google Scholar] [CrossRef]
- Peccoud, J.; Ollivier, A.; Plantegenest, M.; Simon, J.-C. A continuum of genetic divergence from sympatric host races to species in the pea aphid complex. Proc. Natl. Acad. Sci. USA 2009, 106, 7495–7500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, T.H.Q.; Hood, G.R.; Murphy, M.O.; Heilveil, J.S.; Berlocher, S.H.; Nosil, P.; Feder, J.L. Genetic divergence along the speciation continuum: The transition from host race to species in Rhagoletis (Diptera: Tephritidae). Evolution 2013, 67, 2561–2576. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Frick, K.E.; Simkover, H.G.; Telford, H.S. Bionomics of the cherry fruit flies in eastern Washington. Tech. Bull. Agric. Exp. Station. Washingt. State Inst. Agric. Sci. 1954, 13, 1–66. [Google Scholar]
- Berlocher, S.H. Radiation and divergence in the Rhagoletis pomonella species group: Inferences from allozymes. Evolution 2000, 54, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Michel, A.P.; Rull, J.; Aluja, M.; Feder, J.L. The genetic structure of hawthorn-infesting Rhagoletis pomonella populations in Mexico: Implications for sympatric host race formation. Mol. Ecol. 2007, 16, 2867–2878. [Google Scholar] [CrossRef]
- Xie, X.; Rull, J.; Michel, A.P.; Velez, S.; Forbes, A.A.; Lobo, N.F.; Aluja, M.; Feder, J.L. Hawthorn-infesting populations of Rhagoletis pomonella in Mexico and speciation mode plurality. Evolution 2007, 61, 1091–1105. [Google Scholar] [CrossRef]
- Xie, X.; Michel, A.P.; Schwarz, D.; Rull, J.; Velez, S.; Forbes, A.A.; Aluja, M.; Feder, J.L. Radiation and divergence in the Rhagoletis pomonella species complex: Inferences from DNA sequence data. J. Evol. Biol. 2008, 21, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Blanc, F.L.; Keifer, H.H. The cherry fruit fly in North America. Morphological differences between the eastern and western subspecies of the cherry fruit fly, Rhagoletis cingulata (Loew). Calif. Dep. Agric. Bull. 1955, 44, 77–88. [Google Scholar]
- Beckstead, W.A.; Ebbert, M.T.W.; Rowe, M.J.; McClellan, D.A. Evolutionary pressure on mitochondrial cytochrome b is consistent with a role of cytbI7T affecting longevity during caloric restriction. PLoS ONE 2009, 4, e5836. [Google Scholar] [CrossRef] [PubMed]
- Galtier, N.; Nabholz, B.; Glémin, S.; Hurst, G.D.D. Mitochondrial DNA as a marker of molecular diversity: A reappraisal. Mol. Ecol. 2009, 18, 4541–4550. [Google Scholar] [CrossRef]
- Silva, G.; Lima, F.P.; Martel, P.; Castilho, R. Thermal adaptation and clinal mitochondrial DNA variation of European anchovy. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuler, H.; Bertheau, C.; Egan, S.P.; Feder, J.L.; Riegler, M.; Schlick-Steiner, B.C.; Steiner, F.M.; Johannesen, J.; Kern, P.; Tuba, K.; et al. Evidence for a recent horizontal transmission and spatial spread of Wolbachia from endemic Rhagoletis cerasi (Diptera: Tephritidae) to invasive Rhagoletis cingulata in Europe. Mol. Ecol. 2013, 22, 4101–4111. [Google Scholar] [CrossRef] [PubMed]
- Bordenstein, S.R. Symbiosis and the origin of species. In Insect Symbiosis; Bourtzis, K., Miller, T., Eds.; CRC Press: Boca Raton, FL, USA, 2003; pp. 283–304. ISBN 9780203009918. [Google Scholar]
- Gotoh, T.; Noda, H.; Fujita, T.; Iwadate, K.; Higo, Y.; Saito, S.; Ohtsuka, S. Wolbachia and nuclear–nuclear interactions contribute to reproductive incompatibility in the spider mite Panonychus mori (Acari: Tetranychidae). Heredity 2005, 94, 237–246. [Google Scholar] [CrossRef]
- Werren, J.H. Wolbachia and speciation. In Endless Forms: Species and Speciation; Howard, D.J., Berlocher, S.H., Eds.; Oxford University Press: New York, NY, USA, 1998; pp. 245–260. [Google Scholar]
- Bordenstein, S.R.; Werren, J.H. Bidirectional incompatibility among divergent Wolbachia and incompatibility level differences among closely related Wolbachia in Nasonia. Heredity 2007, 99, 278–287. [Google Scholar] [CrossRef]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Engelstädter, J.; Hurst, G.D.D. The ecology and evolution of microbes that manipulate host reproduction. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 127–149. [Google Scholar] [CrossRef]
- Brucker, R.M.; Bordenstein, S.R. Speciation by symbiosis. Trends Ecol. Evol. 2012, 27, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Arthofer, W.; Riegler, M.; Schneider, D.; Krammer, M.; Miller, W.J.; Stauffer, C. Hidden Wolbachia diversity in field populations of the European cherry fruit fly, Rhagoletis cerasi (Diptera, Tephritidae). Mol. Ecol. 2009, 18, 3816–3830. [Google Scholar] [CrossRef] [PubMed]
- Riegler, M.; Stauffer, C. Wolbachia infections and superinfections in cytoplasmically incompatible populations of the European cherry fruit fly Rhagoletis cerasi (Diptera, Tephritidae). Mol. Ecol. 2002, 11, 2425–2434. [Google Scholar] [CrossRef] [PubMed]
- Schuler, H.; Köppler, K.; Daxböck-Horvath, S.; Rasool, B.; Krumböck, S.; Schwarz, D.; Hoffmeister, T.S.; Schlick-Steiner, B.C.; Steiner, F.M.; Telschow, A.; et al. The hitchhiker’s guide to Europe: The infection dynamics of an ongoing Wolbachia invasion and mitochondrial selective sweep in Rhagoletis cerasi. Mol. Ecol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Price, P.W. Species interactions and the evolution of biodiversity. In Plant-Animal Interactions; Pellmyr, O., Herrera, C., Eds.; Blackwell Science Ltd: Oxford, UK, 2002; pp. 3–26. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Site Location | Lat. N, Long. W. | n |
|---|---|---|---|
| 1 | Salmon Arm, British Columbia, Canada | 50°54′54″, 119°21′26″ | 19 |
| 2 | L. Kalama Road, Woodland, Washington | 45°56′24″, 122°40′41″ | 43 |
| 3 | Lewis River, Woodland, Washington | 45°56′20″, 122°38′5″ | 34 |
| 4 | Trout Creek Rd, Hood River, Oregon | 45°32′10″, 121°37′14″ | 16 |
| 5 | Ronald, Kittitas County, Washington | 47°23′50″, 121°02′67″ | 48 |
| 6 | Somers, Montana | 48°05′24″, 114°13′48″ | 11 |
| 7 | Chiricahua Mountains, Arizona | 31°55′44″, 109°5′15″ | 24 |
| 8 | Davis Mountains, Texas | 30°38′47″, 104°01′08″ | 23 |
| 9 | San Martin, Texmelucan, Mexico | 19°16′ 20″, 98°25′03″ | 24 |
| 10 | Huamantla, Tlaxcala, Mexico | 19°18′54″, 97°52′28″ | 24 |
| 11 | Los Lirios, Coahuila, Mexico | 25°20′58″, 100°17′46″ | 24 |
| 12 | Nueva Leon, Mexico | 25°48′48″, 100°22′02″ | 24 |
| 13 | Live Oak, Suwannee County, Florida | 30°22′06″, 83°14′38″ | 14 |
| 14 | Green Creek, Cape May, New Jersey | 39°02′11″, 74°54′05″ | 16 |
| 15 | South Bend, Indiana | 41°45′28″, 86°12′4″ | 20 |
| 16 | Urbana, Illinois | 45°38′25″, 122°35′33″ | mtDNA |
| 17 | Gila Cliff Dwelling Mon., New Mexico | 33°10′44″, 108°12′14″ | mtDNA |
| 18 | Perry, Georgia | 32°27′19″, 83°43′41″ | 9 |
| 19 | Lake Lizzie, Florida | 27°11′4″, 80°50′26″ | 15 |
| 20 | Xalapa, Veracruz, Mexico | 19°32′39″, 96°57′04″ | 32 |
| Level of Comparison | Region | n | Distance (s.e.) |
|---|---|---|---|
| Between species that co-occur in region | |||
| R. chionanthi versus R. cingulata | Southeastern USA | 1 | 0.608 |
| R. osmanthi versus R. cingulata | Southeastern USA | 1 | 0.555 |
| R. osmanthi versus R. chionanthi | Southeastern USA | 1 | 0.395 |
| R. turpiniae versus R. cingulata | Southern Mexico | 2 | 0.574 (0.086) |
| Between species in different geographic regions | |||
| R. indifferens versus R. cingulata | Pacific Northwest (PNW)/Eastern USA | 18 | 0.613 (0.014) |
| R. indifferens versus R. cingulata | PNW/Southern Mexico | 12 | 0.463 (0.007) |
| Within species between different geographic regions | |||
| R. cingulata | Eastern USA/Southern Mexico | 6 | 0.460 (0.033) |
| Within species in same geographic region | |||
| R. indifferens | PNW | 10 | 0.046 (0.004) |
| R. cingulata | Eastern USA | 3 | 0.060 (0.012) |
| R. cingulata | Southern Mexico | 1 | 0.103 |
| Cross (m x f) | n Cages | Mean Eggs Laid ± s.e. | Percent Egg Hatch ± s.e. | Mean n Eggs Hatched ± s.e. |
|---|---|---|---|---|
| R. indifferens x R. indifferens | 21 | 29.2 ± 5.5 | 83.0 ± 4.9a | 25.0 ± 5.0 |
| R. cingulata x R. indifferens | 10 | 41.5 ± 6.8 | 52.0 ± 11.5ab | 20.8 ± 7.1 |
| R. indifferens x R. cingulata | 14 | 35.7 ± 12.3 | 64.0 ± 6.5b | 22.8 ± 7.6 |
| R. cingulata x R. cingulata | 6 | 34.7 ± 9.7 | 46.0 ± 14.5b | 11.2 ± 4.5 |
| F3,47 | 0.36 | 4.48 | 0.541 | |
| p | 0.782 | 0.0076 | 0.656 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doellman, M.M.; Schuler, H.; Jean Saint, G.; Hood, G.R.; Egan, S.P.; Powell, T.H.Q.; Glover, M.M.; Bruzzese, D.J.; Smith, J.J.; Yee, W.L.; et al. Geographic and Ecological Dimensions of Host Plant-Associated Genetic Differentiation and Speciation in the Rhagoletis cingulata (Diptera: Tephritidae) Sibling Species Group. Insects 2019, 10, 275. https://doi.org/10.3390/insects10090275
Doellman MM, Schuler H, Jean Saint G, Hood GR, Egan SP, Powell THQ, Glover MM, Bruzzese DJ, Smith JJ, Yee WL, et al. Geographic and Ecological Dimensions of Host Plant-Associated Genetic Differentiation and Speciation in the Rhagoletis cingulata (Diptera: Tephritidae) Sibling Species Group. Insects. 2019; 10(9):275. https://doi.org/10.3390/insects10090275
Chicago/Turabian StyleDoellman, Meredith M., Hannes Schuler, Gilbert Jean Saint, Glen R. Hood, Scott P. Egan, Thomas H.Q. Powell, Mary M. Glover, Daniel J. Bruzzese, James J. Smith, Wee L. Yee, and et al. 2019. "Geographic and Ecological Dimensions of Host Plant-Associated Genetic Differentiation and Speciation in the Rhagoletis cingulata (Diptera: Tephritidae) Sibling Species Group" Insects 10, no. 9: 275. https://doi.org/10.3390/insects10090275