Chemical Modification of Aptamers for Increased Binding Affinity in Diagnostic Applications: Current Status and Future Prospects


Abstract
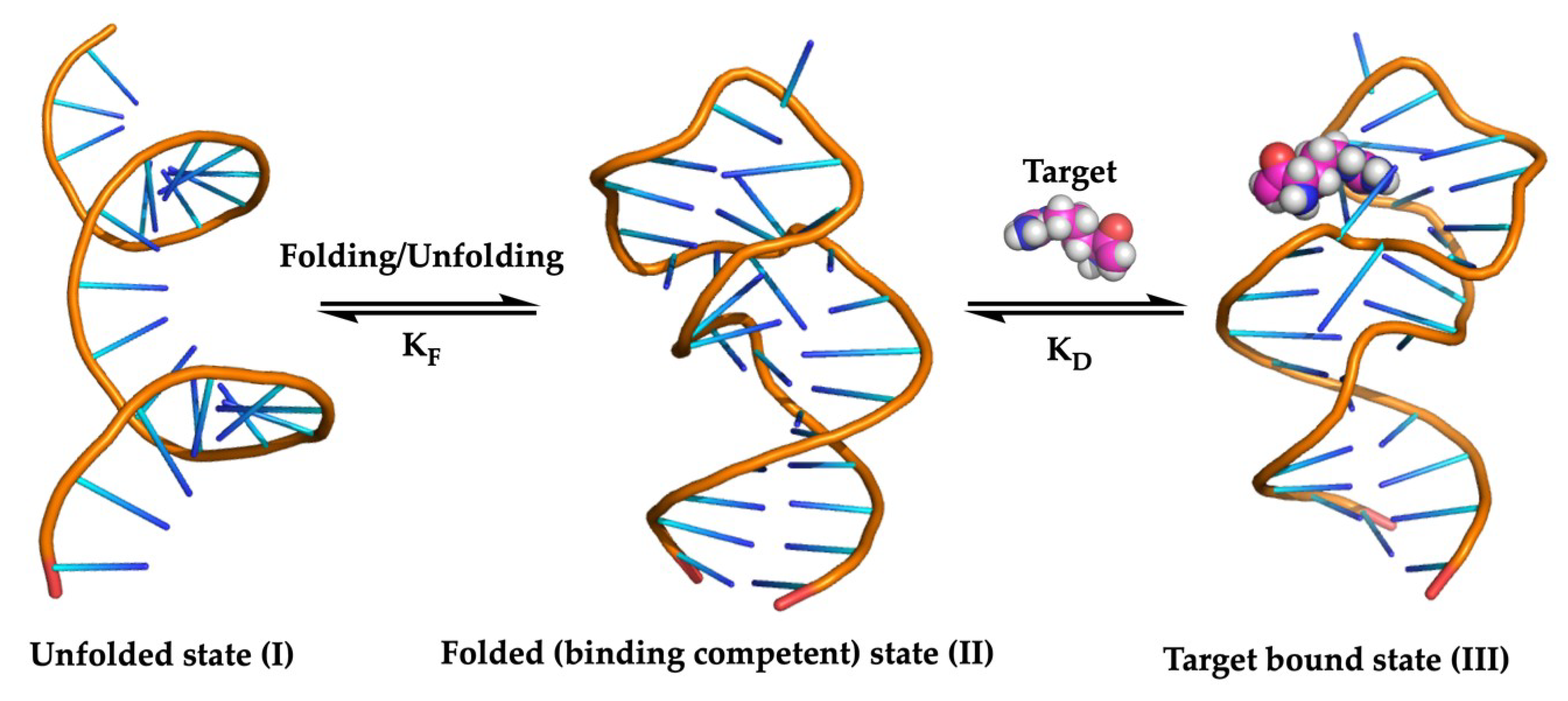
:1. Introduction
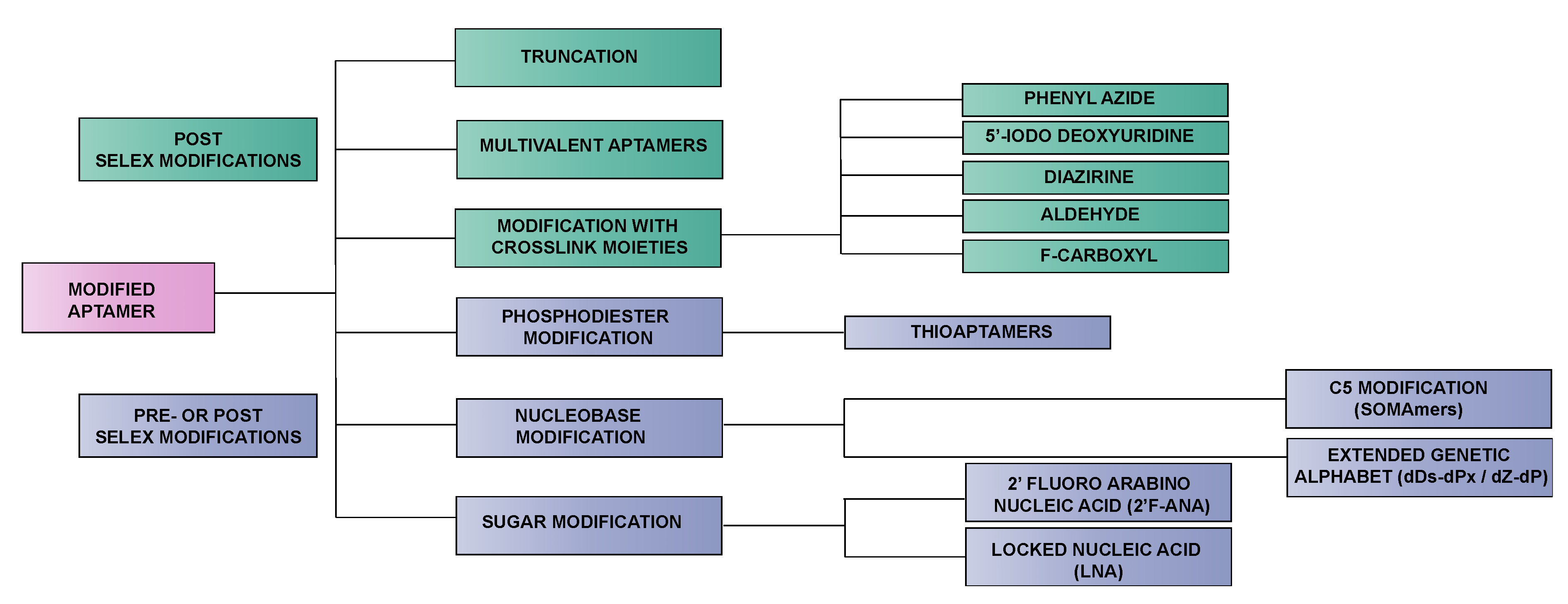
2. Methods for Introducing Chemical Modifications into Aptamers
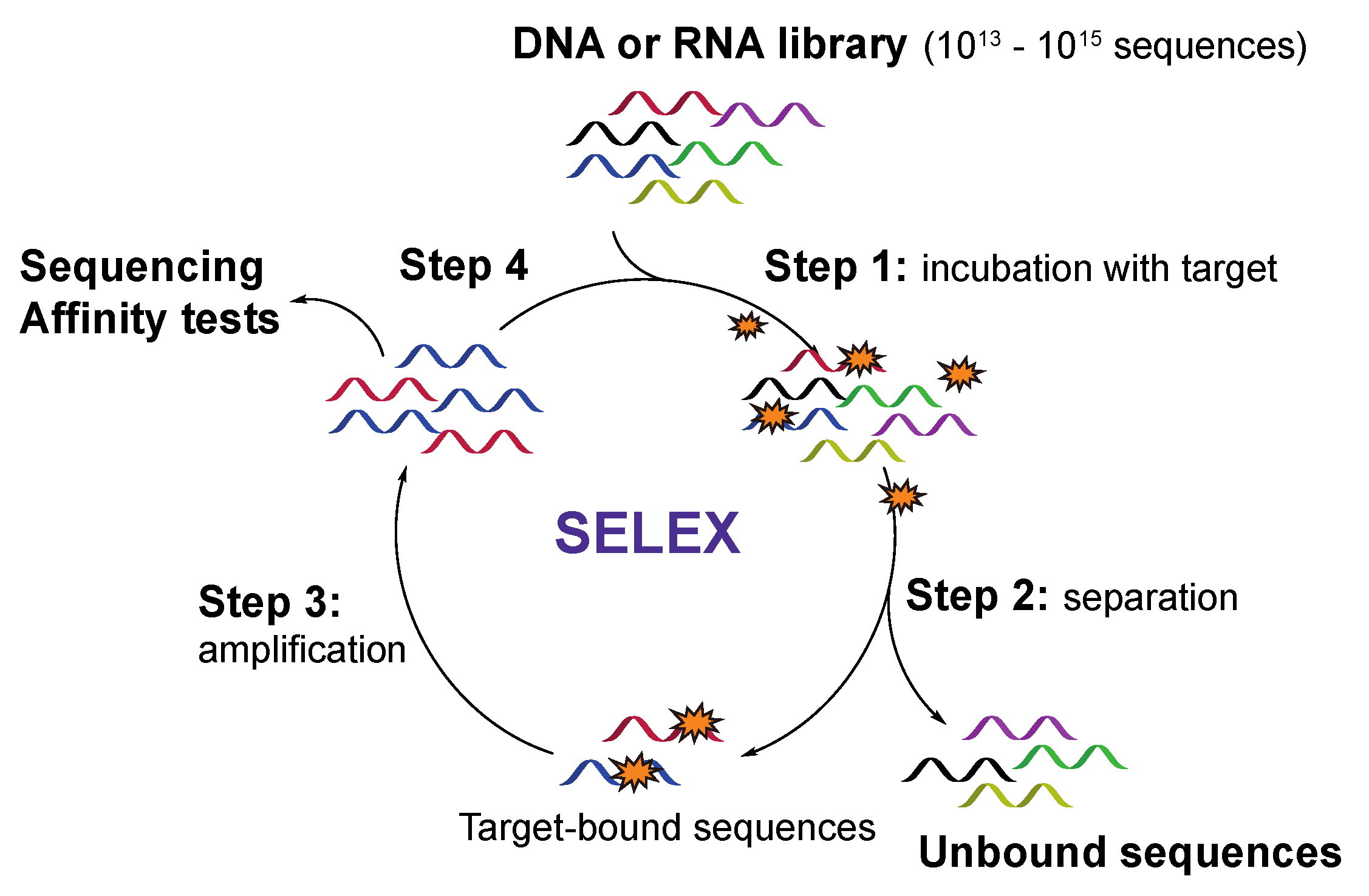
2.1. The SELEX Strategy for Aptamer Discovery
2.2. Pre-SELEX vs. Post-SELEX Modifications
3. Strategies for Improving Binding Affinity Through Chemical Aptamer Modification: Non-Covalent Target Binding
3.1. Modified Aptamers for Enhanced Non-Covalent Target Interaction
3.1.1. Truncated Aptamers
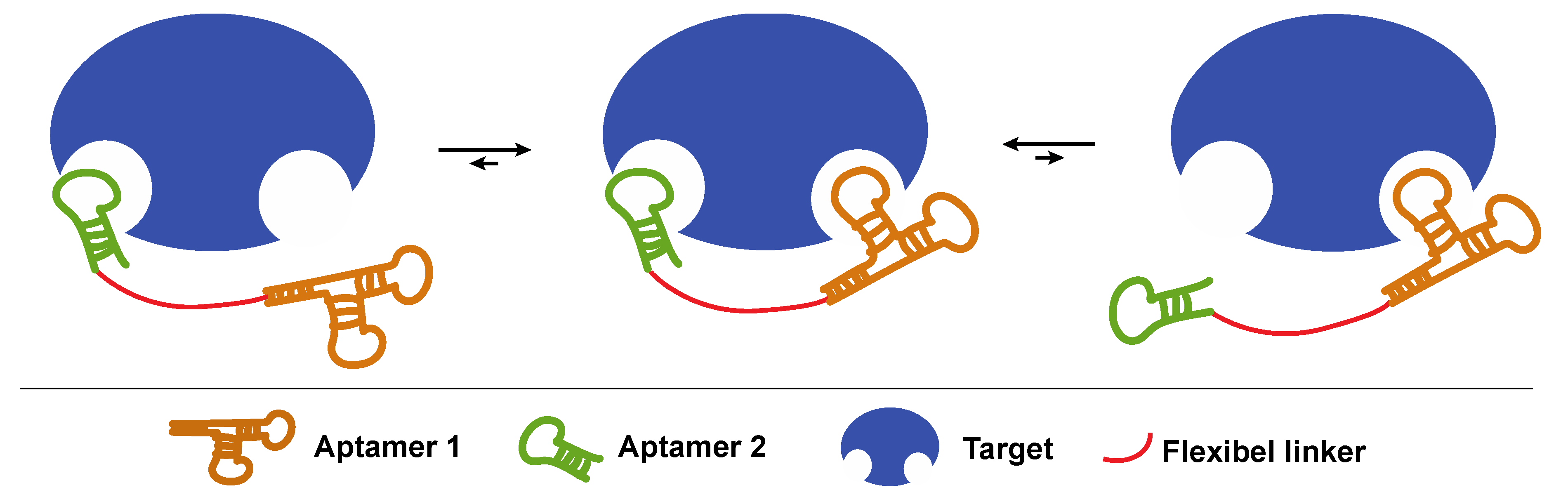
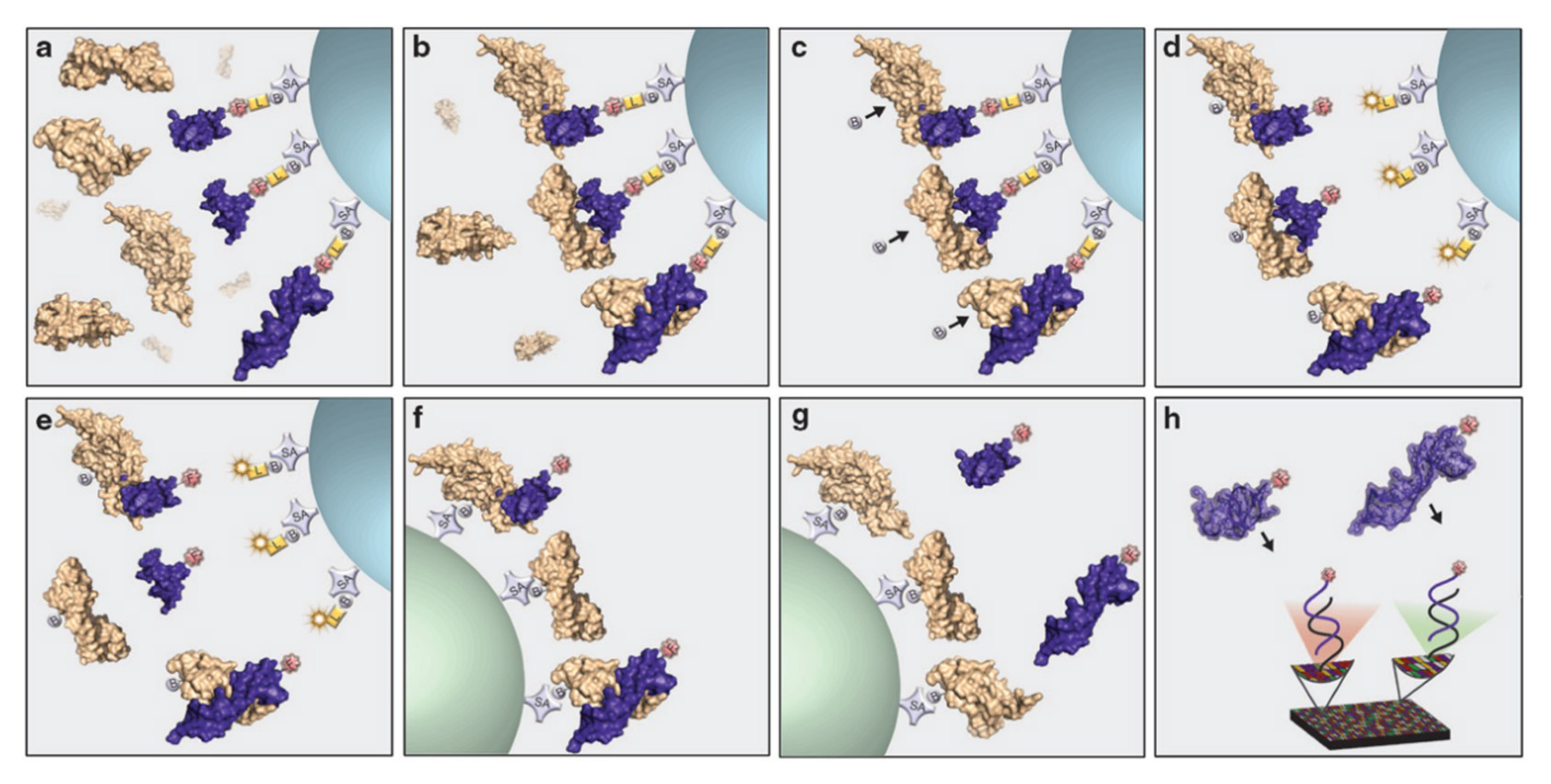
3.1.2. Joining Binding Motifs: Bivalent and Multivalent Aptamer Constructs
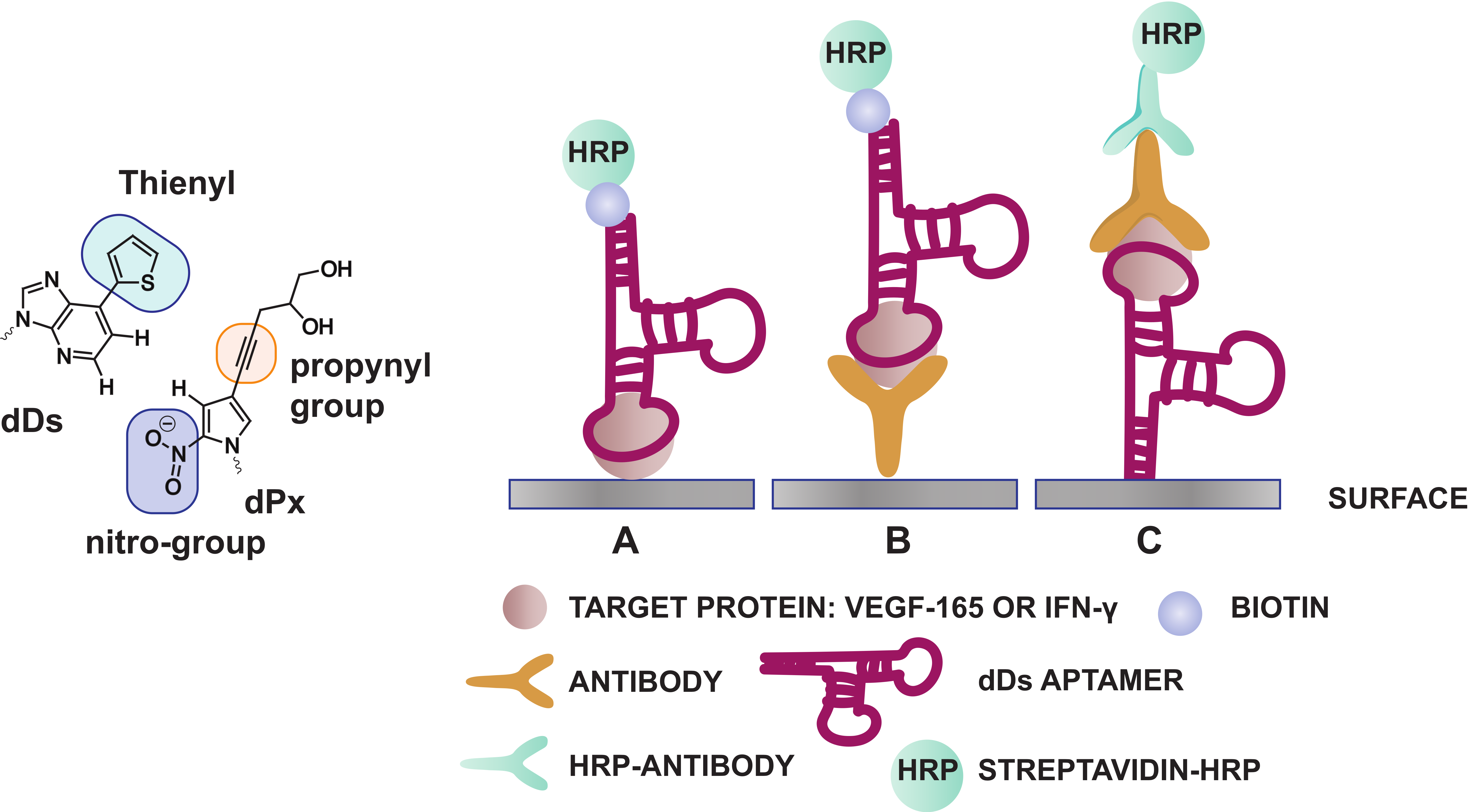
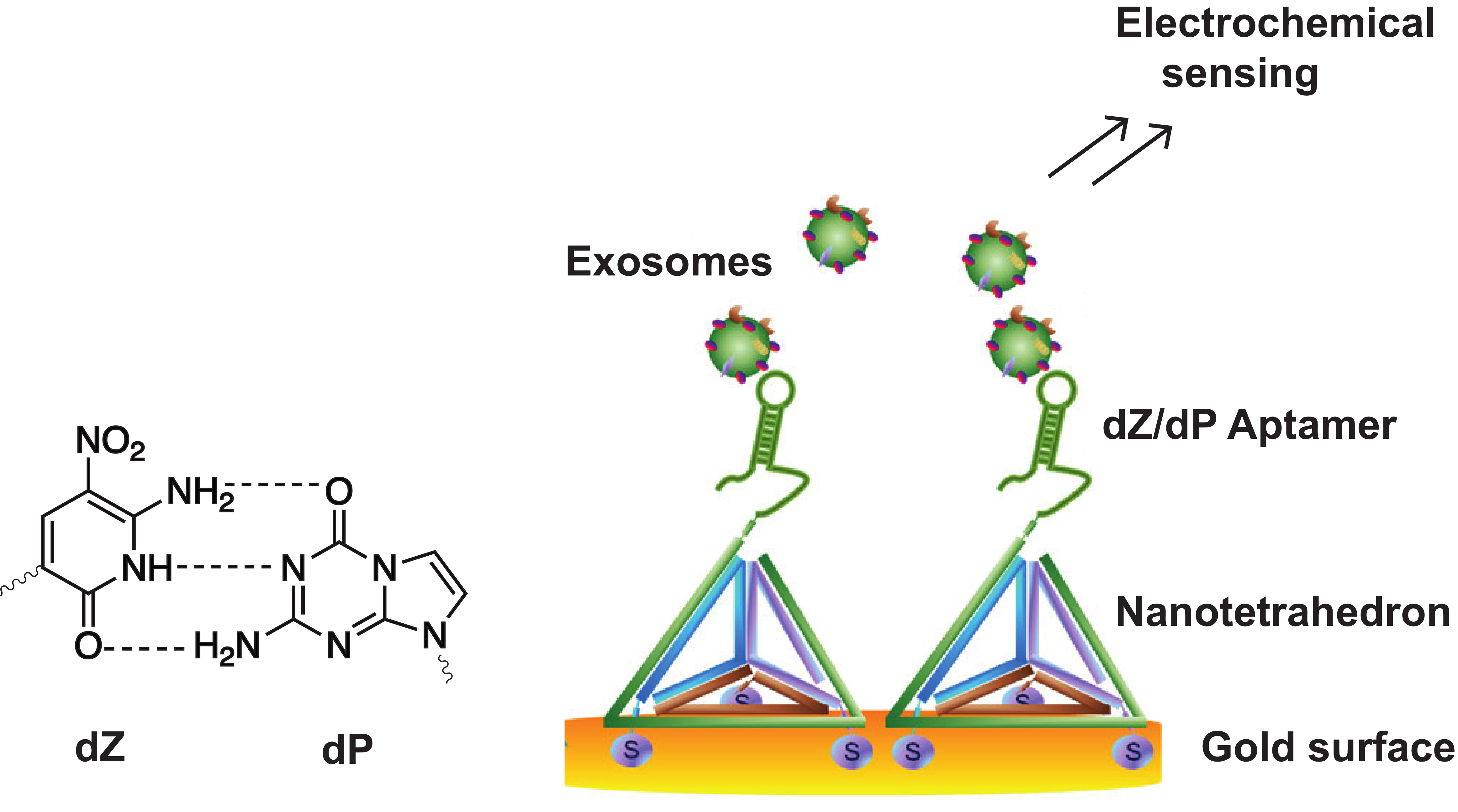
3.1.3. Exploiting the Extended Genetic Alphabet: Artificial Nucleobase Incorporation into Aptamers
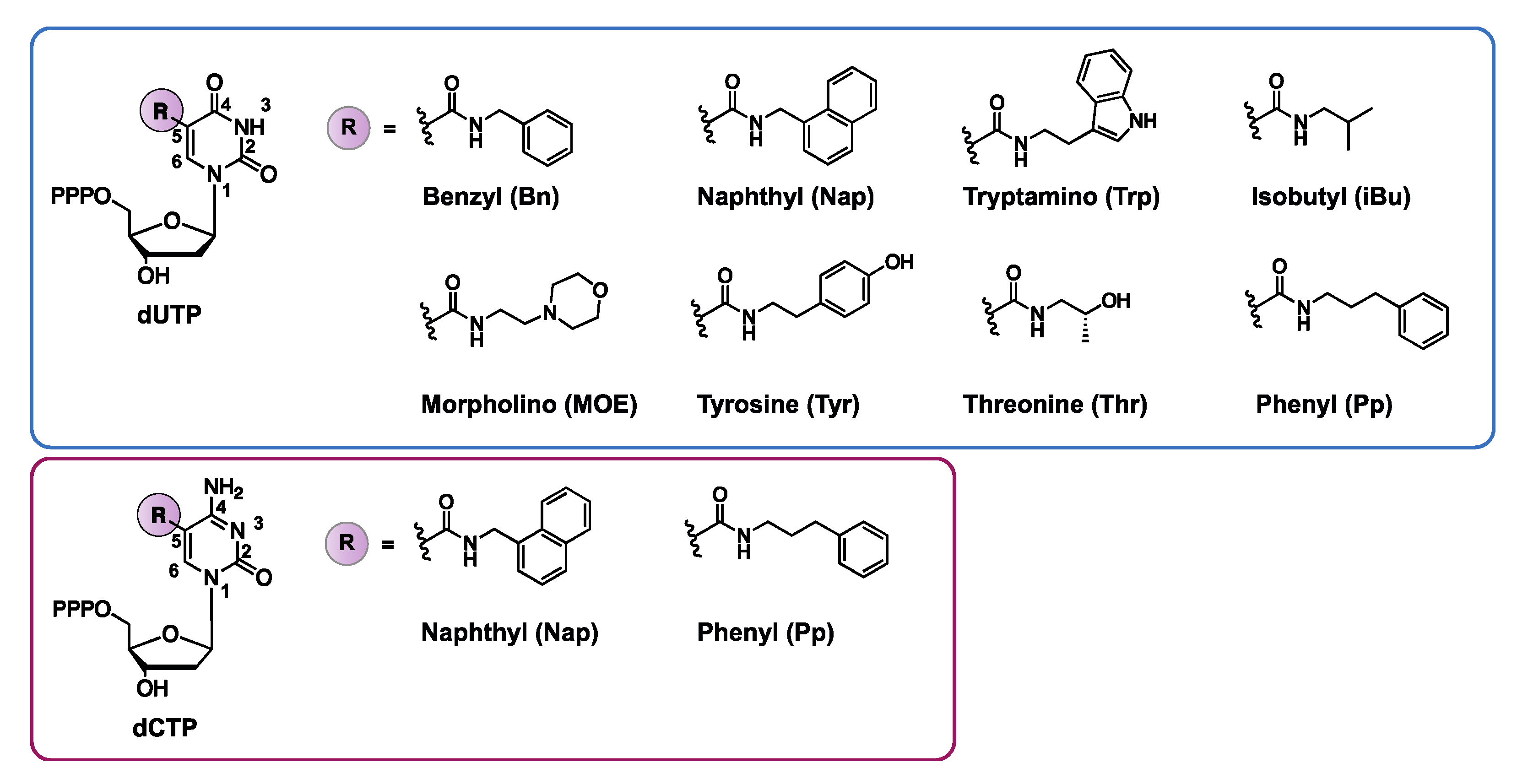
3.1.4. Nucleotides with Amino Acid Like Side Chains
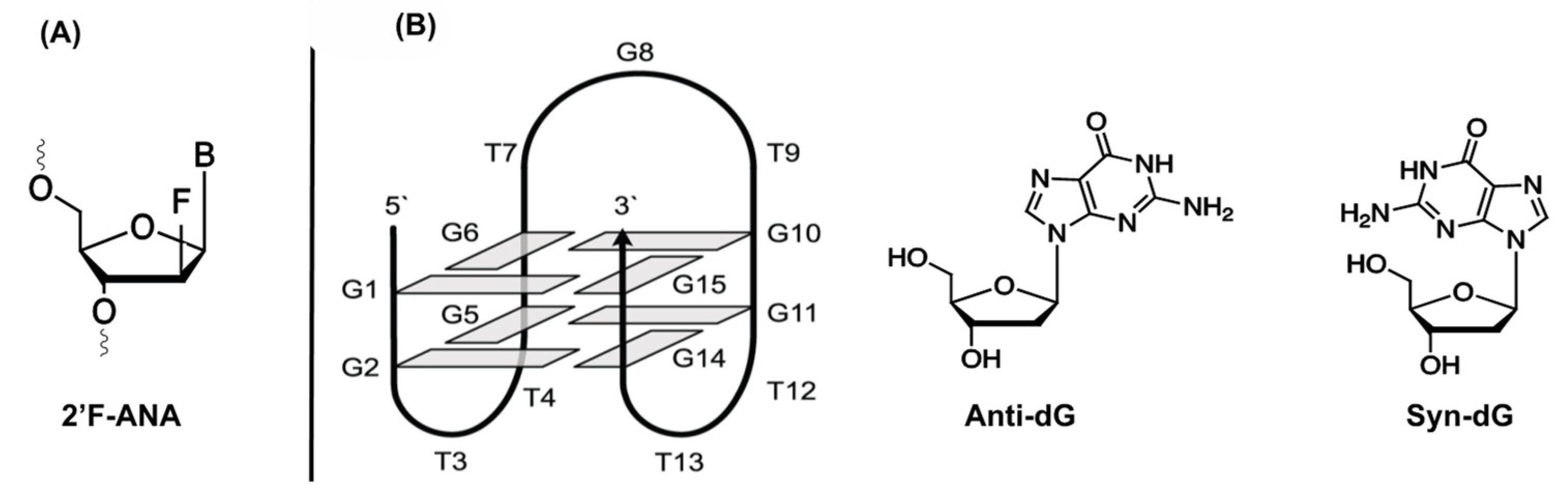
3.1.5. 2′-Fluoro Arabino Nucleic Acid (2′F-ANA)
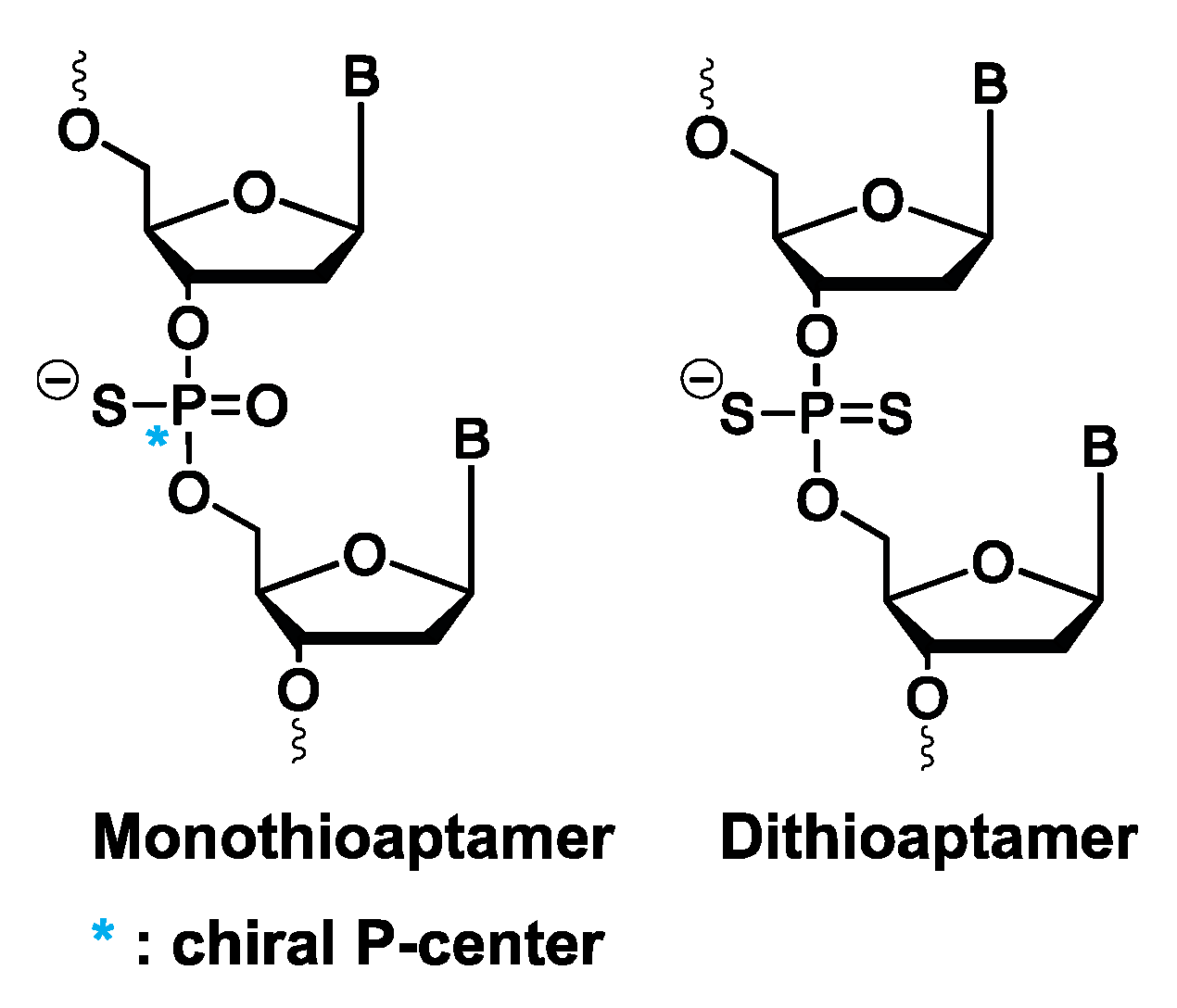
3.1.6. Mono- and Dithioaptamers
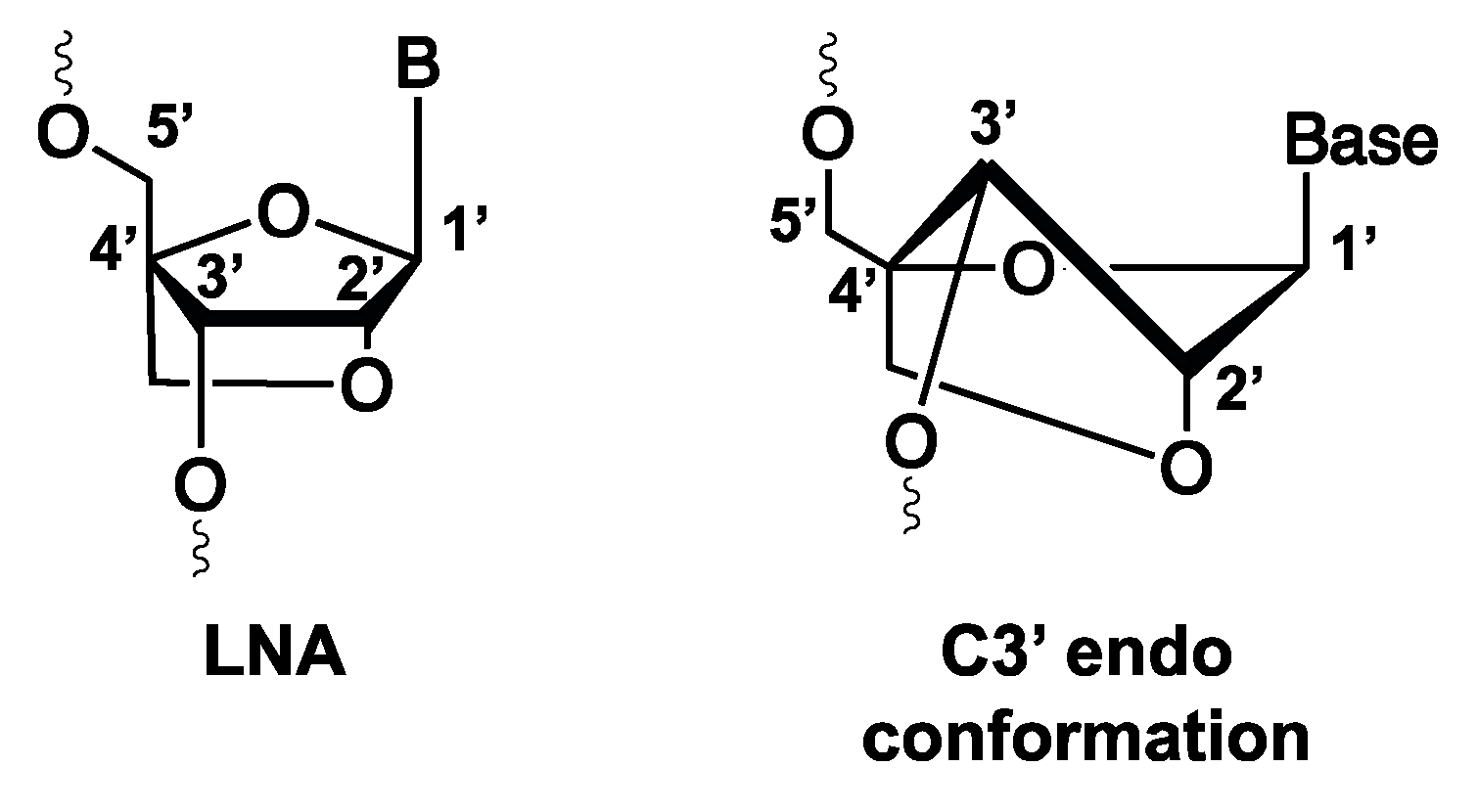
3.1.7. Locked Nucleic Acid (LNA) and Their Potential Usefulness for Diagnostic Tool Development
4. Modified Aptamers for Covalent Target Trapping
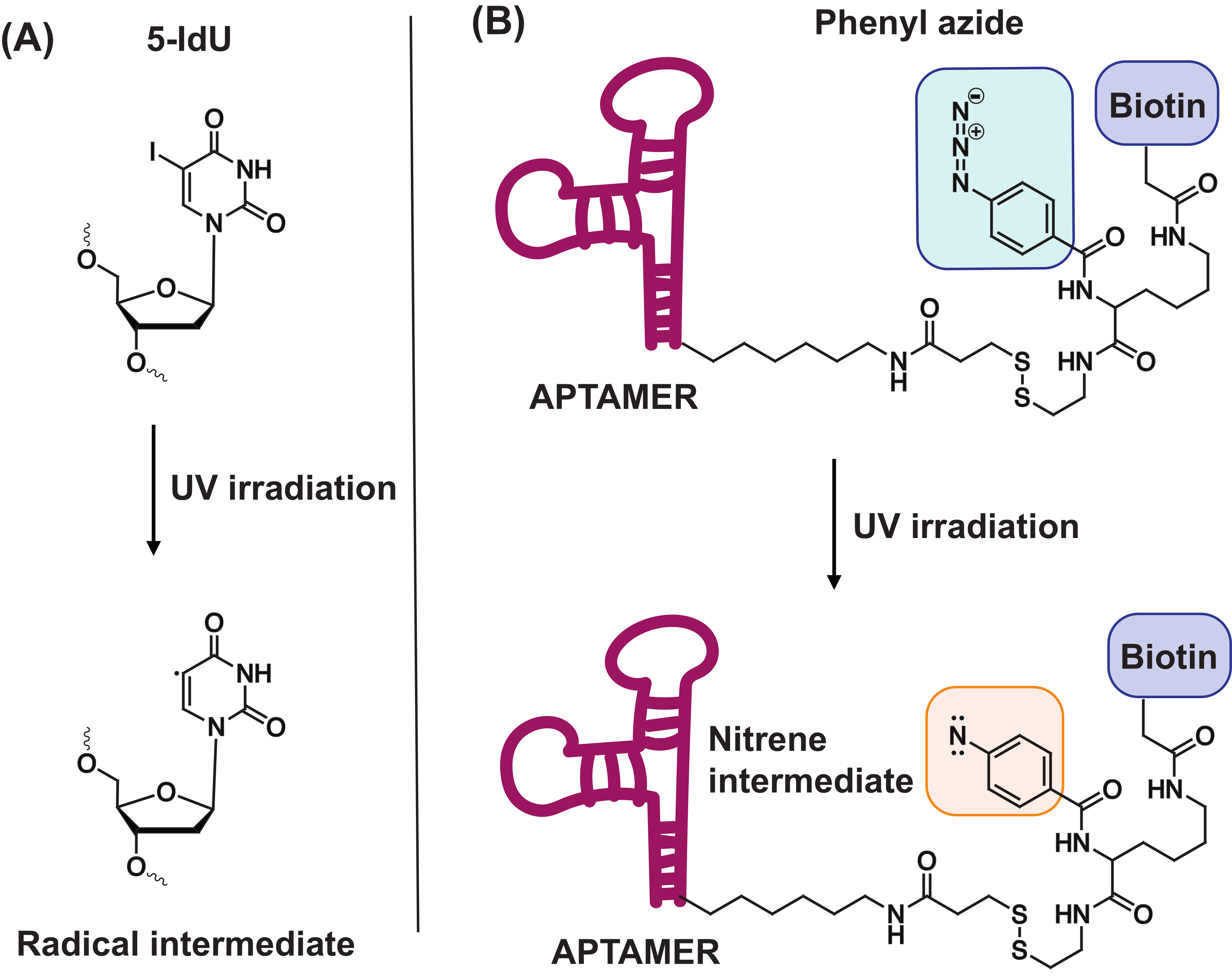
4.1. 5-Iodo-2′-Deoxyuridine (5-IdU)
4.2. Phenyl Azide-Modified Aptamers
4.3. Novel Crosslinking Aptamers with Future Potential for Diagnostic Tool Development
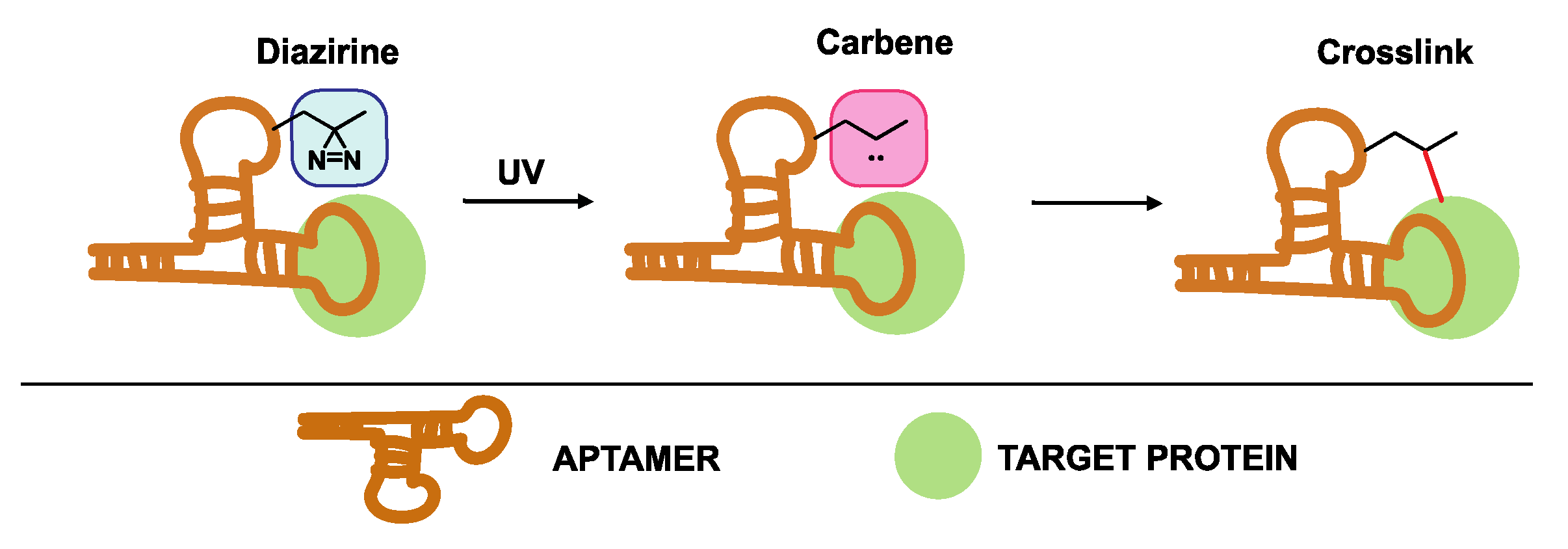
4.3.1. Diazirine-Modified Aptamers
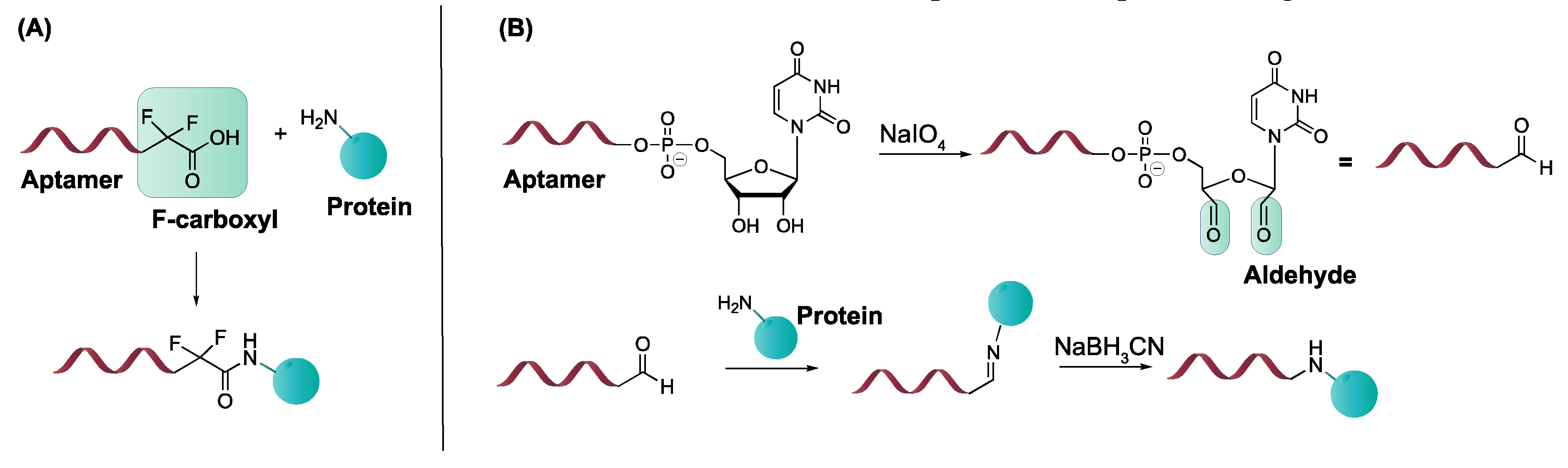
4.3.2. α, α-Gem-Difluoromethyl Carboxylic Acid (F-carboxyl) and Aldehyde-Modified Aptamers
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RNA | Ribonucleic acid |
| DNA | Deoxyribonucleic acid |
| A | Adenine |
| C | Cytosine |
| G | Guanine |
| T | Thymine |
| U | Uridine |
| ALISA | Aptamer-linked immobilized sorbent assays |
| KF | Folding constant |
| KD | Dissociation constant |
| SELEX | Synthetic evolution of ligands by exponential enrichment |
| PCR | Polymerase chain reaction |
| RT-PCR | Reverse transcription polymerase chain reaction |
| BLI | Biolayer interferometry |
| PE | r-phycoerythrin |
| FAM | Carboxyfluorescein |
| ELISA | Enzyme-linked immunosorbent assay |
| ELONA | Enzyme-linked oligonucleotide assay |
| DKK | Dickkopf |
| Muc-1 | Mucin-1 |
| dZ | 6-amino-5-nitro-3-(1′-β-D-2′-deoxyribofuranosyl)-2(1H)-pyridone |
| dP | 2-amino-8-(1′-β-D-2′-deoxyribofuranosyl)-imidazo[1,2-a]-1,3,5-triazin-4(8H)-one |
| Ds | 7-(2-thienyl)imidazo[4,5-b]pyridine |
| Px | 2-nitro-4-propynylpyrrole |
| PTK | Protein tyrosine kinase |
| Ag2S | Silver sulfide |
| VEGF | Vascular endothelial growth factor |
| IFN | Interferon |
| VWF | Von Willebrand Factor |
| pM | Picomolar |
| HRP | Horseradish peroxidase |
| mAb | Monoclonal antibody |
| dU | Deoxyuridine |
| dC | Deoxycytidine |
| dUTP | Deoxyuridines triphosphate |
| dCTP | Deoxycytidine triphosphate |
| Bn | 5-benzylaminocarbonyl |
| Nap | 5-naphthylmethylaminocarbonyl |
| Trp | 5-tryptaminocarbonyl |
| iBu | 5-isobutylaminocarbonyl |
| MOE | 5-morpholinoethylaminocarbonyl |
| Tyr | 5-hydroxyphenylethylaminocarboxyl |
| Thr | 5-hydroxyphenylethylaminocarboxyl |
| Pp | 5-phenylpropylaminocarbonyl |
| SOMAmer | Slow off-rate modified aptamers |
| UV | Ultraviolet |
| Simoa | Single molecule array |
| TNF | Tumor necrosis factor |
| GDP | Guanine diphosphate |
| HVP | Hydrolyzed vegetable protein |
| VLP | Virus-like particle |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NF-IL6 | Nuclear factor for interleukin-6 |
| HIV | Human immunodeficiency virus |
| TGF-β1 | Transforming growth factor beta 1 |
| RANK | Receptor activator of nuclear factor κB |
| SELDI- | Surface enhanced laser desorption ionization mass spectrometry |
| MS | Mass spectrometry |
| 2′-F-ANA | 2′-Fluoroarabino nucleic acid |
| TBA | Thrombin binding aptamer |
| 5-IdU | 5-iodo-2′-deoxyuridine |
| LNA | Locked nucleic acid |
| NaIO4 | Sodium periodate |
| NaBH3CN | Sodium cyanoborohydride |
| NHS | N-hydroxysuccinimide |
| IgG | Immunoglobulin G |
| F-carboxyl | α, α-gem-difluoromethyl carboxylic acid |
| SA | Streptavidin |
References
- Strehlitz, B.; Reinemann, C.; Linkorn, S.; Stoltenburg, R. Aptamers for pharmaceuticals and their application in environmental analytics. Bioanal. Rev. 2012, 4, 1–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.Y.; Acquah, C.; Sidhu, A.; Ongkudon, C.M.; Yon, L.S.; Danquah, M.K. SELEX modifications and bioanalytical techniques for aptamer–target binding characterization. Crit. Rev. Anal. Chem. 2016, 46, 521–537. [Google Scholar] [CrossRef] [PubMed]
- Hermann, T.; Patel, D.J. Adaptive recognition by nucleic acid aptamers. Science 2000, 287, 820–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biosensing, A.; Khan, N.I.; Song, E. Lab-on-a-chip systems for aptamer-based biosensing. Micromachines 2020, 11, 220. [Google Scholar] [CrossRef] [Green Version]
- Pilehvar, S.; Reinemann, C.; Bottari, F.; Vanderleyden, E.; Van Vlierberghe, S.; Blust, R.; Strehlitz, B.; De Wael, K. A joint action of aptamers and gold nanoparticles chemically trapped on a glassy carbon support for the electrochemical sensing of ofloxacin. Sens. Actuators B Chem. 2017, 240, 1024–1035. [Google Scholar] [CrossRef]
- Mehlhorn, A.; Rahimi, P.; Joseph, Y. Aptamer-based biosensors for antibiotic detection: A review. Biosensors 2018, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Scoville, D.J.; Uhm, T.K.B.; Shallcross, J.A.; Whelan, R.J. Selection of DNA aptamers for ovarian cancer biomarker CA125 using one-pot SELEX and high-throughput sequencing. J. Nucleic Acids 2017, 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Vivekananda, J.; Kiel, J.L. Anti-Francisella tularensis DNA aptamers detect tularemia antigen from different subspecies by Aptamer-Linked Immobilized Sorbent Assay. Lab. Investig. 2006, 86, 610–618. [Google Scholar] [CrossRef]
- Kawde, A.-N.; Xu, H.; Wang, S.; Liu, G.; Mao, X.; Zeng, Q. Aptamer-functionalized gold nanoparticles as probes in a dry-reagent strip biosensor for protein analysis. Anal. Chem. 2008, 81, 669–675. [Google Scholar] [CrossRef]
- Li, J.; Xu, M.; Huang, H.; Zhou, J.; Abdel-Halimb, E.S.; Zhang, J.R.; Zhu, J.J. Aptamer-quantum dots conjugates-based ultrasensitive competitive electrochemical cytosensor for the detection of tumor cell. Talanta 2011, 85, 2113–2120. [Google Scholar] [CrossRef]
- Dougherty, C.; Cai, W.; Hong, H. Applications of aptamers in targeted imaging: State of the art. Curr. Top. Med. Chem. 2015, 15, 1138–1152. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Yang, S. Replacing antibodies with aptamers in lateral flow immunoassay. Biosens. Bioelectron. 2015, 71, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, K.; Rana, K.V.; Suri, C.R. Characterization of hapten-protein conjugates: Antibody generation and immunoassay development for pesticides monitoring. Bionanoscience 2013, 3, 137–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoogenboom, H.R. Selecting and screening recombinant antibody libraries. Nat. Biotechnol. 2005, 23, 1105–1116. [Google Scholar] [CrossRef]
- Liu, C.W.; Chang, H.C.; Yang, R.J. Aptamer-based sensor for quantitative detection of mercury (II) ions by attenuated total reflection surface enhanced infrared absorption spectroscopy. Anal. Chim. Acta 2018, 1033, 137–147. [Google Scholar] [CrossRef]
- Baker, B.R.; Lai, R.Y.; Wood, M.S.; Doctor, E.H.; Heeger, A.J.; Plaxco, K.W. An electronic, aptamer-based small-molecule sensor for the rapid, label-free detection of cocaine in adulterated samples and biological fluids. J. Am. Chem. Soc. 2006, 128, 3138–3139. [Google Scholar] [CrossRef]
- Bates, P.J.; Reyes-Reyes, E.M.; Malik, M.T.; Murphy, E.M.; O’Toole, M.G.; Trent, J.O. G-quadruplex oligonucleotide AS1411 as a cancer-targeting agent: Uses and mechanisms. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1414–1428. [Google Scholar] [CrossRef]
- Zou, X.; Wu, J.; Gu, J.; Shen, L.; Mao, L. Application of aptamers in virus detection and antiviral therapy. Front. Microbiol. 2019, 10, 1462. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, R.; Wei, H.; Li, Y. Selection of aptamers against pathogenic bacteria and their diagnostics application. World J. Microbiol. Biotechnol. 2018, 34, 149. [Google Scholar] [CrossRef]
- Cerchia, L.; de Franciscis, V. Targeting cancer cells with nucleic acid aptamers. Trends Biotechnol. 2010, 28, 517–525. [Google Scholar] [CrossRef]
- Thiviyanathan, V.; Gorenstein, D.G. Aptamers and the next generation of diagnostic reagents. Proteomics Clin. Appl. 2012, 6, 563–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Ponniah, G.; Zhang, H.M.; Nowak, C.; Neill, A.; Gonzalez-Lopez, N.; Patel, R.; Cheng, G.; Kita, A.Z.; Andrien, B. In vitro and in vivo modifications of recombinant and human IgG antibodies. MABS 2014, 6, 1145–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, S.; Yao, H.; Wang, L.; Lu, J.; Jiang, F.; Lu, A.; Zhang, G. Chemical modifications of nucleic acid aptamers for therapeutic purposes. Int. J. Mol. Sci. 2017, 18, 1683. [Google Scholar] [CrossRef] [PubMed]
- Biphenyls, C.P. Aptamers as targeted therapeutics: Current potential and challenges. Nat. Rev. Drug Discov. 2015, 91, 165–171. [Google Scholar] [CrossRef]
- Morita, Y.; Leslie, M.; Kameyama, H.; Volk, D.E.; Tanaka, T. Aptamer therapeutics in cancer: Current and future. Cancers 2018, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Parashar, A. Aptamers in therapeutics. J. Clin. Diagn. Res. 2016, 10, BE01–BE06. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Marshall, K.A.; Ellington, A.D. In vitro selection of RNA aptamers. Methods Enzymol. 2000, 318, 193–214. [Google Scholar] [CrossRef]
- Stoltenburg, R.; Reinemann, C.; Strehlitz, B. SELEX-A (r)evolutionary method to generate high-affinity nucleic acid ligands. Biomol. Eng. 2007, 24, 381–403. [Google Scholar] [CrossRef]
- Pfeiffer, F.; Tolle, F.; Rosenthal, M.; Brändle, G.M.; Ewers, J.; Mayer, G. Identification and characterization of nucleobase-modified aptamers by click-SELEX. Nat. Protoc. 2018, 13, 1153–1180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lai, B.S.; Juhas, M. Recent advances in aptamer discovery and applications. Molecules 2019, 24, 941. [Google Scholar] [CrossRef] [Green Version]
- Bayat, P.; Nosrati, R.; Alibolandi, M.; Rafatpanah, H.; Abnous, K.; Khedri, M.; Ramezani, M. SELEX methods on the road to protein targeting with nucleic acid aptamers. Biochimie 2018, 154, 132–155. [Google Scholar] [CrossRef] [PubMed]
- Lapa, S.A.; Chudinov, A.V.; Timofeev, E.N. The toolbox for modified aptamers. Mol. Biotechnol. 2016, 58, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.R.; Gelinas, A.D.; Zhang, C.; Rohloff, J.C.; Carter, J.D.; O’Connell, D.; Waugh, S.M.; Wolk, S.K.; Mayfield, W.S.; Burgin, A.B.; et al. Unique motifs and hydrophobic interactions shape the binding of modified DNA ligands to protein targets. Proc. Natl. Acad. Sci. USA 2012, 109, 19971–19976. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Hirota, M.; Waugh, S.M.; Murakami, I.; Suzuki, T.; Muraguchi, M.; Shibamori, M.; Ishikawa, Y.; Jarvis, T.C.; Carter, J.D.; et al. Chemically modified DNA aptamers bind interleukin-6 with high affinity and inhibit signaling by blocking its interaction with interleukin-6 receptor. J. Biol. Chem. 2014, 289, 8706–8719. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Berdis, A.J. Non-natural nucleotides as probes for the mechanism and fidelity of DNA polymerases. Biochim. Biophys. Acta Proteins Proteom. 2010, 1804, 1064–1080. [Google Scholar] [CrossRef] [Green Version]
- Hendrickson, C.L.; Devine, K.G.; Benner, S.A. Probing minor groove recognition contacts by DNA polymerases and reverse transcriptases using 3-deaza-2’-deoxyadenosine. Nucleic Acids Res. 2004, 32, 2241–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, J.C.; Kool, E.T. Functional hydrogen-bonding map of the minor groove binding tracks of six DNA polymerases. Biochemistry 2000, 39, 12979–12988. [Google Scholar] [CrossRef]
- Pinheiro, V.B.; Holliger, P. Towards XNA nanotechnology: New materials from synthetic genetic polymers. Trends Biotechnol. 2014, 32, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Laos, R.; Thomson, J.M.; Benner, S.A. DNA polymerases engineered by directed evolution to incorporate nonstandard nucleotides. Front. Microbiol. 2014, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staiger, N.; Marx, A. A DNA polymerase with increased reactivity for ribonucleotides and C5-modified deoxyribonucleotides. ChemBioChem 2010, 11, 1963–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röthlisberger, P.; Hollenstein, M. Aptamer chemistry. Adv. Drug Deliv. Rev. 2018, 134, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Eaton, B.E.; Gold, L.; Hicke, B.J.; Janjić, N.; Jucker, F.M.; Sebesta, D.P.; Tarasow, T.M.; Willis, M.C.; Zichi, D.A. Post-SELEX combinatorial optimization of aptamers. Bioorg. Med. Chem. 1997, 5, 1087–1096. [Google Scholar] [CrossRef]
- Karlsen, K.K.; Wengel, J. Locked nucleic acid and aptamers. Nucleic Acid Ther. 2012, 22, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Cowperthwaite, M.C.; Ellington, A.D. Bioinformatic analysis of the contribution of primer sequences to aptamer structures. J. Mol. Evol. 2008, 67, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Yung, L.Y.L. Probing high affinity sequences of DNA aptamer against VEGF 165. PLoS ONE 2012, 7, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Hu, B.; Gao, S.X.; Liu, D.J.; Sun, M.J.; Jiao, B.H.; Wang, L.H. A saxitoxin-binding aptamer with higher affinity and inhibitory activity optimized by rational site-directed mutagenesis and truncation. Toxicon 2015, 101, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Hu, B.; Zheng, X.; Cao, Y.; Liu, D.; Sun, M.; Jiao, B.; Wang, L. Gonyautoxin 1/4 aptamers with high-affinity and high-specificity: From efficient selection to aptasensor application. Biosens. Bioelectron. 2016, 79, 938–944. [Google Scholar] [CrossRef]
- Soheili, V.; Taghdisi, S.M.; Hassanzadeh Khayyat, M.; Fazly Bazzaz, B.B.S.; Ramezani, M.; Abnous, K. Colorimetric and ratiometric aggregation assay for streptomycin using gold nanoparticles and a new and highly specific aptamer. Microchim. Acta 2016, 183, 1687–1697. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, Y.; Sheng, Z.; Li, T.; Li, X. A colorimetric detection method of pesticide acetamiprid by fine-tuning aptamer length. Anal. Biochem. 2016, 513, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Sha, J.; Li, Z.; Wang, W.; Zhang, H. High affinity truncated aptamers for ultra-sensitive colorimetric detection of bisphenol A with label-free aptasensor. Food Chem. 2020, 317, 126459. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Liu, S.; Shi, H.; Zhao, G. A highly sensitive and selective aptamer-based colorimetric sensor for the rapid detection of PCB 77. J. Hazard. Mater. 2018, 341, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhang, Y.; Nie, J.; Zhao, S.; Tian, Y.; Zhou, N. Gold nanoparticle based photometric determination of tobramycin by using new specific DNA aptamers. Microchim. Acta 2018, 185. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Yuan, L.; Jin, K.; Han, X.; Tian, Y.; Zhou, N. Electrochemical detection of tobramycin based on enzymes-assisted dual signal amplification by using a novel truncated aptamer with high affinity. Biosens. Bioelectron. 2018, 122, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, X.; He, J.; Hu, X.; Li, Y.; Li, X.; Fan, L.; Yu, H.Z. Systematic truncating of aptamers to create high-performance graphene oxide (GO)-based aptasensors for the multiplex detection of mycotoxins. Analyst 2019, 144, 3826–3835. [Google Scholar] [CrossRef]
- Alhadrami, H.A.; Chinnappan, R.; Eissa, S.; Rahamn, A.A.; Zourob, M. High affinity truncated DNA aptamers for the development of fluorescence based progesterone biosensors. Anal. Biochem. 2017, 525, 78–84. [Google Scholar] [CrossRef]
- Ha, N.R.; Jung, I.P.; La, I.J.; Jung, H.S.; Yoon, M.Y. Ultra-sensitive detection of kanamycin for food safety using a reduced graphene oxide-based fluorescent aptasensor. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.J.; Kim, A.R.; Yoon, M.Y.; You, Y.; Chua, B.; Son, A. Development of quantum dot aptasensor and its portable analyzer for the detection of di-2-ethylhexyl phthalate. Biosens. Bioelectron. 2018, 121, 1–9. [Google Scholar] [CrossRef]
- Sun, L.; Zhao, Q. Direct fluorescence anisotropy approach for aflatoxin B1 detection and affinity binding study by using single tetramethylrhodamine labeled aptamer. Talanta 2018, 189, 442–450. [Google Scholar] [CrossRef]
- McKeague, M.; Velu, R.; Hill, K.; Bardóczy, V.; Mészáros, T.; DeRosa, M.C. Selection and characterization of a novel DNA aptamer for label-free fluorescence biosensing of ochratoxin A. Toxins 2014, 6, 2435–2452. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Lim, H.J.; Lee, S.D.; Son, A. Highly sensitive detection of bisphenol a by nanoaptamer assay with truncated aptamer. ACS Appl. Mater. Interfaces 2017, 9, 14889–14898. [Google Scholar] [CrossRef] [PubMed]
- Chinnappan, R.; AlZabn, R.; Mir, T.A.; Bader, M.; Zourob, M. Fluorometric determination of okadaic acid using a truncated aptamer. Microchim. Acta 2019, 186, 406. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Duan, N.; Gu, H.; Wang, H.; Wang, Z. Fluorometric determination of lipopolysaccharides via changes of the graphene oxide-enhanced fluorescence polarization caused by truncated aptamers. Microchim. Acta 2019, 186, 173. [Google Scholar] [CrossRef]
- Chinnappan, R.; AlZabn, R.; Abu-Salah, K.M.; Zourob, M. An aptamer based fluorometric microcystin-LR assay using DNA strand-based competitive displacement. Microchim. Acta 2019, 186, 1–10. [Google Scholar] [CrossRef]
- Chinnappan, R.; Rahamn, A.A.; AlZabn, R.; Kamath, S.; Lopata, A.L.; Abu-Salah, K.M.; Zourob, M. Aptameric biosensor for the sensitive detection of major shrimp allergen, tropomyosin. Food Chem. 2020, 314, 126133. [Google Scholar] [CrossRef]
- Almusharraf, A.Y.; Eissa, S.; Zourob, M. Truncated aptamers for total and glycated hemoglobin, and their integration into a graphene oxide-based fluorometric method for high-throughput screening for diabetes. Microchim. Acta 2018, 185, 1–8. [Google Scholar] [CrossRef]
- Chinnappan, R.; AlAmer, S.; Eissa, S.; Rahamn, A.A.; Abu Salah, K.M.; Zourob, M. Fluorometric graphene oxide-based detection of Salmonella enteritis using a truncated DNA aptamer. Microchim. Acta 2018, 185, 61. [Google Scholar] [CrossRef]
- Bhamidipati, M.; Cho, H.Y.; Lee, K.B.; Fabris, L. SERS-based quantification of biomarker expression at the single cell level enabled by gold nanostars and truncated aptamers. Bioconjug. Chem. 2018, 29, 2970–2981. [Google Scholar] [CrossRef]
- Li, X.; Zhang, W.; Liu, L.; Zhu, Z.; Ouyang, G.; An, Y.; Zhao, C.; Yang, C.J. In vitro selection of DNA aptamers for metastatic breast cancer cell recognition and tissue imaging. Anal. Chem. 2014, 86, 6596–6603. [Google Scholar] [CrossRef]
- Li, W.; Wang, S.; Zhou, L.; Cheng, Y.; Fang, J. An ssDNA aptamer selected by Cell-SELEX for the targeted imaging of poorly differentiated gastric cancer tissue. Talanta 2019, 199, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, W.; Tseng, Y.; Zhang, J.; Liu, J. Developing slow-off dickkopf-1 aptamers for early-diagnosis of hepatocellular carcinoma. Talanta 2019, 194, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Stoltenburg, R.; Krafčiková, P.; Víglaský, V.; Strehlitz, B. G-quadruplex aptamer targeting Protein A and its capability to detect Staphylococcus aureus demonstrated by ELONA. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Lu, Z.; Jiang, H.; Yang, Z.; Liu, X.; Ding, H.; Li, H.; Dong, J.; Huang, A.; Fang, T.; et al. Aptamer selection and application in multivalent binding-based electrical impedance detection of inactivated H1N1 virus. Biosens. Bioelectron. 2018, 110, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Hoffman, B.E.; Lis, J.T. RNA aptamers as effective protein antagonists in a multicellular organism. Proc. Natl. Acad. Sci. USA 1999, 96, 10033–10038. [Google Scholar] [CrossRef] [Green Version]
- Vorobyeva, M.; Vorobjev, P.; Venyaminova, A. Multivalent aptamers: Versatile tools for diagnostic and therapeutic applications. Molecules 2016, 21, 1613. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Hu, J.; Bezerra, A.B.; Holtan, M.D.; Brooks, J.C.; Easley, C.J. Protein quantification using controlled DNA melting transitions in bivalent probe assemblies. Anal. Chem. 2015, 87, 9576–9579. [Google Scholar] [CrossRef] [Green Version]
- Sheng, W.; Chen, T.; Tan, W.; Fan, Z.H. Multivalent DNA nanospheres for enhanced capture of cancer cells in microfluidic devices. ACS Nano 2013, 7, 7067–7076. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Tyagi, D.; Lyu, M.; Carrier, A.J.; Nganou, C.; Youden, B.; Wang, W.; Cui, S.; Servos, M.; Oakes, K.; et al. Regenerative nanooctopus based on multivalent-aptamer-functionalized magnetic microparticles for effective cell capture in whole blood. Anal. Chem. 2019, 91, 4017–4022. [Google Scholar] [CrossRef]
- Ding, C.; Zhang, C.; Cheng, S.; Xian, Y. Multivalent aptamer functionalized Ag2S nanodots/hybrid cell membrane-coated magnetic nanobioprobe for the ultrasensitive isolation and detection of circulating tumor cells. Adv. Funct. Mater. 2020, 1909781. [Google Scholar] [CrossRef]
- Chen, T.; Hongdilokkul, N.; Liu, Z.; Thirunavukarasu, D.; Romesberg, F.E. The expanding world of DNA and RNA. Curr. Opin. Chem. Biol. 2016, 34, 80–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimoto, M.; Hikida, Y.; Hirao, I. Site-specific functional labeling of nucleic acids by in vitro replication and transcription using unnatural base pair systems. Isr. J. Chem. 2013, 53, 450–468. [Google Scholar] [CrossRef]
- Yang, Z.; Sismour, A.M.; Sheng, P.; Puskar, N.L.; Benner, S.A. Enzymatic incorporation of a third nucleobase pair. Nucleic Acids Res. 2007, 35, 4238–4249. [Google Scholar] [CrossRef] [PubMed]
- Kimoto, M.; Yamashige, R.; Matsunaga, K.I.; Yokoyama, S.; Hirao, I. Generation of high-affinity DNA aptamers using an expanded genetic alphabet. Nat. Biotechnol. 2013, 31, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, K.; Kimoto, M.; Hirao, I. High-affinity DNA aptamer generation targeting von willebrand factor a1-domain by genetic alphabet expansion for systematic evolution of ligands by exponential enrichment using two types of libraries composed of five different bases. J. Am. Chem. Soc. 2017, 139, 324–334. [Google Scholar] [CrossRef]
- Kimoto, M.; Shermane Lim, Y.W.; Hirao, I. Molecular affinity rulers: Systematic evaluation of DNA aptamers for their applicabilities in ELISA. Nucleic Acids Res. 2019, 47, 8362–8374. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhang, L.; Wan, S.; Cansiz, S.; Cui, C.; Liu, Y.; Cai, R.; Hong, C.; Teng, I.-T.; Shi, M.; et al. Aptasensor with expanded nucleotide using DNA nanotetrahedra for electrochemical detection of cancerous exosomes. ACS Nano 2017, 11, 3943–3949. [Google Scholar] [CrossRef]
- Glushakova, L.G.; Sharma, N.; Hoshika, S.; Bradley, A.C.; Bradley, K.M.; Yang, Z.; Benner, S.A. Detecting respiratory viral RNA using expanded genetic alphabets and self-avoiding DNA. Anal. Biochem. 2015, 489, 62–72. [Google Scholar] [CrossRef]
- Glushakova, L.G.; Bradley, A.; Bradley, K.M.; Alto, B.W.; Hoshika, S.; Hutter, D.; Sharma, N.; Yang, Z.; Kim, M.J.; Benner, S.A. High-throughput multiplexed xMAP Luminex array panel for detection of twenty two medically important mosquito-borne arboviruses based on innovations in synthetic biology. J. Virol. Methods 2015, 214, 60–74. [Google Scholar] [CrossRef] [Green Version]
- Hoshika, S.; Leal, N.A.; Kim, M.J.; Kim, M.S.; Karalkar, N.B.; Kim, H.J.; Bates, A.M.; Watkins, N.E.; SantaLucia, H.A.; Meyer, A.J.; et al. Hachimoji DNA and RNA: A genetic system with eight building blocks. Science 2019, 363, 884–887. [Google Scholar] [CrossRef]
- Malyshev, D.A.; Dhami, K.; Lavergne, T.; Chen, T.; Dai, N.; Foster, J.M.; Corrêa, I.R.; Romesberg, F.E. A semi-synthetic organism with an expanded genetic alphabet. Nature 2014, 509, 385–388. [Google Scholar] [CrossRef] [Green Version]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T.; et al. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 5, e15004. [Google Scholar] [CrossRef] [Green Version]
- Gelinas, A.D.; Davies, D.R.; Edwards, T.E.; Rohloff, J.C.; Carter, J.D.; Zhang, C.; Gupta, S.; Ishikawa, Y.; Hirota, M.; Nakaishi, Y.; et al. Crystal structure of interleukin-6 in complex with a modified nucleic acid ligand. J. Biol. Chem. 2014, 289, 8720–8734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gawande, B.N.; Rohloff, J.C.; Carter, J.D.; Von Carlowitz, I.; Zhang, C.; Schneider, D.J.; Janjic, N. Selection of DNA aptamers with two modified bases. Proc. Natl. Acad. Sci. USA 2017, 114, 2898–2903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlShamaileh, H.; Veedu, R.N. Next-generation nucleic acid aptamers with two-base-modified nucleotides have improved binding affinity and potency. ChemBioChem 2017, 18, 1565–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brody, E.N.; Gold, L.; Lawn, R.M.; Walker, J.J.; Zichi, D. High-content affinity-based proteomics: Unlocking protein biomarker discovery. Expert Rev. Mol. Diagn. 2010, 10, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Katilius, E.; Olivas, E.; Dumont Milutinovic, M.; Walt, D.R. Incorporation of slow off-rate modified aptamers reagents in single molecule array assays for cytokine detection with ultrahigh sensitivity. Anal. Chem. 2016, 88, 8385–8389. [Google Scholar] [CrossRef] [Green Version]
- Gold, L.; Walker, J.J.; Wilcox, S.K.; Williams, S. Advances in human proteomics at high scale with the SOMAscan proteomics platform. New Biotechnol. 2012, 29, 543–549. [Google Scholar] [CrossRef]
- Rohloff, J.C.; Gelinas, A.D.; Jarvis, T.C.; Ochsner, U.A.; Schneider, D.J.; Gold, L.; Janjic, N. Nucleic acid ligands with protein-like side chains: Modified aptamers and their use as diagnostic and therapeutic agents. Mol. Ther. Nucleic Acids 2014, 3, e201. [Google Scholar] [CrossRef]
- Ostroff, R.M.; Mehan, M.R.; Stewart, A.; Ayers, D.; Brody, E.N.; Williams, S.A.; Levin, S.; Black, B.; Harbut, M.; Carbone, M.; et al. Early detection of malignant pleural mesothelioma in asbestos-exposed individuals with a noninvasive proteomics-based surveillance tool. PLoS ONE 2012, 7, e46091. [Google Scholar] [CrossRef]
- Webber, J.; Stone, T.C.; Katilius, E.; Smith, B.C.; Gordon, B.; Mason, M.D.; Tabi, Z.; Brewis, I.A.; Clayton, A. Proteomics analysis of cancer exosomes using a novel modified aptamer-based array (somascanTM) platform. Mol. Cell. Proteom. 2014, 13, 1050–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehan, M.R.; Williams, S.A.; Siegfried, J.M.; Bigbee, W.L.; Weissfeld, J.L.; Wilson, D.O.; Pass, H.I.; Rom, W.N.; Muley, T.; Meister, M.; et al. Validation of a blood protein signature for non-small cell lung cancer. Clin. Proteom. 2014, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Thirstrup, D.; Jarvis, T.C.; Schneider, D.J.; Wilcox, S.K.; Carter, J.; Zhang, C.; Gelinas, A.; Weiss, A.; Janjic, N.; et al. Rapid histochemistry using slow off-rate modified aptamers with anionic competition. Appl. Immunohistochem. Mol. Morphol. 2011, 19, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Tsim, S.; Kelly, C.; Alexander, L.; McCormick, C.; Thomson, F.; Woodward, R.; Foster, J.E.; Stobo, D.B.; Paul, J.; Maskell, N.A.; et al. Diagnostic and Prognostic Biomarkers in the Rational Assessment of Mesothelioma (DIAPHRAGM) study: Protocol of a prospective, multicentre, observational study. BMJ Open 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Deterding, R.R.; Wagner, B.D.; Kirk Harris, J.; DeBoer, E.M. Pulmonary aptamer signatures in children’s interstitial and diffuse lung disease. Am. J. Respir. Crit. Care Med. 2019, 200, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- DeBoer, E.M.; Kroehl, M.E.; Wagner, B.D.; Accurso, F.J.; Harris, J.K.; Lynch, D.A.; Sagel, S.D.; Deterding, R.R. Proteomic profiling identifies novel circulating markers associated with bronchiectasis in cystic fibrosis. Proteom. Clin. Appl. 2017, 11, 9–10. [Google Scholar] [CrossRef]
- Hathout, Y.; Brody, E.; Clemens, P.R.; Cripe, L.; DeLisle, R.K.; Furlong, P.; Gordish-Dressman, H.; Hache, L.; Henricson, E.; Hoffman, E.P.; et al. Large-scale serum protein biomarker discovery in Duchenne muscular dystrophy. Proc. Natl. Acad. Sci. USA 2015, 112, 7153–7158. [Google Scholar] [CrossRef] [Green Version]
- Kiddle, S.J.; Sattlecker, M.; Proitsi, P.; Simmons, A.; Westman, E.; Bazenet, C.; Nelson, S.K.; Williams, S.; Hodges, A.; Johnston, C.; et al. Candidate blood proteome markers of Alzheimer’s disease onset and progression: A systematic review and replication study. J. Alzheimer’s Dis. 2014, 38, 515–531. [Google Scholar] [CrossRef] [Green Version]
- De Groote, M.A.; Nahid, P.; Jarlsberg, L.; Johnson, J.L.; Weiner, M.; Muzanyi, G.; Janjic, N.; Sterling, D.G.; Ochsner, U.A. Elucidating novel serum biomarkers associated with pulmonary tuberculosis treatment. PLoS ONE 2013, 8, e61002. [Google Scholar] [CrossRef] [Green Version]
- De Groote, M.A.; Sterling, D.G.; Hraha, T.; Russell, T.M.; Green, L.S.; Wall, K.; Kraemer, S.; Ostroff, R.; Janjic, N.; Ochsner, U.A. Discovery and validation of a six-marker serum protein signature for the diagnosis of active pulmonary tuberculosis. J. Clin. Microbiol. 2017, 55, 3057–3071. [Google Scholar] [CrossRef] [Green Version]
- De Groote, M.A.; Higgins, M.; Hraha, T.; Wall, K.; Wilson, M.L.; Sterling, D.G.; Janjic, N.; Reves, R.; Ochsner, U.A.; Belknap, R. Highly multiplexed proteomic analysis of quantiferon supernatants to identify biomarkers of latent tuberculosis infection. J. Clin. Microbiol. 2017, 55, 391–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, T.M.; Green, L.S.; Rice, T.; Kruh-Garcia, N.A.; Dobos, K.; De Groote, M.A.; Hraha, T.; Sterling, D.G.; Janjic, N.; Ochsner, U.A. Potential of high-affinity, slow off-rate modified aptamer reagents for Mycobacterium tuberculosis proteins as tools for infection models and diagnostic applications. J. Clin. Microbiol. 2017, 55, 3072–3088. [Google Scholar] [CrossRef] [Green Version]
- Nahid, P.; Bliven-Sizemore, E.; Jarlsberg, L.G.; De Groote, M.A.; Johnson, J.L.; Muzanyi, G.; Engle, M.; Weiner, M.; Janjic, N.; Sterling, D.G.; et al. Aptamer-based proteomic signature of intensive phase treatment response in pulmonary tuberculosis. Tuberculosis 2014, 94, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Altieri, A.; Piyadasa, H.; Recksiedler, B.; Spicer, V.; Mookherjee, N. Cytokines IL-17, TNF and IFN-γ alter the expression of antimicrobial peptides and proteins disparately: A targeted proteomics analysis using SOMAscan technology. Vaccines 2018, 6, 51. [Google Scholar] [CrossRef] [Green Version]
- Hamula, C.L.A.; Zhang, H.; Li, F.; Wang, Z.; Chris Le, X.; Li, X.F. Selection and analytical applications of aptamers binding microbial pathogens. Trends Anal. Chem. 2011, 30, 1587–1597. [Google Scholar] [CrossRef]
- Desai, V.G.; Lee, T.; Moland, C.L.; Vijay, V.; Han, T.; Lewis, S.M.; Herman, E.H.; Fuscoe, J.C. Candidate early predictive plasma protein markers of doxorubicin-induced chronic cardiotoxicity in B6C3F1 mice. Toxicol. Appl. Pharmacol. 2019, 363, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.; Dwivedi, A.K.; Deodhar, S.; Mishra, I.; Cistola, D.P. Aptamer-based search for correlates of plasma and serum water T 2: Implications for early metabolic dysregulation and metabolic syndrome 11 Medical and Health Sciences 1103 Clinical Sciences. Biomark. Res. 2018, 6, 1–12. [Google Scholar] [CrossRef]
- Baird, G.S.; Nelson, S.K.; Keeney, T.R.; Stewart, A.; Williams, S.; Kraemer, S.; Peskind, E.R.; Montine, T.J. Age-dependent changes in the cerebrospinal fluid proteome by slow off-rate modified aptamer array. Am. J. Pathol. 2012, 180, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Kukova, L.Z.; Mansour, S.G.; Coca, S.G.; de Fontnouvelle, C.A.; Thiessen-Philbrook, H.R.; Shlipak, M.G.; El-Khoury, J.M.; Parikh, C.R. Comparison of urine and plasma biomarker concentrations measured by aptamer-based versus immunoassay methods in cardiac surgery patients. J. Appl. Lab. Med. 2019, 4, 331–342. [Google Scholar] [CrossRef]
- Aghaeepour, N.; Lehallier, B.; Baca, Q.; Ganio, E.A.; Wong, R.J.; Ghaemi, M.S.; Culos, A.; El-Sayed, Y.Y.; Blumenfeld, Y.J.; Druzin, M.L.; et al. A proteomic clock of human pregnancy. Am. J. Obstet. Gynecol. 2018, 218, 347. [Google Scholar] [CrossRef] [Green Version]
- Ochsner, U.A.; Katilius, E.; Janjic, N. Detection of Clostridium difficile toxins A, B and binary toxin with slow off-rate modified aptamers. Diagn. Microbiol. Infect. Dis. 2013, 76, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Green, L.S.; Gold, L.; Janjic, N. Systematic selection of modified aptamer pairs for diagnostic sandwich assays. Biotechniques 2014, 56, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Green, L.S.; Rice, T.P.; Olivas, E.; Janjic, N.; Katilius, E. Targeting unique epitopes on highly similar proteins GDF-11 and GDF-8 with modified DNA aptamers. Biochemistry 2019, 58, 4632–4640. [Google Scholar] [CrossRef] [PubMed]
- Trausch, J.J.; Shank-Retzlaff, M.; Verch, T. Development and characterization of an HPV Type-16 specific modified DNA aptamer for the improvement of potency assays. Anal. Chem. 2017, 89, 3554–3561. [Google Scholar] [CrossRef] [PubMed]
- Eid, C.; Palko, J.W.; Katilius, E.; Santiago, J.G. Rapid slow off-rate modified aptamer (SOMAmer)-based detection of c-reactive protein using isotachophoresis and an ionic spacer. Anal. Chem. 2015, 87, 6736–6743. [Google Scholar] [CrossRef]
- Escolano, J.M.; Díaz-Durán, B.; DeMiguel-Ramos, M.; Olivares, J.; Geday, M.A.; Iborra, E. Selection of aptamers to Neisseria meningitidis and Streptococcus pneumoniae surface specific proteins and affinity assay using thin film AlN resonators. Sens. Actuators B Chem. 2017, 246, 591–596. [Google Scholar] [CrossRef]
- Katilius, E.; Carmel, A.B.; Koss, H.; O’Connell, D.; Smith, B.C.; Sanders, G.M.; LaBerge, G.S. Sperm cell purification from mock forensic swabs using SOMAmerTM affinity reagents. Forensic Sci. Int. Genet. 2018, 35, 9–13. [Google Scholar] [CrossRef]
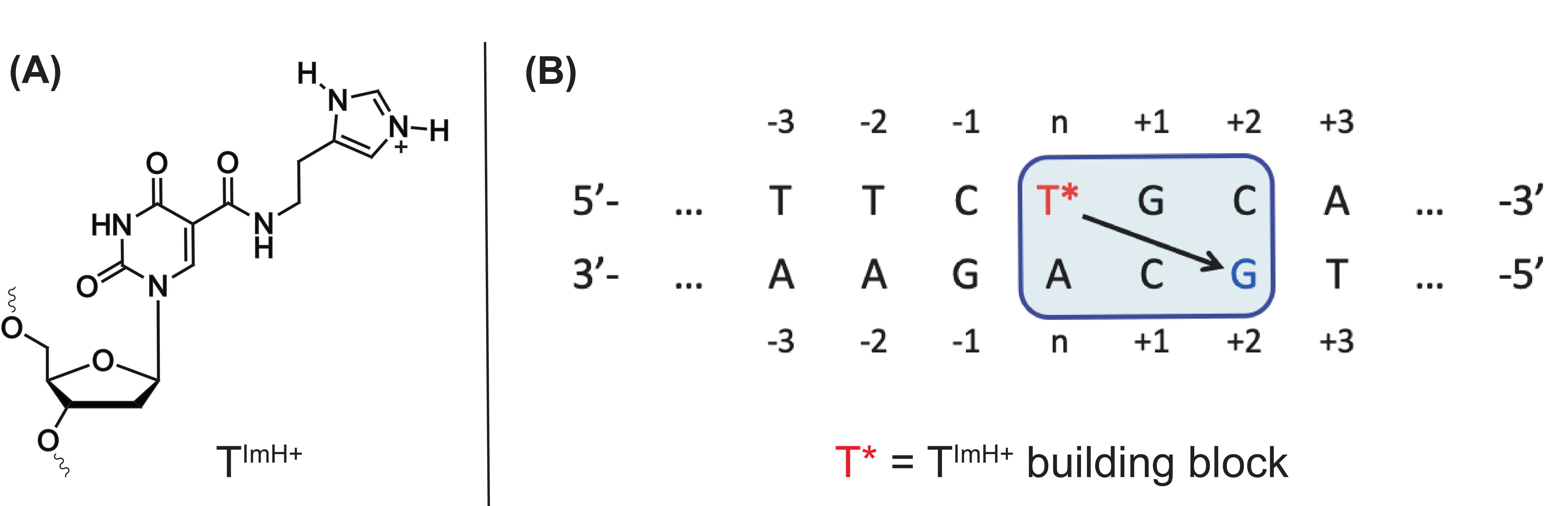
- Buyst, D.; Gheerardijn, V.; Fehér, K.; Van Gasse, B.; Van Den Begin, J.; Martins, J.C.; Madder, A. Identification of a pKa-regulating motif stabilizing imidazole-modified double-stranded DNA. Nucleic Acids Res. 2015, 43, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Verdonck, L.; Buyst, D.; de Vries, A.-M.; Gheerardijn, V.; Madder, A.; Martins, J.C. Tethered imidazole mediated duplex stabilization and its potential for aptamer stabilization. Nucleic Acids Res. 2018, 46, 11671–11686. [Google Scholar] [CrossRef]
- Röthlisberger, P.; Levi-Acobas, F.; Sarac, I.; Marlière, P.; Herdewijn, P.; Hollenstein, M. On the enzymatic incorporation of an imidazole nucleotide into DNA. Org. Biomol. Chem. 2017, 15, 4449–4455. [Google Scholar] [CrossRef]
- Hollenstein, M. Nucleoside triphosphates-building blocks for the modification of nucleic acids. Molecules 2012, 17, 13569–13591. [Google Scholar] [CrossRef] [PubMed]
- Hollenstein, M. Nucleic acid enzymes based on functionalized nucleosides. Curr. Opin. Chem. Biol. 2019, 52, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Roxo, C.; Kotkowiak, W.; Pasternak, A. G-quadruplex-forming aptamers—Characteristics, applications, and perspectives. Molecules 2019, 24, 3781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, C.G.; Damha, M.J. G-quadruplex induced stabilization by 2’-deoxy-2’-fluoro-d-arabinonucleic acids (2’F-ANA). Nucleic Acids Res. 2007, 35, 4977–4988. [Google Scholar] [CrossRef]
- Lietard, J.; Assi, H.A.; Gómez-Pinto, I.; González, C.; Somoza, M.M.; Damha, M.J. Mapping the affinity landscape of Thrombin-binding aptamers on 2F-ANA/DNA chimeric G-Quadruplex microarrays. Nucleic Acids Res. 2017, 45, 1619–1632. [Google Scholar] [CrossRef] [Green Version]
- Rose, K.M.; Alves Ferreira-Bravo, I.; Li, M.; Craigie, R.; Ditzler, M.A.; Holliger, P.; Destefano, J.J. Selection of 2’-Deoxy-2’-fluoroarabino nucleic acid (FANA) aptamers that bind HIV-1 integrase with picomolar affinity. ACS Chem. Biol. 2019, 14, 2166–2175. [Google Scholar] [CrossRef]
- Yang, X.; Gorenstein, D. Progress in thioaptamer development. Curr. Drug Targets 2004, 5, 705–715. [Google Scholar] [CrossRef]
- Volk, D.E.; Power, T.D.; Gorenstein, D.G.; Luxon, B.A. An ab initio study of phosphorothioate and phosphorodithioate interactions with sodium cation. Tetrahedron Lett. 2002, 43, 4443–4447. [Google Scholar] [CrossRef]
- King, D.J.; Ventura, D.A.; Brasier, A.R.; Gorenstein, D.G. Novel combinatorial selection of phosphorothioate oligonucleotide aptamers. Biochemistry 1998, 37, 16489–16493. [Google Scholar] [CrossRef]
- Yang, X. Construction and selection of bead-bound combinatorial oligonucleoside phosphorothioate and phosphorodithioate aptamer libraries designed for rapid PCR-based sequencing. Nucleic Acids Res. 2002, 30, 132e. [Google Scholar] [CrossRef] [Green Version]
- Leary, J.F.; Reece, L.M.; Yang, X.-B.; Gorenstein, D. High-throughput flow cytometric screening of combinatorial chemistry bead libraries for proteomics and drug discovery. Adv. Biomed. Clin. Diagn. Syst. III 2005, 5692, 216. [Google Scholar] [CrossRef]
- Yang, X.; Fennewald, S.; Luxon, B.A.; Aronson, J.; Herzog, N.K.; Gorenstein, D.G. Aptamers containing thymidine 3’-O-phosphorodithioates: Synthesis and binding to nuclear factor-κB. Bioorg. Med. Chem. Lett. 1999, 9, 3357–3362. [Google Scholar] [CrossRef]
- Thiviyanathan, V.; Somasunderam, A.D.; Gorenstein, D.G. Combinatorial selection and delivery of thioaptamers. Biochem. Soc. Trans. 2007, 35, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Lee, M.S.; Copland, J.A.; Luxon, B.A.; Gorenstein, D.G. Combinatorial selection of a single stranded DNA thioaptamer targeting TGF-β1 protein. Bioorg. Med. Chem. Lett. 2008, 18, 1835–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andras, J.; Simon, L.; Redlich, K.; Meszaros, T.; Mandl, P. A4.14 Selection and characterisation of rank specific thioaptamers. Ann. Rheum. Dis. 2015, 74, 42. [Google Scholar] [CrossRef]
- Wang, H.; Yang, X.; Bowick, G.C.; Herzog, N.K.; Luxon, B.A.; Lomas, L.O.; Gorenstein, D.G. Identification of proteins bound to a thioaptamer probe on a proteomics array. Biochem. Biophys. Res. Commun. 2006, 347, 586–593. [Google Scholar] [CrossRef]
- Matharu, Z.; Patel, D.; Gao, Y.; Haque, A.; Zhou, Q.; Revzin, A. Detecting transforming growth factor-β release from liver cells using an aptasensor integrated with microfluidics. Anal. Chem. 2014, 86, 8865–8872. [Google Scholar] [CrossRef] [Green Version]
- Hecht, A.H.; Sommer, G.J.; Durland, R.H.; Yang, X.; Singh, A.K.; Hatch, A.V. Aptamers as affinity reagents in an integrated electrophoretic lab-on-a-chip platform. Anal. Chem. 2010, 82, 8813–8820. [Google Scholar] [CrossRef]
- Koshkin, A.A.; Singh, S.K.; Nielsen, P.; Rajwanshi, V.K.; Kumar, R.; Meldgaard, M.; Olsen, C.E.; Wengel, J. LNA (Locked Nucleic Acids): Synthesis of the adenine, cytosine, guanine, 5-methylcytosine, thymine and uracil bicyclonucleoside monomers, oligomerisation, and unprecedented nucleic acid recognition. Tetrahedron 1998, 54, 3607–3630. [Google Scholar] [CrossRef]
- Elle, I.C.; Karlsen, K.K.; Terp, M.G.; Larsen, N.; Nielsen, R.; Derbyshire, N.; Mandrup, S.; Ditzel, H.J.; Wengel, J. Selection of LNA-containing DNA aptamers against recombinant human CD73. Mol. Biosyst. 2015, 11, 1260–1270. [Google Scholar] [CrossRef] [Green Version]
- Bonifacio, L.; Church, F.C.; Jarstfer, M.B. Effect of locked-nucleic acid on a biologically active G-quadruplex. A structure-activity relationship of the thrombin aptamer. Int. J. Mol. Sci. 2008, 9, 422–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, F.J.; Kalra, N.; Wengel, J.; Vester, B. Aptamers as a model for functional evaluation of LNA and 2’-amino LNA. Bioorg. Med. Chem. Lett. 2009, 19, 6585–6587. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, A.S.; Hansen, L.H.; Vester, B.; Wengel, J. Improvement of a streptavidin-binding aptamer by LNA- and α-l-LNA-substitutions. Bioorg. Med. Chem. Lett. 2014, 24, 2273–2277. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, D.; Tang, Z.; Mallikaratchy, P.; Xiao, Z.; Tan, W. Optimization and modifications of aptamers selected from live cancer cell lines. ChemBioChem 2007, 8, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Mallikaratchy, P.R.; Ruggiero, A.; Gardner, J.R.; Kuryavyi, V.; Maguire, W.F.; Heaney, M.L.; McDevitt, M.R.; Patel, D.J.; Scheinberg, D.A. A multivalent DNA aptamer specific for the B-cell receptor on human lymphoma and leukemia. Nucleic Acids Res. 2011, 39, 2458–2469. [Google Scholar] [CrossRef]
- Juskowiak, B. Nucleic acid-based fluorescent probes and their analytical potential. Anal. Bioanal. Chem. 2011, 399, 3157–3176. [Google Scholar] [CrossRef] [Green Version]
- Hamaguchi, N.; Ellington, A.; Stanton, M. Aptamer beacons for the direct detection of proteins. Anal. Biochem. 2001, 294, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Liy, Z.; Lechy, C.J.; Phan, A.T. Sugar-modified G-quadruplexes: Effects of LNA-, 2’F-RNA-and 2’F-ANA-guanosine chemistries on G-quadruplex structure and stability. Nucleic Acids Res. 2014, 42, 4068–4079. [Google Scholar] [CrossRef]
- Bi, W.; Bai, X.; Gao, F.; Lu, C.; Wang, Y.; Zhai, G.; Tian, S.; Fan, E.; Zhang, Y.; Zhang, K. DNA-templated aptamer probe for identification of target proteins. Anal. Chem. 2017, 89, 4071–4076. [Google Scholar] [CrossRef]
- Mallikaratchy, P.; Tang, Z.; Kwame, S.; Meng, L.; Shangguan, D.; Tan, W. Aptamer directly evolved from live cells recognizes membrane bound immunoglobin heavy mu chain in Burkitt’s lymphoma cells. Mol. Cell. Proteom. 2007, 6, 2230–2238. [Google Scholar] [CrossRef] [Green Version]
- Vinkenborg, J.L.; Mayer, G.; Famulok, M. Aptamer-based affinity labeling of proteins. Angew. Chem. Int. Ed. 2012, 51, 9176–9180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Song, Y.; Zou, Y.; Ge, Y.; An, Y.; Ma, Y.; Zhu, Z.; Yang, C.J. A diazirine-based photoaffinity probe for facile and efficient aptamer-protein covalent conjugation. Chem. Commun. 2014, 50, 4891–4894. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Lu, D.; Bai, H.; Jin, C.; Yan, G.; Ye, M.; Qiu, L.; Chang, R.; Cui, C.; Liang, H.; et al. Using modified aptamers for site specific protein-aptamer conjugations. Chem. Sci. 2016, 7, 2157–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
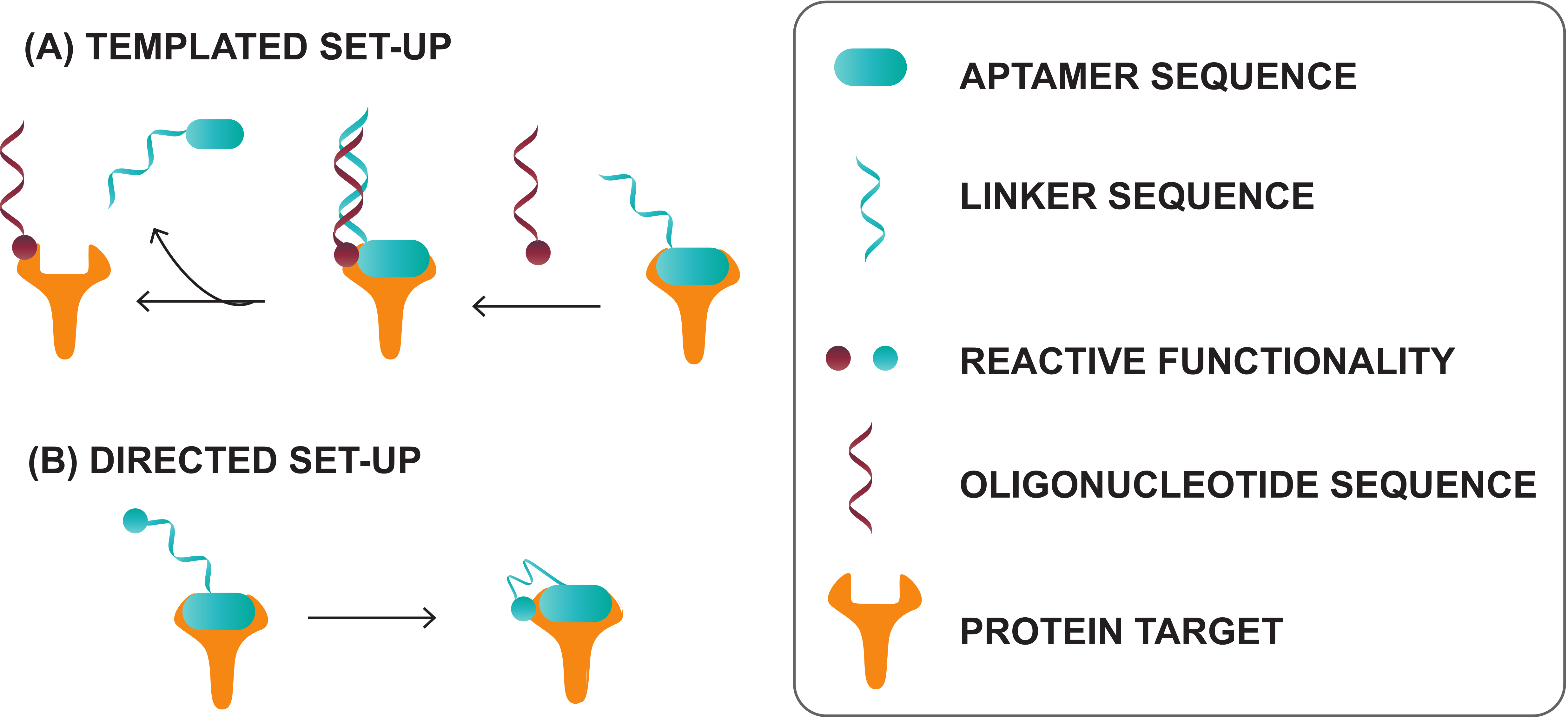
- Skovsgaard, M.B.; Mortensen, M.R.; Palmfeldt, J.; Gothelf, K.V. Aptamer-directed conjugation of DNA to therapeutic antibodies. Bioconjug. Chem. 2019, 30, 2127–2135. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modifications For Non-Covalent Target Binding | |||
| C5 modified nucleotides |  | dZ-dP |  |
| dDs-dPx |  | Thioaptamer |  |
| 2′ F-ANA |  | LNA |  |
| Modifications For Covalent Target Trapping | |||
| 5′-IdU |  | Diazirine |  |
| Aldehyde |  | F-carboxyl |  |
| Phenyl azide |  | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elskens, J.P.; Elskens, J.M.; Madder, A. Chemical Modification of Aptamers for Increased Binding Affinity in Diagnostic Applications: Current Status and Future Prospects. Int. J. Mol. Sci. 2020, 21, 4522. https://doi.org/10.3390/ijms21124522
Elskens JP, Elskens JM, Madder A. Chemical Modification of Aptamers for Increased Binding Affinity in Diagnostic Applications: Current Status and Future Prospects. International Journal of Molecular Sciences. 2020; 21(12):4522. https://doi.org/10.3390/ijms21124522
Chicago/Turabian StyleElskens, Jan P., Joke M. Elskens, and Annemieke Madder. 2020. "Chemical Modification of Aptamers for Increased Binding Affinity in Diagnostic Applications: Current Status and Future Prospects" International Journal of Molecular Sciences 21, no. 12: 4522. https://doi.org/10.3390/ijms21124522