Preeclampsia-Like Features and Partial Lactation Failure in Mice Lacking Cystathionine γ-Lyase—An Animal Model of Cystathioninuria


Abstract
:1. Introduction
2. Results
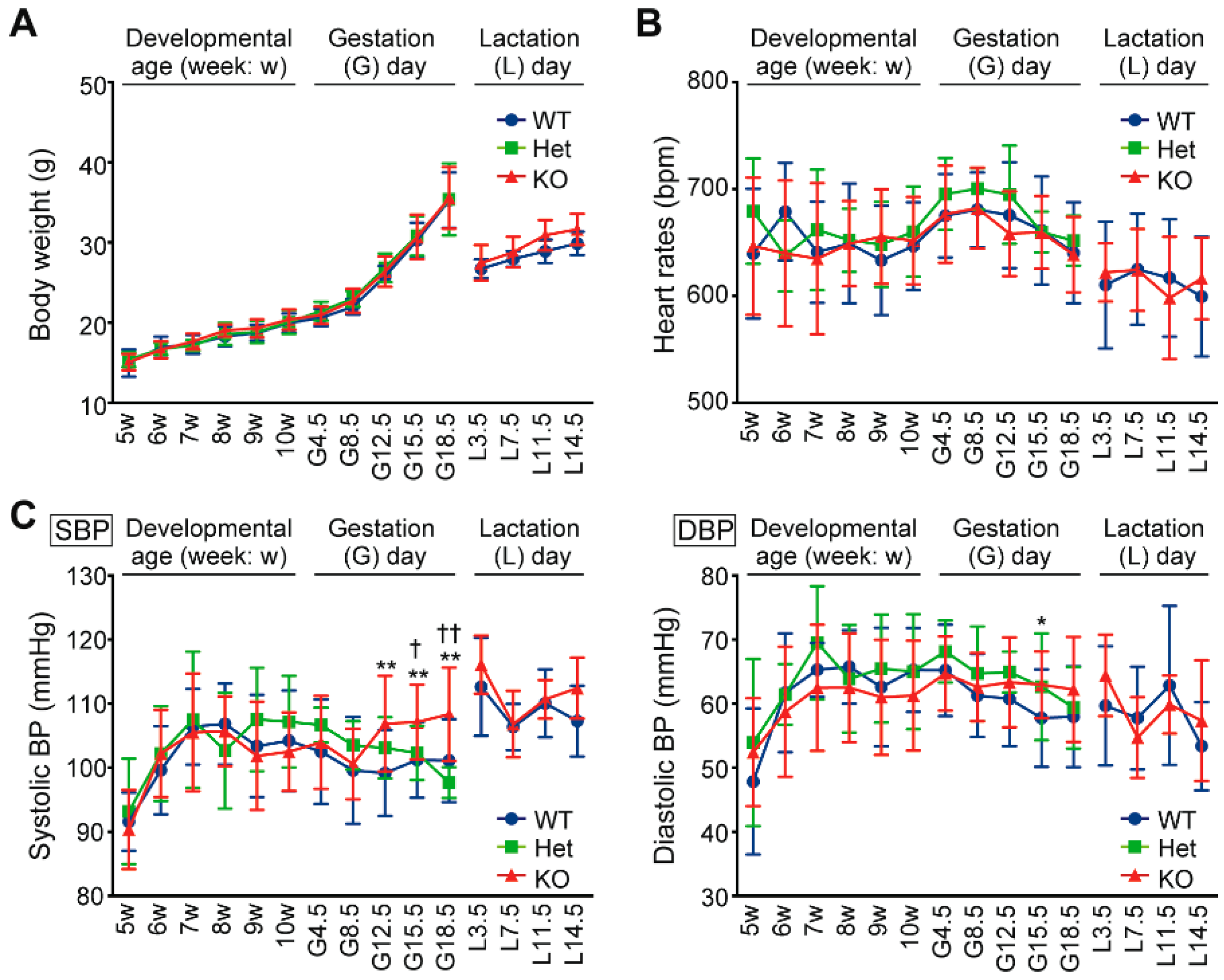
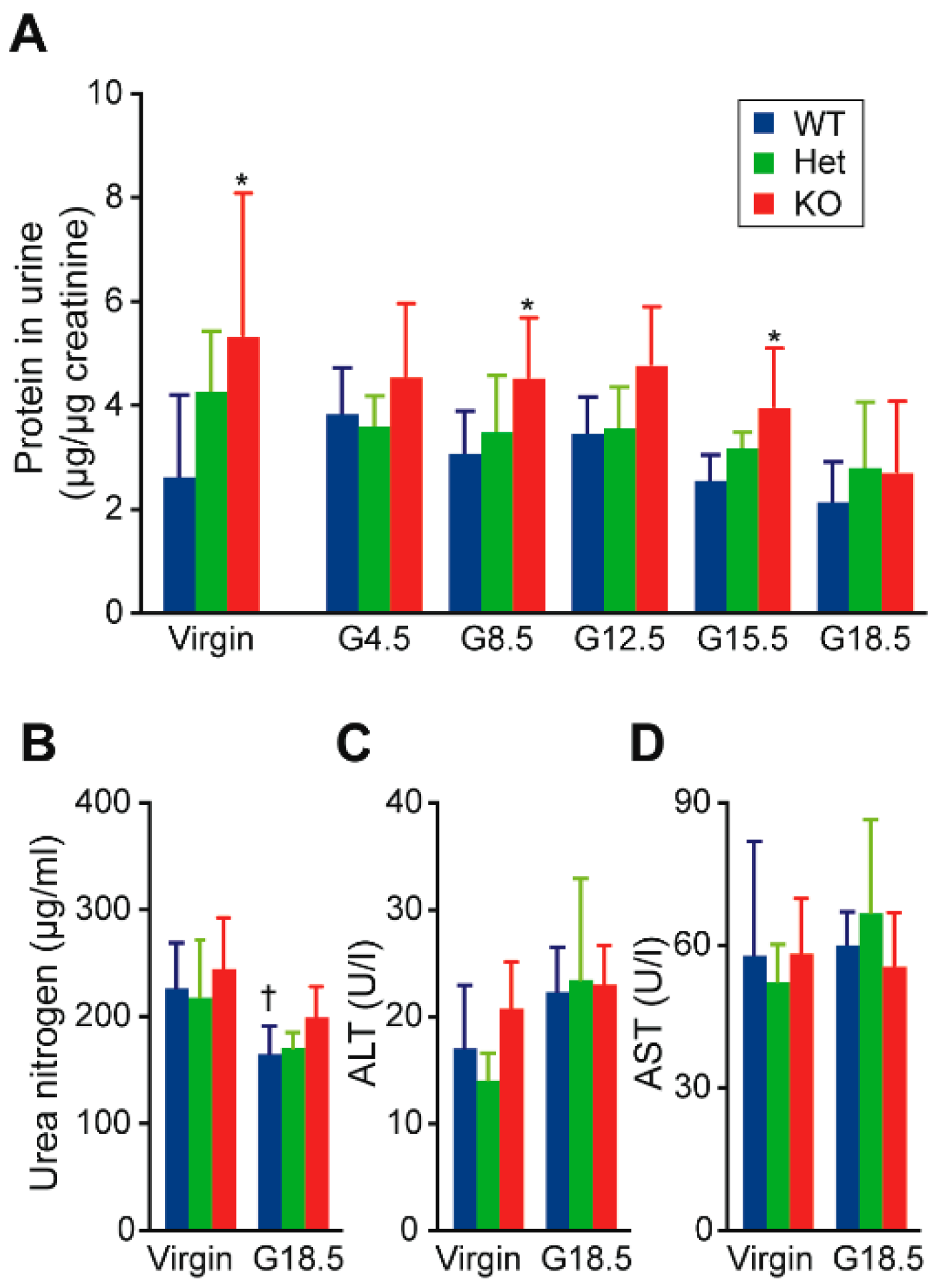
2.1. Hypertension and Proteinuria During Late Gestation Periods in Cth−/− Dams
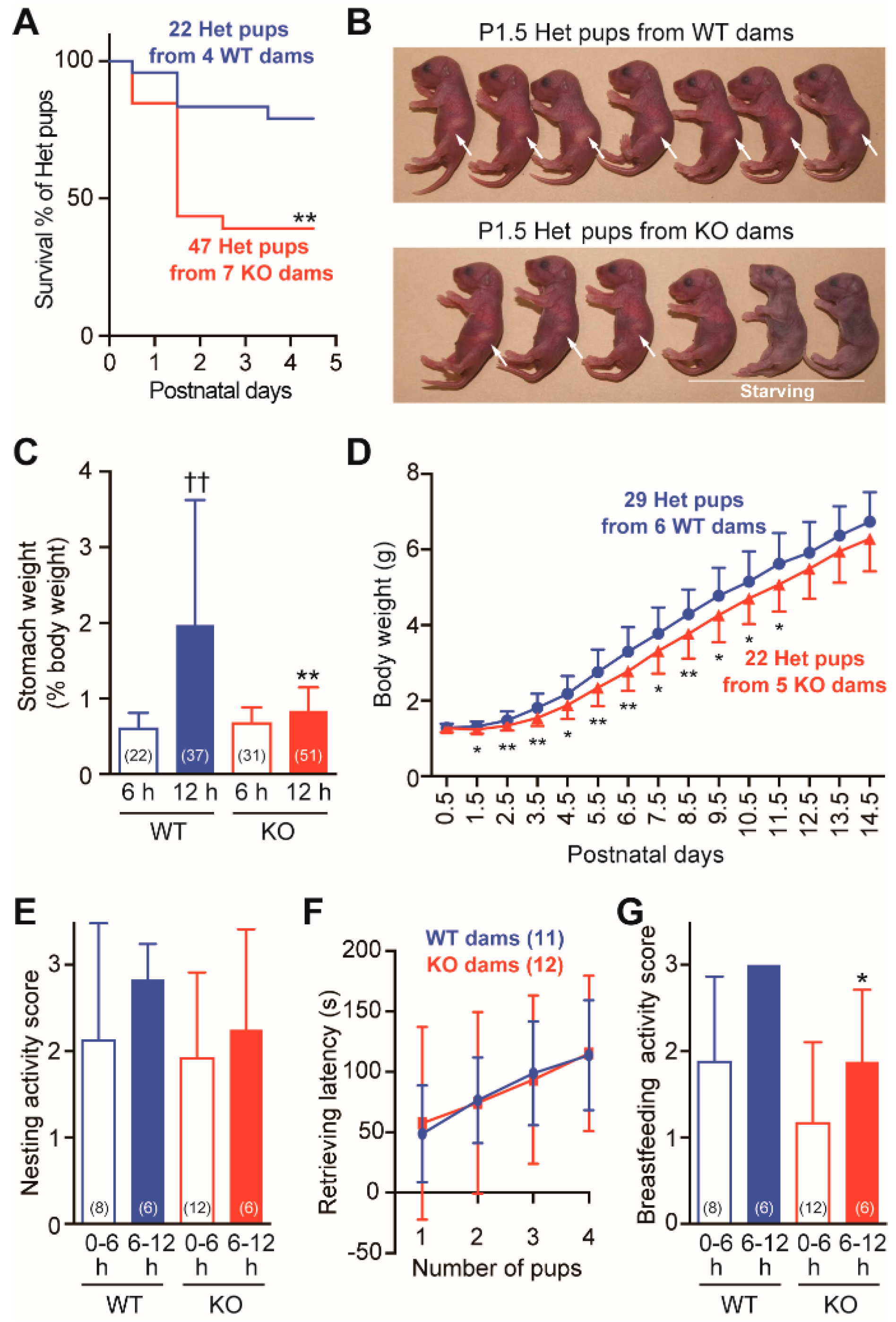
2.2. Frequent Neonatal Deaths in Cth+/− Pups Born to Cth−/− Dams
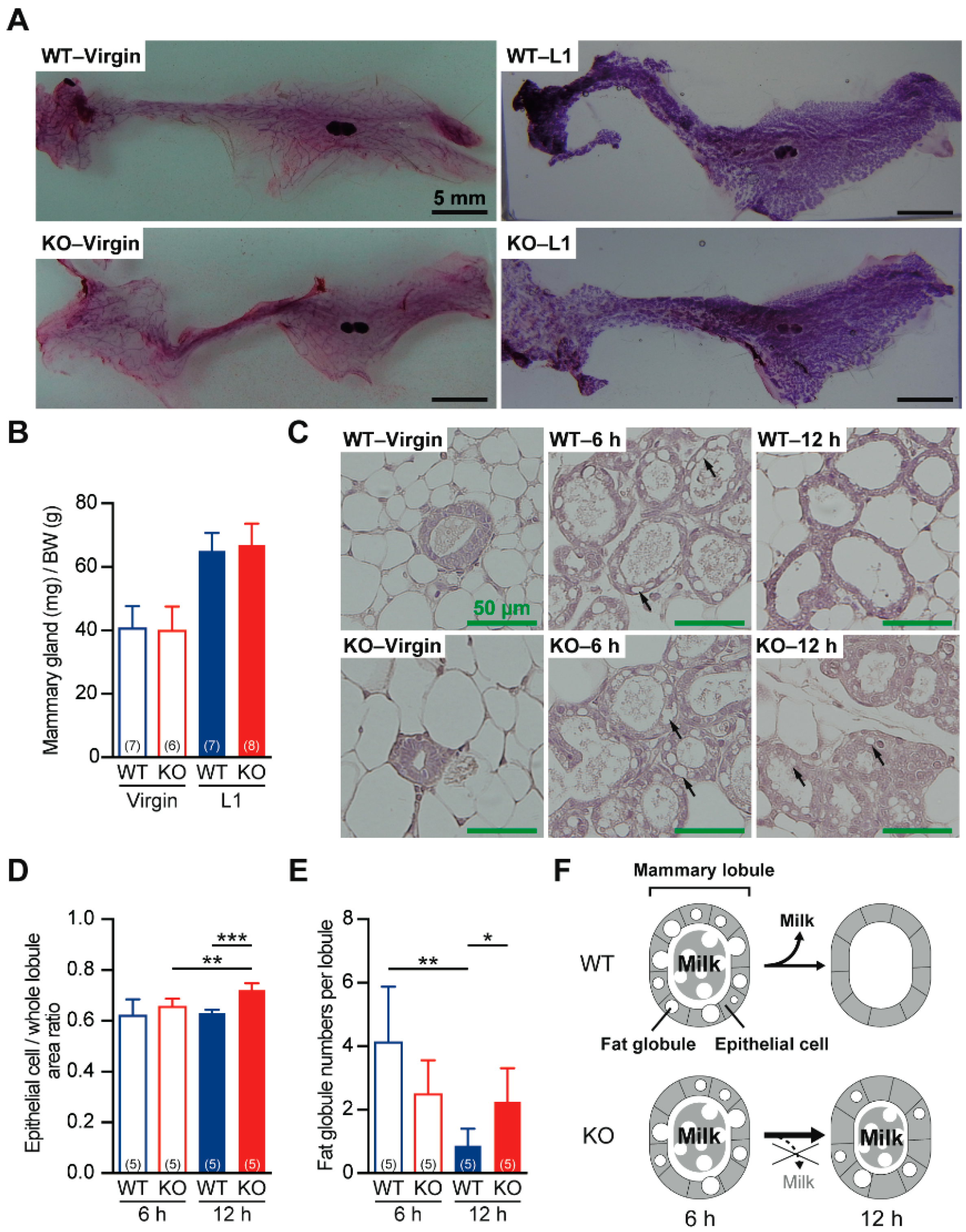
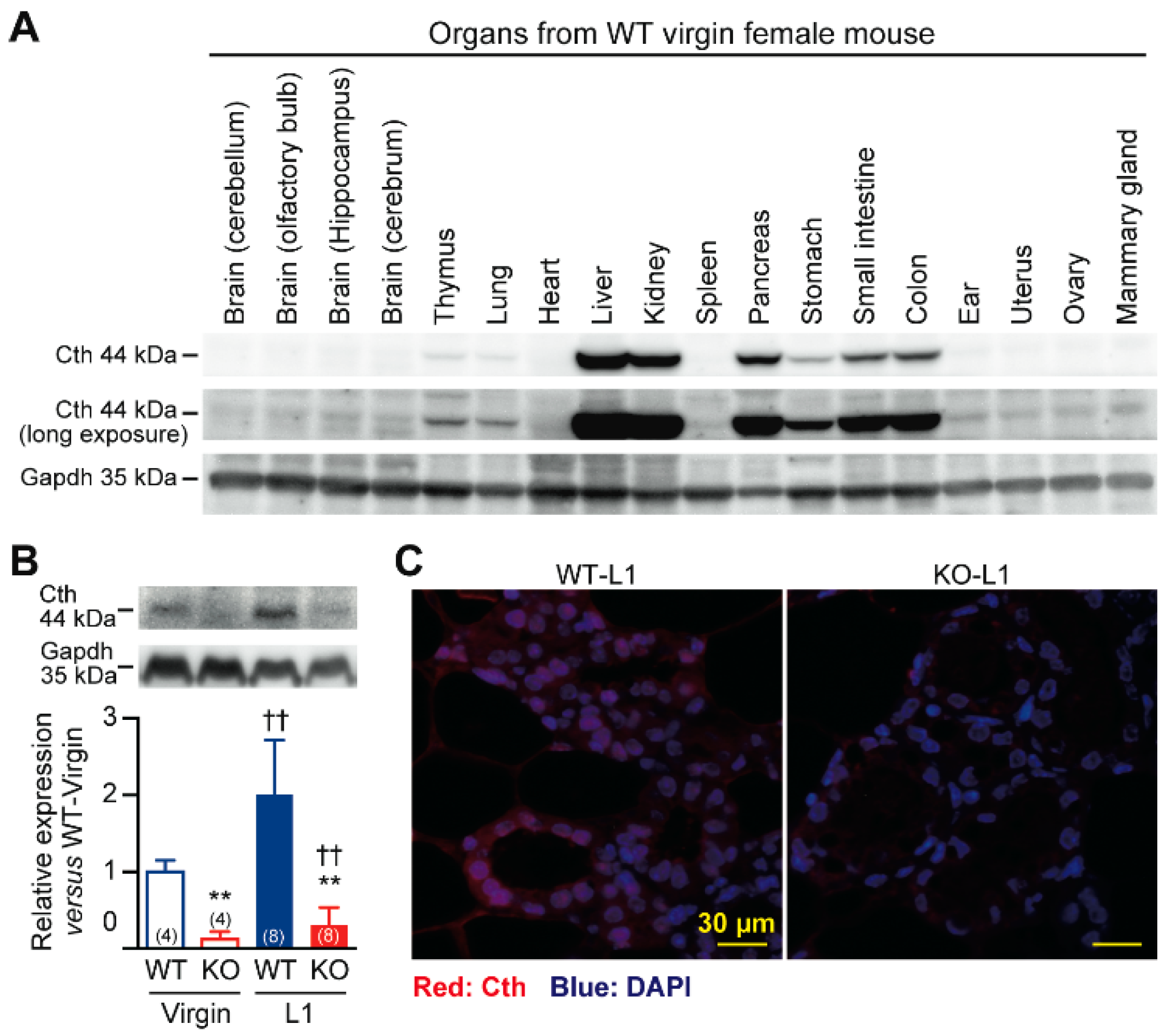
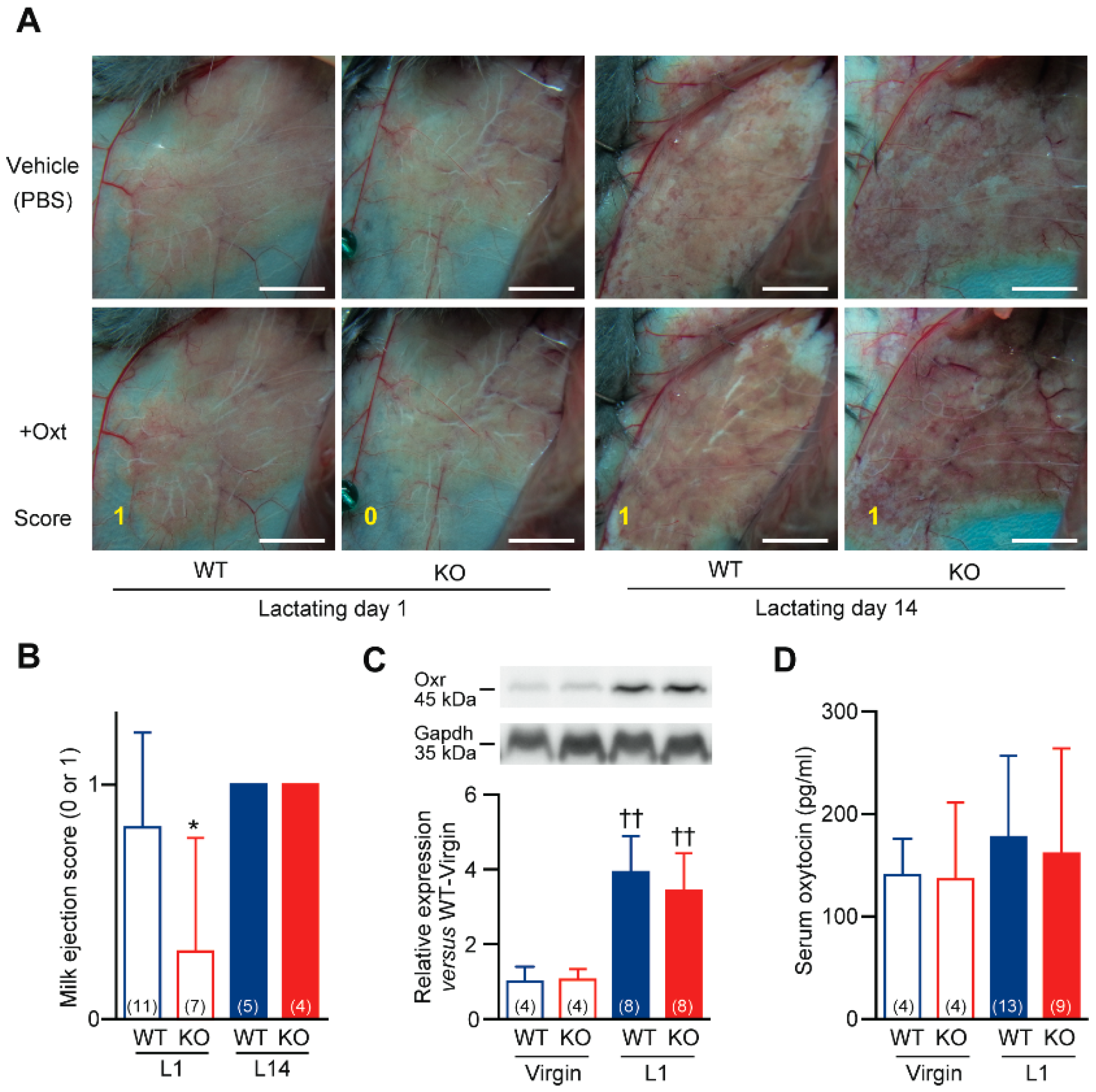
2.3. Defective Milk Ejection from Cth−/− Dams
2.4. Impaired Milk Ejection Responses to Oxytocin in Cth−/− Dams
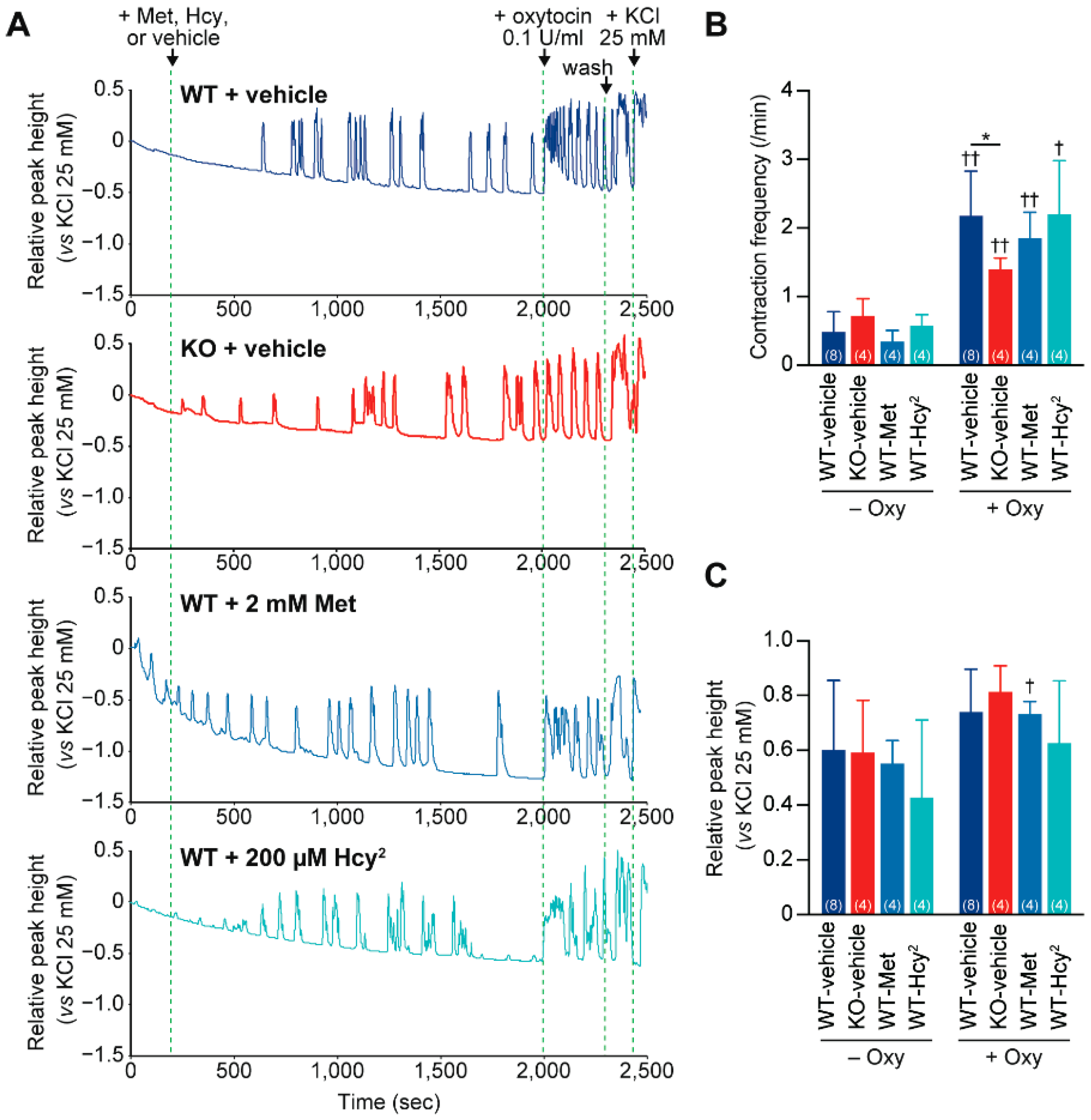
2.5. Impaired Contraction Responses to Oxytocin in Cth−/− Dam Uteruses
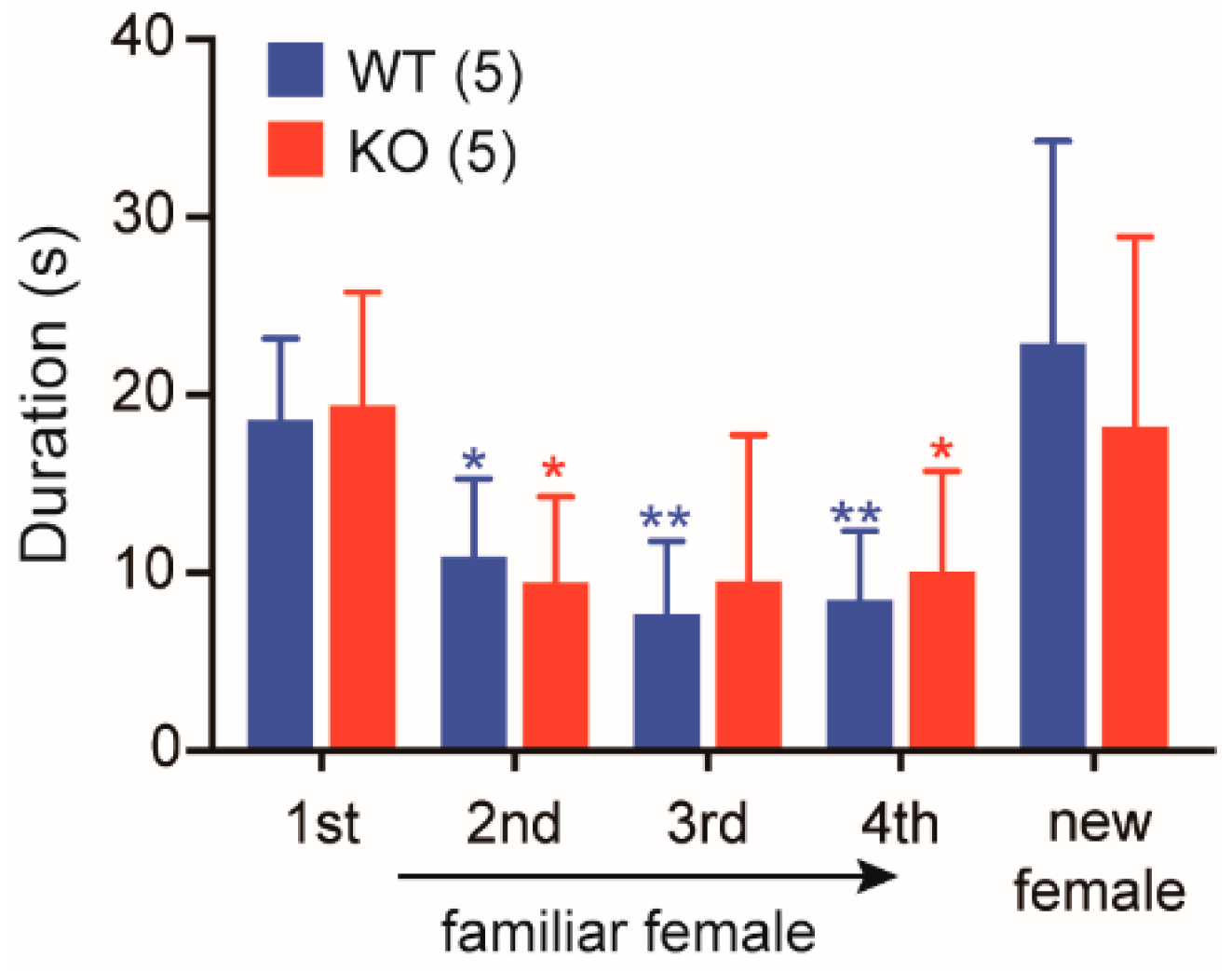
2.6. Normal Social Memory/Recognition in Cth−/− Male Mice
2.7. Cth−/− Dam Milk With Altered Amino Acid Profiles Did not Affect Serum Amino Acid Profiles in Their Pups
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Blood Pressure/Heart Rate Measurement
4.3. Measurement of the Biochemical Parameters in the Serum and Urine
4.4. Maternal Behavior Analyses
4.5. Histological Analyses of Mammary Glands
4.6. Western Blot Analyses
4.7. Milk Ejection Assay
4.8. Uterine Contraction Assay
4.9. Social Memory/Recognition Test
4.10. Amino acid Measurement of Serum and Whey Samples
4.11. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AEF | artificial extracellular fluid |
| Cbs | cystathionine β-synthase |
| CNS | central nervous system |
| Cth | cystathionine γ-lyase |
| CVD | cardiovascular disease |
| H2S | Hydrogen sulfide |
| NTD | neural tube defect |
| Oxtr | oxytocin receptor |
| PPG | propargylglycine |
| V2r | vasopressin receptor 2 |
| WT | wild-type |
References
- Herrmann, M.; Whiting, M.J.; Veillard, A.S.; Ehnholm, C.; Sullivan, D.R.; Keech, A.C. Plasma homocysteine and the risk of venous thromboembolism: Insights from the FIELD study. Clin. Chem. Lab. Med. 2012, 50, 2213–2219. [Google Scholar] [CrossRef]
- Ma, Y.; Peng, D.; Liu, C.; Huang, C.; Luo, J. Serum high concentrations of homocysteine and low levels of folic acid and vitamin B12 are significantly correlated with the categories of coronary artery diseases. BMC Cardiovasc. Disord. 2017, 17, 37. [Google Scholar] [CrossRef]
- Schroecksnadel, K.; Frick, B.; Fuchs, D. Homocysteine is an independent risk factor for myocardial infarction, in particular fatal myocardial infarction in middle-aged women. Circulation 2004, 110, e37–e38. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, Y.; Li, P.; Li, J.; Bao, H.; Zhang, Y.; Wang, B.; Sun, N.; Wang, J.; He, M.; et al. Association between percent decline in serum total homocysteine and risk of first stroke. Neurology 2017, 89, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Wald, D.S.; Law, M.; Morris, J.K. Homocysteine and cardiovascular disease: Evidence on causality from a meta-analysis. BMJ 2002, 325, 1202. [Google Scholar] [CrossRef] [PubMed]
- Mudd, S.H.; Levy, H.L.; Kraus, J.P. Disorders of transsulfuration. In The Metabolic and Molecular Basis of Inherited Disease, 8th ed.; Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D., Eds.; McGraw–Hill: New York, NY, USA, 2001; pp. 2007–2056. [Google Scholar]
- Dekker, G.A.; de Vries, J.I.; Doelitzsch, P.M.; Huijgens, P.C.; von Blomberg, B.M.; Jakobs, C.; van Geijn, H.P. Underlying disorders associated with severe early-onset preeclampsia. Am. J. Obstet. Gynecol. 1995, 173, 1042–1048. [Google Scholar] [CrossRef]
- Wouters, M.G.; Boers, G.H.; Blom, H.J.; Trijbels, F.J.; Thomas, C.M.; Borm, G.F.; Steegers-Theunissen, R.P.; Eskes, T.K. Hyperhomocysteinemia: A risk factor in women with unexplained recurrent early pregnancy loss. Fertil. Steril. 1993, 60, 820–825. [Google Scholar] [CrossRef]
- Burke, G.; Robinson, K.; Refsum, H.; Stuart, B.; Drumm, J.; Graham, I. Intrauterine growth retardation, perinatal death, and maternal homocysteine levels. N. Engl. J. Med. 1992, 326, 69–70. [Google Scholar] [PubMed]
- El-Khairy, L.; Vollset, S.E.; Refsum, H.; Ueland, P.M. Plasma total cysteine, pregnancy complications, and adverse pregnancy outcomes: The Hordaland homocysteine study. Am. J. Clin. Nutr. 2003, 77, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Kirke, P.N.; Mills, J.L.; Scott, J.M. Homocysteine metabolism in pregnancies complicated by neural tube defects. Nutrition 1997, 13, 994–995. [Google Scholar] [CrossRef]
- Trends in wheat-flour fortification with folic acid and iron—worldwide, 2004 and 2007. MMWR Morb. Mortal. Wkly Rep. 2008, 57, 8–10.
- Bell, K.N.; Oakley, G.P., Jr. Update on prevention of folic acid-preventable spina bifida and anencephaly. Birth Defects Res. A Clin. Mol. Teratol. 2009, 85, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Chrysant, S.G.; Chrysant, G.S. The current status of homocysteine as a risk factor for cardiovascular disease: A mini review. Expert Rev. Cardiovasc. Ther. 2018, 16, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Xu, C.H.; Xu, Y.N.; Wang, Y.L.; Wang, M. The effect of folate fortification on folic acid-based homocysteine-lowering intervention and stroke risk: A meta-analysis. Public Health Nutr. 2015, 18, 1514–1521. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.V.; Newcombe, P.; Hubacek, J.A.; Sofat, R.; Ricketts, S.L.; Cooper, J.; Breteler, M.M.; Bautista, L.E.; Sharma, P.; Whittaker, J.C.; et al. Effect modification by population dietary folate on the association between MTHFR genotype, homocysteine, and stroke risk: A meta-analysis of genetic studies and randomised trials. Lancet 2011, 378, 584–594. [Google Scholar] [CrossRef]
- Keller, R.; Chrastina, P.; Pavlikova, M.; Gouveia, S.; Ribes, A.; Kolker, S.; Blom, H.J.; Baumgartner, M.R.; Bartl, J.; Dionisi Vici, C.; et al. Newborn screening for homocystinurias: Recent recommendations versus current practice. J. Inherit. Metab. Dis. 2019, 42, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Osada, J.; Aratani, Y.; Kluckman, K.; Reddick, R.; Malinow, M.R.; Maeda, N. Mice deficient in cystathionine beta-synthase: Animal models for mild and severe homocyst(e)inemia. Proc. Natl. Acad. Sci. USA 1995, 92, 1585–1589. [Google Scholar] [CrossRef] [PubMed]
- Akahoshi, N.; Kobayashi, C.; Ishizaki, Y.; Izumi, T.; Himi, T.; Suematsu, M.; Ishii, I. Genetic background conversion ameliorates semi-lethality and permits behavioral analyses in cystathionine beta-synthase-deficient mice, an animal model for hyperhomocysteinemia. Hum. Mol. Genet. 2008, 17, 1994–2005. [Google Scholar] [CrossRef]
- Namekata, K.; Enokido, Y.; Ishii, I.; Nagai, Y.; Harada, T.; Kimura, H. Abnormal lipid metabolism in cystathionine beta-synthase-deficient mice, an animal model for hyperhomocysteinemia. J. Biol. Chem. 2004, 279, 52961–52969. [Google Scholar] [CrossRef]
- Ishii, I.; Akahoshi, N.; Yamada, H.; Nakano, S.; Izumi, T.; Suematsu, M. Cystathionine gamma-lyase-deficient mice require dietary cysteine to protect against acute lethal myopathy and oxidative injury. J. Biol. Chem. 2010, 285, 26358–26368. [Google Scholar] [CrossRef]
- Perry, T.L.; Hardwick, D.F.; Hansen, S.; Love, D.L.; Israels, S. Cystathioninuria in two healty siblings. N. Engl. J. Med. 1968, 278, 590–592. [Google Scholar] [CrossRef] [PubMed]
- Akahoshi, N.; Kamata, S.; Kubota, M.; Hishiki, T.; Nagahata, Y.; Matsuura, T.; Yamazaki, C.; Yoshida, Y.; Yamada, H.; Ishizaki, Y.; et al. Neutral aminoaciduria in cystathionine beta-synthase-deficient mice, an animal model of homocystinuria. Am. J. Physiol. Renal Physiol. 2014, 306, F1462–F1476. [Google Scholar] [CrossRef] [PubMed]
- Akahoshi, N.; Izumi, T.; Ishizaki, Y.; Ishii, I. Differential induction of cystathionine gamma-lyase in the livers and kidneys of mouse dams during gestation and lactation. Biol. Pharm. Bull. 2006, 29, 1799–1802. [Google Scholar] [CrossRef]
- Barber, T.; Triguero, A.; Martinez-Lopez, I.; Torres, L.; Garcia, C.; Miralles, V.J.; Vina, J.R. Elevated expression of liver gamma-cystathionase is required for the maintenance of lactation in rats. J. Nutr. 1999, 129, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza, R.; Garcia, C.; Rus, A.D.; Pallardo, F.V.; Barber, T.; Torres, L.; Miralles, V.J.; Vina, J.R. Inhibition of liver trans-sulphuration pathway by propargylglycine mimics gene expression changes found in the mammary gland of weaned lactating rats: Role of glutathione. Biochem. J. 2003, 373, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ahmad, S.; Cai, M.; Rennie, J.; Fujisawa, T.; Crispi, F.; Baily, J.; Miller, M.R.; Cudmore, M.; Hadoke, P.W.; et al. Dysregulation of hydrogen sulfide producing enzyme cystathionine gamma-lyase contributes to maternal hypertension and placental abnormalities in preeclampsia. Circulation 2013, 127, 2514–2522. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Ishii, I.; Shinmura, K.; Tamaki, K.; Hishiki, T.; Akahoshi, N.; Ida, T.; Nakanishi, T.; Kamata, S.; Kumagai, Y.; et al. Hyperhomocysteinemia abrogates fasting-induced cardioprotection against ischemia/reperfusion by limiting bioavailability of hydrogen sulfide anions. J. Mol. Med. 2015, 93, 879–889. [Google Scholar] [CrossRef]
- Bernard, V.; Young, J.; Binart, N. Prolactin—A pleiotropic factor in health and disease. Nat. Rev. Endocrinol. 2019, 15, 356–365. [Google Scholar] [CrossRef]
- Lincoln, D.W.; Wakerley, J.B. Factors governing the periodic activation of supraoptic and paraventricular neurosecretory cells during suckling in the rat. J. Physiol. 1975, 250, 443–461. [Google Scholar] [CrossRef]
- Higuchi, T.; Tadokoro, Y.; Honda, K.; Negoro, H. Detailed analysis of blood oxytocin levels during suckling and parturition in the rat. J. Endocrinol. 1986, 110, 251–256. [Google Scholar] [CrossRef]
- Gimpl, G.; Fahrenholz, F. The oxytocin receptor system: Structure, function, and regulation. Physiol. Rev. 2001, 81, 629–683. [Google Scholar] [CrossRef] [PubMed]
- Baxley, S.E.; Jiang, W.; Serra, R. Misexpression of wingless-related MMTV integration site 5A in mouse mammary gland inhibits the milk ejection response and regulates connexin43 phosphorylation. Biol. Reprod. 2011, 85, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Bankir, L.; Bichet, D.G.; Morgenthaler, N.G. Vasopressin: Physiology, assessment and osmosensation. J. Intern. Med. 2017, 282, 284–297. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Schoneberg, T.; Liu, J.; Schulz, A.; Ecelbarger, C.A.; Promeneur, D.; Nielsen, S.; Sheng, H.; Grinberg, A.; Deng, C.; et al. Generation and phenotype of mice harboring a nonsense mutation in the V2 vasopressin receptor gene. J. Clin. Investig. 2000, 106, 1361–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, J.N.; Aldag, J.M.; Insel, T.R.; Young, L.J. Oxytocin in the medial amygdala is essential for social recognition in the mouse. J. Neurosci. 2001, 21, 8278–8285. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Liu, H.X.; Hirai, H.; Torashima, T.; Nagai, T.; Lopatina, O.; Shnayder, N.A.; Yamada, K.; Noda, M.; Seike, T.; et al. CD38 is critical for social behaviour by regulating oxytocin secretion. Nature 2007, 446, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.N.; Young, L.J.; Hearn, E.F.; Matzuk, M.M.; Insel, T.R.; Winslow, J.T. Social amnesia in mice lacking the oxytocin gene. Nat. Genet. 2000, 25, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Ha, Y.S.; Soni, N.; An, G.I.; Lee, W.; Kim, M.H.; Huynh, P.T.; Ahn, H.; Bhatt, N.; Lee, Y.J.; et al. Immobilization of the gas signaling molecule H2S by radioisotopes: Detection, quantification, and in vivo imaging. Angew. Chem. Int. Ed. Engl. 2016, 55, 9365–9370. [Google Scholar] [CrossRef]
- Han, S.J.; Noh, M.R.; Jung, J.M.; Ishii, I.; Yoo, J.; Kim, J.I.; Park, K.M. Hydrogen sulfide-producing cystathionine gamma-lyase is critical in the progression of kidney fibrosis. Free Radic. Biol. Med. 2017, 112, 423–432. [Google Scholar] [CrossRef]
- Szijarto, I.A.; Marko, L.; Filipovic, M.R.; Miljkovic, J.L.; Tabeling, C.; Tsvetkov, D.; Wang, N.; Rabelo, L.A.; Witzenrath, M.; Diedrich, A.; et al. Cystathionine gamma-lyase-produced hydrogen sulfide controls endothelial NO bioavailability and blood pressure. Hypertension 2018, 71, 1210–1217. [Google Scholar] [CrossRef]
- Yamada, H.; Akahoshi, N.; Kamata, S.; Hagiya, Y.; Hishiki, T.; Nagahata, Y.; Matsuura, T.; Takano, N.; Mori, M.; Ishizaki, Y.; et al. Methionine excess in diet induces acute lethal hepatitis in mice lacking cystathionine gamma-lyase, an animal model of cystathioninuria. Free Radic. Biol. Med. 2012, 52, 1716–1726. [Google Scholar] [CrossRef] [PubMed]
- Hagiya, Y.; Kamata, S.; Mitsuoka, S.; Okada, N.; Yoshida, S.; Yamamoto, J.; Ohkubo, R.; Abiko, Y.; Yamada, H.; Akahoshi, N.; et al. Hemizygosity of transsulfuration genes confers increased vulnerability against acetaminophen-induced hepatotoxicity in mice. Toxicol. Appl. Pharmacol. 2015, 282, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Ishii, I.; Kamata, S.; Hagiya, Y.; Abiko, Y.; Kasahara, T.; Kumagai, Y. Protective effects of hydrogen sulfide anions against acetaminophen-induced hepatotoxicity in mice. J. Toxicol. Sci. 2015, 40, 837–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiyama, M.; Shinkai, Y.; Unoki, T.; Shim, I.; Ishii, I.; Kumagai, Y. The capture of cadmium by reactive polysulfides attenuates cadmium-induced adaptive responses and hepatotoxicity. Chem. Res. Toxicol. 2017, 30, 2209–2217. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.; Unoki, T.; Shinkai, Y.; Ishii, I.; Ida, T.; Akaike, T.; Yamamoto, M.; Kumagai, Y. Environmental electrophile-mediated toxicity in mice lacking Nrf2, CSE, or both. Environ. Health Perspect. 2019, 6, 67002. [Google Scholar] [CrossRef] [PubMed]
- Abiko, Y.; Yoshida, E.; Ishii, I.; Fukuto, J.M.; Akaike, T.; Kumagai, Y. Involvement of reactive persulfides in biological bismethylmercury sulfide formation. Chem. Res. Toxicol. 2015, 28, 1301–1306. [Google Scholar] [CrossRef]
- Vest, A.R.; Cho, L.S. Hypertension in pregnancy. Curr. Atheroscler. Rep. 2014, 16, 395. [Google Scholar] [CrossRef]
- Kuma, A.; Hatano, M.; Matsui, M.; Yamamoto, A.; Nakaya, H.; Yoshimori, T.; Ohsumi, Y.; Tokuhisa, T.; Mizushima, N. The role of autophagy during the early neonatal starvation period. Nature 2004, 432, 1032–1036. [Google Scholar] [CrossRef]
- Sonne, S.R.; Bhalla, V.K.; Barman, S.A.; White, R.E.; Zhu, S.; Newman, T.M.; Prasad, P.D.; Smith, S.B.; Offermanns, S.; Ganapathy, V. Hyperhomocysteinemia is detrimental to pregnancy in mice and is associated with preterm birth. Biochim. Biophys. Acta 2013, 1832, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Bridges, R.S. Neuroendocrine regulation of maternal behavior. Front. Neuroendocrinol. 2015, 36, 178–196. [Google Scholar] [CrossRef]
- Argiolas, A.; Gessa, G.L. Central functions of oxytocin. Neurosci. Biobehav. Rev. 1991, 15, 217–231. [Google Scholar] [CrossRef]
- Caldwell, H.K.; Aulino, E.A.; Freeman, A.R.; Miller, T.V.; Witchey, S.K. Oxytocin and behavior: Lessons from knockout mice. Dev. Neurobiol. 2017, 77, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Nishimori, K.; Young, L.J.; Guo, Q.; Wang, Z.; Insel, T.R.; Matzuk, M.M. Oxytocin is required for nursing but is not essential for parturition or reproductive behavior. Proc. Natl. Acad. Sci. USA 1996, 93, 11699–11704. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, Y.; Yoshida, M.; Bielsky, I.F.; Ross, H.E.; Kawamata, M.; Onaka, T.; Yanagisawa, T.; Kimura, T.; Matzuk, M.M.; Young, L.J.; et al. Pervasive social deficits, but normal parturition, in oxytocin receptor-deficient mice. Proc. Natl. Acad. Sci. USA 2005, 102, 16096–16101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudd, S.H.; Finkelstein, J.D.; Refsum, H.; Ueland, P.M.; Malinow, M.R.; Lentz, S.R.; Jacobsen, D.W.; Brattstrom, L.; Wilcken, B.; Wilcken, D.E.; et al. Homocysteine and its disulfide derivatives: A suggested consensus terminology. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1704–1706. [Google Scholar] [CrossRef] [PubMed]
- Akahoshi, N.; Yokoyama, A.; Nagata, T.; Miura, A.; Kamata, S.; Ishii, I. Abnormal amino acid profiles of blood and cerebrospinal fluid from cystathionine β-synthase-deficient mice, an animal model of homocystinuria. Biol. Pharm. Bull. 2019, 42, 1054–1057. [Google Scholar] [CrossRef]
- Dapson, R.W. The history, chemistry and modes of action of carmine and related dyes. Biotech. Histochem. 2007, 82, 173–187. [Google Scholar] [CrossRef]
- Ishii, I.; Akahoshi, N.; Yu, X.N.; Kobayashi, Y.; Namekata, K.; Komaki, G.; Kimura, H. Murine cystathionine gamma-lyase: Complete cDNA and genomic sequences, promoter activity, tissue distribution and developmental expression. Biochem. J. 2004, 381, 113–123. [Google Scholar] [CrossRef]
- Plante, I.; Stewart, M.K.; Laird, D.W. Evaluation of mammary gland development and function in mouse models. J. Vis. Exp. 2011, 53, e2828. [Google Scholar] [CrossRef]
- Yamamoto, J.; Kamata, S.; Miura, A.; Nagata, T.; Kainuma, R.; Ishii, I. Differential adaptive responses to 1- or 2-day fasting in various mouse tissues revealed by quantitative PCR analysis. FEBS Open Bio. 2015, 5, 357–368. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum from Virgin | Serum from Primipara | Whey from Primiparas | ||||
|---|---|---|---|---|---|---|
| WT (n = 9) | Cth−/− (n = 7) | WT (n = 7) | Cth−/− (n = 12) | WT (n = 7) | Cth−/− (n = 12) | |
| Ala | 446 ± 78 | 471 ± 115 | 618 ± 395 | 1164 ± 799 # | 309 ± 79 | 540 ± 121 *** |
| Arg | 133 ± 27 | 196 ± 39 ** | 46.0 ± 40.3 ## | 25.8 ± 27.1 ## | 35.3 ± 19.3 | 31.8 ± 9.7 |
| Asp/Asn | 12.4 ± 5.7 | 15.0 ± 9.9 | 16.6 ± 5.0 | 15.2 ± 6.9 | 40.8 ± 15.8 | 24.5 ± 8.0 * |
| Gln | 837 ± 125 | 846 ± 144 | 873 ± 318 | 1344 ± 482 *,## | 74.4 ± 21.1 | 181 ± 46 *** |
| Glu | 20.7 ± 11.8 | 18.8 ± 11.6 | 58.5 ± 30.2 # | 36.2 ± 19.2 # | 211 ± 48 | 190 ± 43 |
| Gly | 403 ± 60 | 424 ± 71 | 359 ± 153 | 520 ± 185 | 271 ± 60 | 414 ± 77 *** |
| His | 41.6 ± 7.4 | 85.3 ± 11.5 *** | 51.9 ± 25.4 | 195 ± 67 ***,## | 7.07 ± 2.52 | 24.0 ± 5.0 *** |
| Ile | 121 ± 16 | 110 ± 33 | 113 ± 44 | 169 ± 60 *,# | 29.7 ± 19.4 | 35.0 ± 14.8 |
| Leu | 156 ± 13 | 153 ± 45 | 158 ± 77 | 274 ± 113 *,## | 24.6 ± 8.9 | 34.9 ± 13.6 |
| Lys | 277 ± 50 | 287 ± 87 | 290 ± 89 | 663 ± 292 **,## | 70.0 ± 19.2 | 113 ± 37 ** |
| Met | 72.0 ± 8.8 | 94.2 ± 10.8 *** | 128 ± 51# | 147 ± 44 ## | 5.49 ± 1.22 | 10.1 ± 2.7 *** |
| Phe | 96.5 ± 17.6 | 89.1 ± 15.9 | 93.2 ± 53.3 | 92.4 ± 39.9 | 1.25 ± 1.34 | 2.60 ± 1.25 |
| Pro | 121 ± 20 | 140 ± 50 | 298 ± 237 | 288 ± 187 # | 312 ± 177 | 237 ± 71 |
| Ser | 166 ± 34 | 162 ± 37 | 189 ± 98 | 239 ± 96 # | 234 ± 66 | 306 ± 84 |
| Thr | 185 ± 28 | 190 ± 60 | 248 ± 108 | 368 ± 148 ## | 111 ± 38 | 158 ± 40 * |
| Tyr | 156 ± 29 | 148 ± 45 | 86.8 ± 61.5 # | 145 ± 86 | N.D. | N.D. |
| Val | 258 ± 21 | 239 ± 80 | 234 ± 87 | 386 ± 165 *,# | 76.0 ± 43.5 | 105 ± 59 |
| Cystathionine | 18.5 ± 3.0 | 127 ± 34 *** | 54.7 ± 44.0 | 606 ± 298 ***,## | 36.9 ± 14.8 | 361 ± 95 *** |
| Citrulline | 85.3 ± 8.9 | 215 ± 57 *** | 65.8 ± 14.4 # | 242 ± 77 *** | 6.76 ± 0.88 | 32.3 ± 8.0 *** |
| Ornithine | 68.1 ± 9.9 | 87.1 ± 25.1 | 92.2 ± 47.1 | 220 ± 131 **,## | 21.5 ± 4.0 | 58.9 ± 14.4 *** |
| Taurine | 385 ± 247 | 487 ± 226 | 835 ± 256 ## | 725 ± 422 | 384 ± 27 | 396 ± 89 |
| Total Cys | 289 ± 41 | 197 ± 18 *** | 508 ± 82 ## | 384 ± 85 **,## | 154 ± 80 | 164 ± 35 |
| Total Hcy | 10.8 ± 5.8 | 181 ± 16 *** | 11.3 ± 1.8 | 111 ± 24 ***,## | 6.09 ± 11.33 | 49.6 ± 13.4 *** |
| Total GSH | 97.5 ± 52.1 | 68.4 ± 16.6 | 176 ± 76 # | 136 ± 120 | 23.6 ± 3.5 | 22.3 ± 3.7 |
| Total Cys-gly | 3.71 ± 0.7 | N.D. | 5.93 ± 1.95 # | N.D. | 41.7 ± 8.4 | 43.2 ± 14.8 |
| Total γ-Glu-Cys | 11.2 ± 1.6 | 7.32 ± 0.31 ** | 21.1 ± 4.9 ## | 12.2 ± 3.3 **,## | 27.3 ± 48.1 | 43.3 ± 19.0 |
| Cth+/− Pups at 6 h after Birth | Cth+/− Pups at 12 h after Birth | |||
|---|---|---|---|---|
| From Wild-Type | From Cth−/− | From Wild-Type | From Cth−/− | |
| (n = 3 Litters) | (n = 5 Litters) | (n = 4 Litters) | (n = 4 Litters) | |
| Ala | 449 ± 392 | 219 ± 34 | 163 ± 23 | 168 ± 27 # |
| Arg | 6.72 ± 0.13 | 18.0 ± 13.1 | 33.0 ± 18.2 | 26.8 ± 9.8 |
| Asp/Asn | 115 ± 44 | 104 ± 22 | 96.4 ± 25.4 | 122 ± 22 |
| Gln | 1801 ± 888 | 1666 ± 284 | 1585 ± 408 | 1661 ± 285 |
| Glu | 104 ± 28 | 92.2 ± 28.8 | 86.0 ± 10.2 | 94.6 ± 2 |
| Gly | 240 ± 46 | 176 ± 32 | 209 ± 24 | 216 ± 36 |
| His | 20.6 ± 2.1 | 20.2 ± 7.6 | 15.2 ± 0.7 # | 17.3 ± 4.3 |
| Ile | 83.2 ± 35.8 | 70.0 ± 14.5 | 79.3 ± 5.6 | 82.2 ± 23 |
| Leu | 137 ± 60 | 110 ± 26 | 110 ± 5 | 114 ± 36 |
| Lys | 752 ± 194 | 625 ± 94 | 580 ± 47 | 668 ± 100 |
| Met | 118 ± 23 | 136 ± 26 | 83.6 ± 3.2 | 96.3 ± 16.3 # |
| Phe | 118 ± 34 | 117 ± 21 | 112 ± 6 | 120 ± 16 |
| Pro | 188 ± 107 | 155 ± 42 | 174 ± 9 | 171 ± 35 |
| Ser | 57.3 ± 11.6 | 57.2 ± 9.1 | 71.4 ± 7.8 | 62.6 ± 12.3 |
| Thr | 197 ± 102 | 170 ± 34 | 122 ± 9 | 127 ± 23 |
| Tyr | 140 ± 57 | 138 ± 31 | 105 ± 10 | 120 ± 22 |
| Val | 218 ± 64 | 180 ± 21 | 148 ± 7 | 160 ± 37 |
| Cystathionine | 97.5 ± 15.7 | 110 ± 21 | 80.8 ± 16.8 | 100 ± 10 |
| Citrulline | 56.6 ± 7 | 66.7 ± 10.9 | 64.1 ± 3.2 | 68.0 ± 6.7 |
| Ornithine | 131 ± 42 | 121 ± 15 | 96.9 ± 22.5 | 113 ± 35 |
| Taurine | 1874 ± 489 | 1334 ± 172 | 1292 ± 204 | 1455 ± 201 |
| Total Cys | 271 ± 15 | 281 ± 37 | 259 ± 16 | 279 ± 34 |
| Total Hcy | 4.93 ± 0.50 | 10.7 ± 3.7 * | 3.98 ± 0.92 | 6.41 ± 2.63 |
| Total GSH | 105 ± 46 | 68.2 ± 26.0 | 67.7 ± 13.7 | 69.2 ± 12.3 |
| Total γ-Glu-Cys | 20.0 ± 6.6 | 23.4 ± 2.1 | 24.9 ± 4.6 | 23.6 ± 3.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akahoshi, N.; Handa, H.; Takemoto, R.; Kamata, S.; Yoshida, M.; Onaka, T.; Ishii, I. Preeclampsia-Like Features and Partial Lactation Failure in Mice Lacking Cystathionine γ-Lyase—An Animal Model of Cystathioninuria. Int. J. Mol. Sci. 2019, 20, 3507. https://doi.org/10.3390/ijms20143507
Akahoshi N, Handa H, Takemoto R, Kamata S, Yoshida M, Onaka T, Ishii I. Preeclampsia-Like Features and Partial Lactation Failure in Mice Lacking Cystathionine γ-Lyase—An Animal Model of Cystathioninuria. International Journal of Molecular Sciences. 2019; 20(14):3507. https://doi.org/10.3390/ijms20143507
Chicago/Turabian StyleAkahoshi, Noriyuki, Hiroki Handa, Rintaro Takemoto, Shotaro Kamata, Masahide Yoshida, Tatsushi Onaka, and Isao Ishii. 2019. "Preeclampsia-Like Features and Partial Lactation Failure in Mice Lacking Cystathionine γ-Lyase—An Animal Model of Cystathioninuria" International Journal of Molecular Sciences 20, no. 14: 3507. https://doi.org/10.3390/ijms20143507