Effects of Diazepam on Low-Frequency and High-Frequency Electrocortical γ-Power Mediated by α1- and α2-GABAA Receptors


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
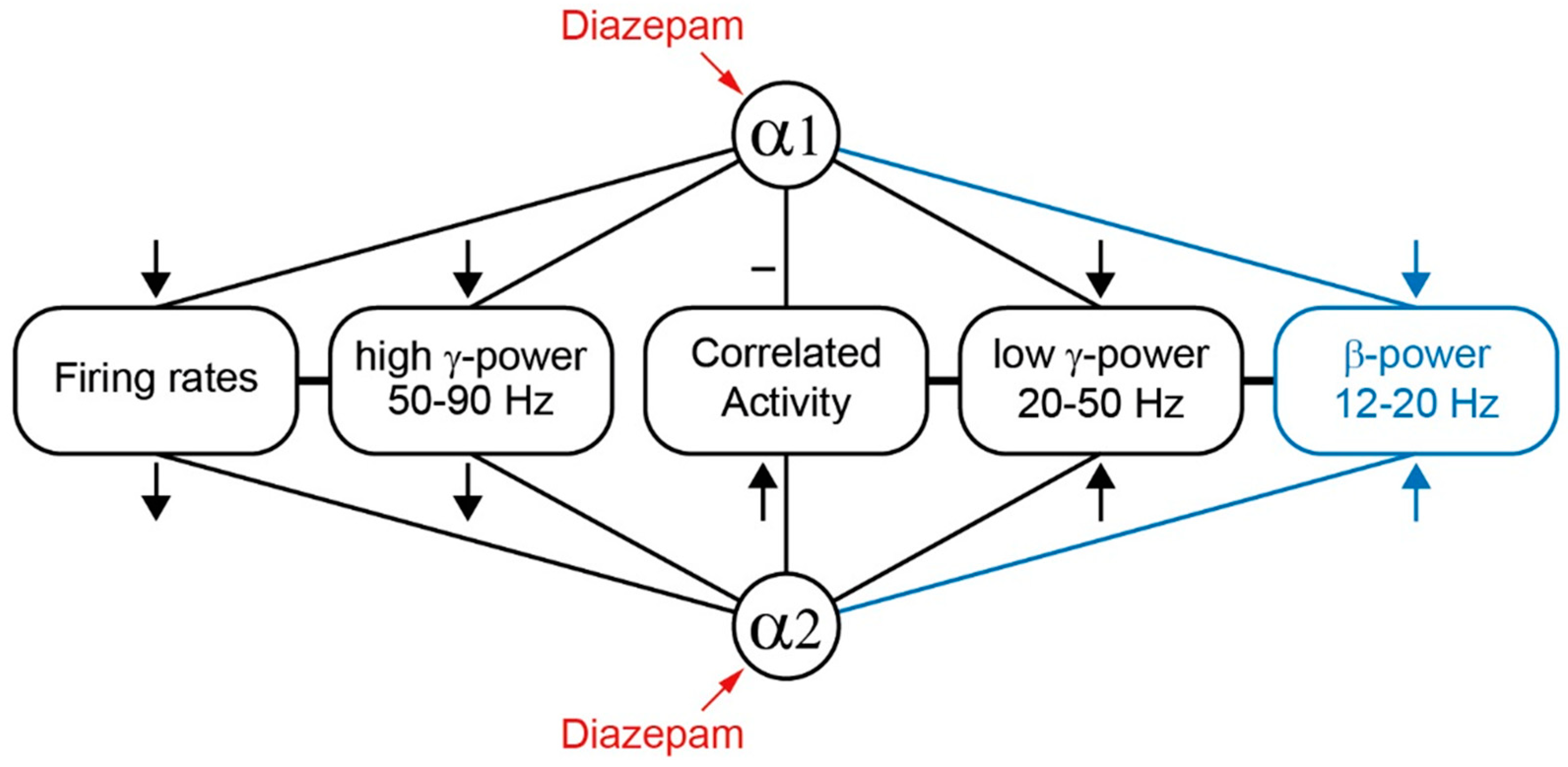
Abstract
:1. Introduction
2. Results
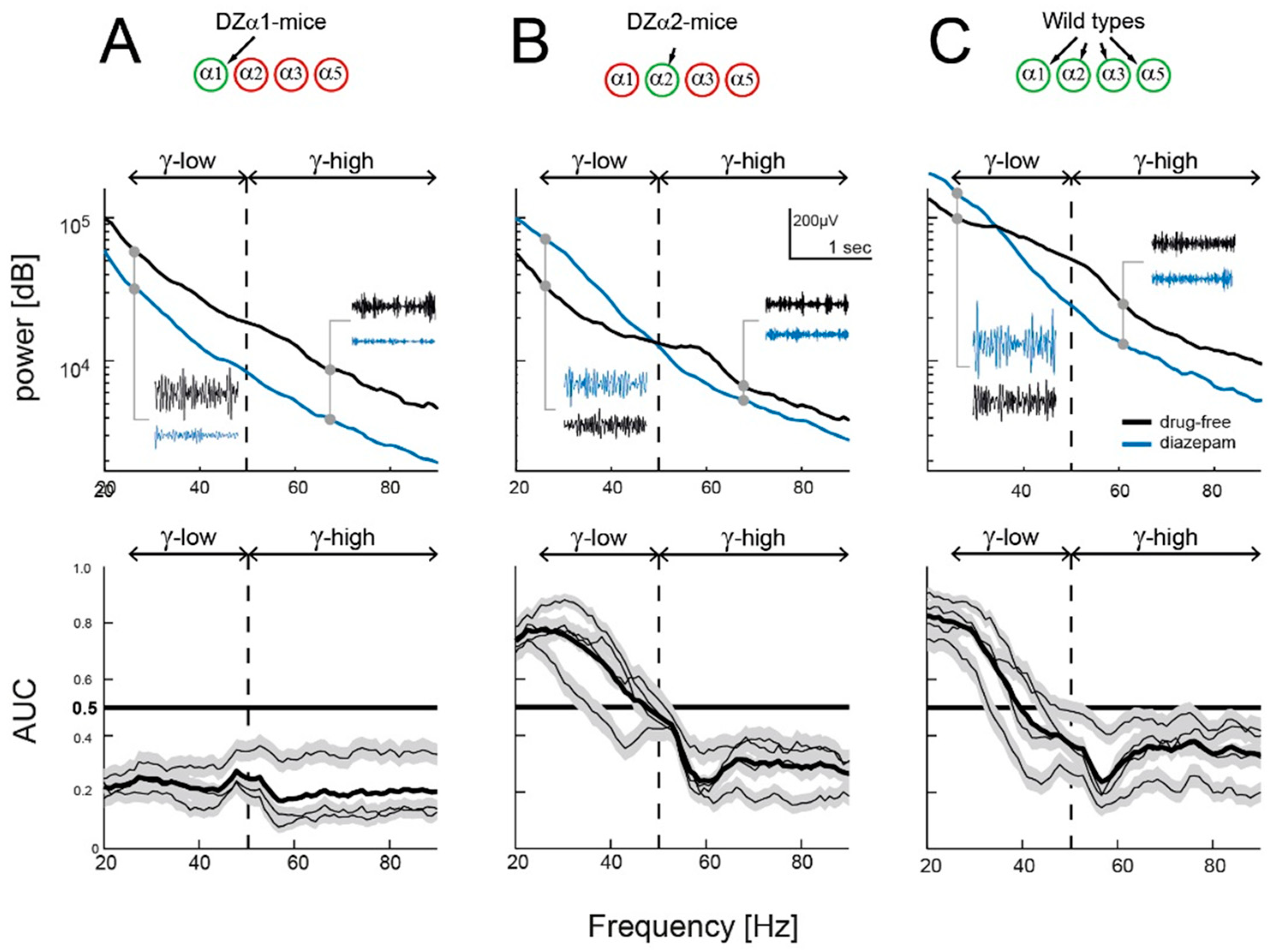
2.1. Diazepam’s Effects on Low- and High γ-Power
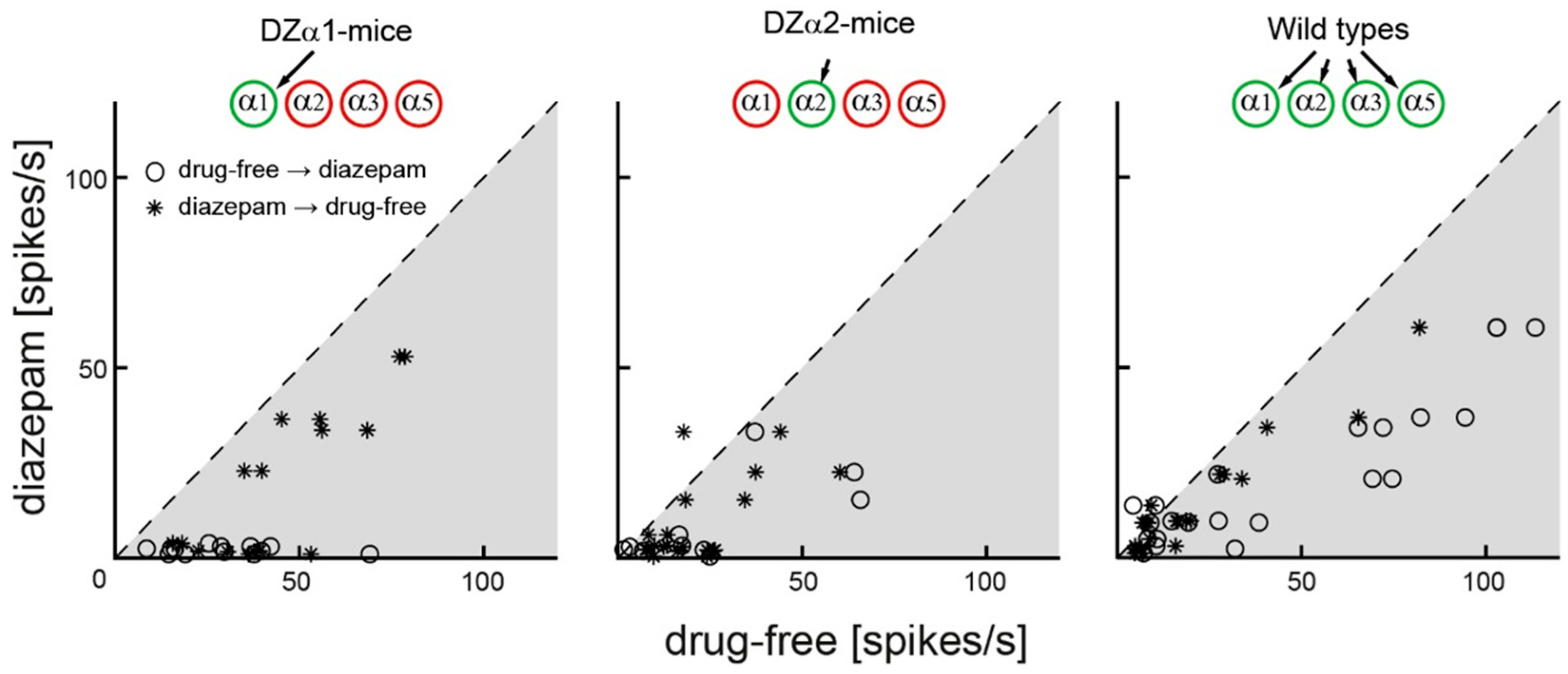
2.2. Diazepam Curtails Action Potential Firing in All Genotypes
2.3. Diazepam Increases Correlated Firing in the Low γ-Range via α2- but Not α1-GABAA Receptors
3. Discussion
3.1. Modulation of Low γ-Power via α2-GABAA Receptors
3.2. Diazepam Curtails High γ-Power and Action Potential Firing in All Genotypes
3.3. Related Studies on Single Knockin Mice
3.4. Diazepam-Induced Changes in Motor Activity
3.5. Limitations of the Present Study
4. Materials and Methods
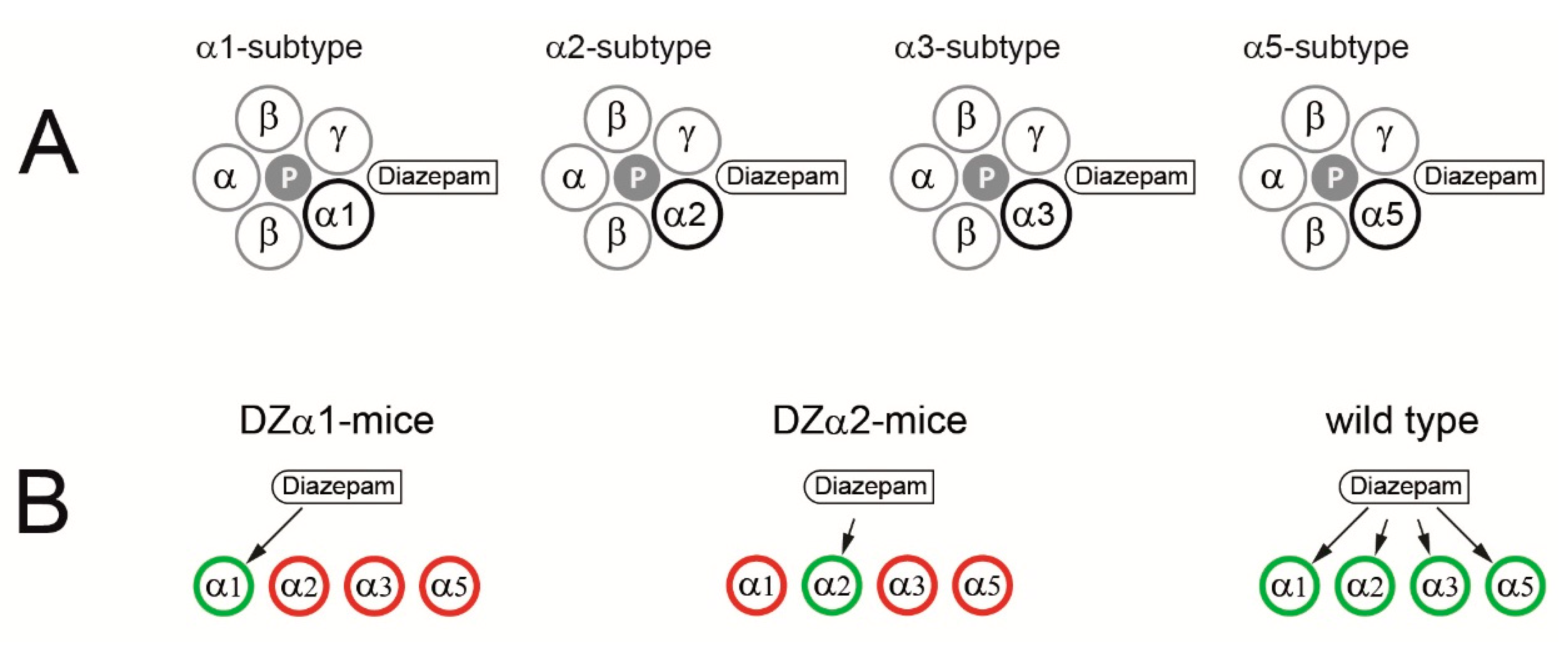
4.1. Animals
4.2. Implantation of Recording Electrodes
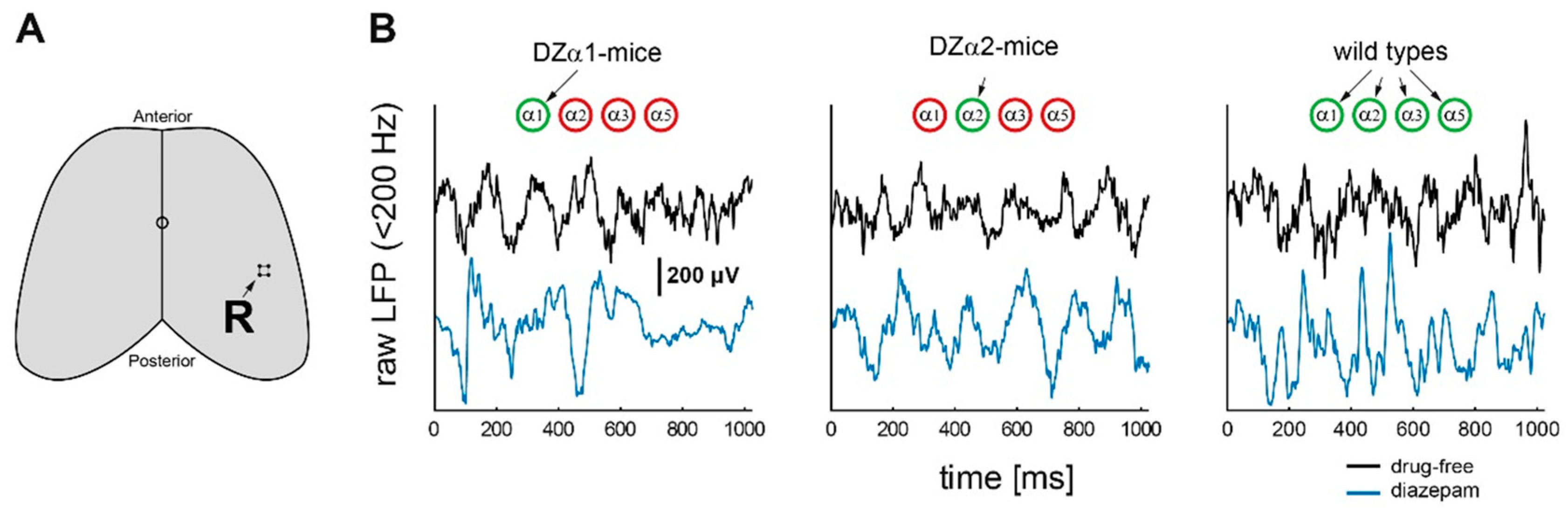
4.3. Drug Administration and Electrophysiological Measurements
4.4. Data Structure
LFP Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AUC | Area under the receiver-operated curve |
| EEG | Electroencephalogram |
| GABA | γ-aminobutyric acid |
| HCN1 | Hyperpolarization-activated cyclic nucleotide-gated channel 1 |
| LFP | Local field potential |
| NMDA | N-methyl-D-aspartate |
| ROC | Receiver operating characteristics |
References
- Rudolph, U.; Knoflach, F. Beyond classical benzodiazepines: Novel therapeutic potential of GABAA receptor subtypes. Nat. Rev. Drug Discov. 2011, 10, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Fritschy, J.M.; Panzanelli, P. GABAA receptors and plasticity of inhibitory neurotransmission in the central nervous system. Eur. J. Neurosci. 2014, 39, 1845–1865. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, U.; Mohler, H. GABAA receptor subtypes: Therapeutic potential in Down syndrome, affective disorders, schizophrenia, and autism. Annu. Rev. Pharmacol. Toxicol 2014, 54, 483–507. [Google Scholar] [CrossRef] [PubMed]
- Benson, J.A.; Low, K.; Keist, R.; Mohler, H.; Rudolph, U. Pharmacology of recombinant gamma-aminobutyric acida receptors rendered diazepam-insensitive by point-mutated alpha-subunits. FEBS Lett. 1998, 431, 400–404. [Google Scholar] [CrossRef]
- Prenosil, G.A.; Schneider Gasser, E.M.; Rudolph, U.; Keist, R.; Fritschy, J.M.; Vogt, K.E. Specific subtypes of GABAA receptors mediate phasic and tonic forms of inhibition in hippocampal pyramidal neurons. J. Neurophysiol. 2006, 96, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Ralvenius, W.T.; Benke, D.; Acuna, M.A.; Rudolph, U.; Zeilhofer, H.U. Analgesia and unwanted benzodiazepine effects in point-mutated mice expressing only one benzodiazepine-sensitive GABAA receptor subtype. Nat. Commun. 2015, 6, 6803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralvenius, W.T.; Neumann, E.; Pagani, M.; Acuna, M.A.; Wildner, H.; Benke, D.; Fischer, N.; Rostaher, A.; Schwager, S.; Detmar, M.; et al. Itch suppression in mice and dogs by modulation of spinal alpha2 and alpha3GABAA receptors. Nat. Commun. 2018, 9, 3230. [Google Scholar] [CrossRef]
- Sigel, E.; Steinmann, M.E. Structure, function, and modulation of GABA(A) receptors. J. Biol. Chem. 2012, 287, 40224–40231. [Google Scholar] [CrossRef]
- Masiulis, S.; Desai, R.; Uchanski, T.; Martin, I.S.; Laverty, D.; Karia, D.; Malinauskas, T.; Zivanov, J.; Pardon, E.; Kotecha, A.; et al. GABA(A) receptor signalling mechanisms revealed by structural pharmacology. Nature 2019, 566, 454–459. [Google Scholar] [CrossRef]
- Rudolph, U.; Crestani, F.; Benke, D.; Brunig, I.; Benson, J.A.; Fritschy, J.M.; Martin, J.R.; Bluethmann, H.; Mohler, H. Benzodiazepine actions mediated by specific gamma-aminobutyric acid(A) receptor subtypes. Nature 1999, 401, 796–800. [Google Scholar] [CrossRef]
- Löw, K.; Crestani, F.; Keist, R.; Benke, D.; Brünig, I.; Benson, J.A.; Fritschy, J.-M.; Rülicke, T.; Bluethmann, H.; Möhler, H.; et al. Molecular and neuronal substrate for the selectice attenuation of anxiety. Science 2000, 290, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Crestani, F.; Keist, R.; Fritschy, J.M.; Benke, D.; Vogt, K.; Prut, L.; Bluthmann, H.; Mohler, H.; Rudolph, U. Trace fear conditioning involves hippocampal alpha5 GABA(A) receptors. Proc. Natl. Acad. Sci. USA 2002, 99, 8980–8985. [Google Scholar] [CrossRef] [PubMed]
- Tobler, I.; Kopp, C.; Deboer, T.; Rudolph, U. Diazepam-induced changes in sleep: Role of the alpha 1 GABA(A) receptor subtype. Proc. Natl. Acad. Sci. USA 2001, 98, 6464–6469. [Google Scholar] [CrossRef] [PubMed]
- Christian, E.P.; Snyder, D.H.; Song, W.; Gurley, D.A.; Smolka, J.; Maier, D.L.; Ding, M.; Gharahdaghi, F.; Liu, X.F.; Chopra, M.; et al. EEG-beta/gamma spectral power elevation in rat: A translatable biomarker elicited by GABA(Aalpha2/3)-positive allosteric modulators at nonsedating anxiolytic doses. J. Neurophysiol. 2015, 113, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Edenberg, H.J.; Dick, D.M.; Xuei, X.L.; Tian, H.J.; Almasy, L.; Bauer, L.O.; Crowe, R.R.; Goate, A.; Hesselbrock, V.; Jones, K.; et al. Variations in GABRA2, encoding the alpha 2 subunit of the GABA(A) receptor, are associated with alcohol dependence and with brain oscillations. Am. J. Hum. Genet. 2004, 74, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.J.; Plourde, G. Attenuation of High-Frequency (50–200 Hz) Thalamocortical EEG Rhythms by Propofol in Rats Is More Pronounced for the Thalamus than for the Cortex. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Hambrecht-Wiedbusch, V.S.; Silverstein, B.H.; Mashour, G.A. Electroencephalographic coherence and cortical acetylcholine during ketamine-induced unconsciousness. Br. J. Anaesth. 2015, 114, 979–989. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Shu, S.; Bayliss, D.A. HCN1 channel subunits are a molecular substrate for hypnotic actions of ketamine. J. Neurosci. 2009, 29, 600–609. [Google Scholar] [CrossRef]
- Hudetz, A.G.; Vizuete, J.A.; Pillay, S. Differential Effects of Isoflurane on High-frequency and Low-frequency gamma Oscillations in the Cerebral Cortex and Hippocampus in Freely Moving Rats. Anesthesiology 2011, 114, 588–595. [Google Scholar] [CrossRef]
- Hentschke, H.; Schwarz, C.; Antkowiak, B. Neocortex is the major target of sedative concentrations of volatile anaesthetics: Strong depression of firing rates and increase of GABAA receptor-mediated inhibition. Eur. J. Neurosci. 2005, 21, 93–102. [Google Scholar] [CrossRef]
- Kopp, C.; Rudolph, U.; Low, K.; Tobler, I. Modulation of rhythmic brain activity by diazepam: GABA(A) receptor subtype and state specificity. Proc. Natl. Acad. Sci. USA 2004, 101, 3674–3679. [Google Scholar] [CrossRef] [PubMed]
- Kopp, C.; Rudolph, U.; Keist, R.; Tobler, I. Diazepam-induced changes on sleep and the EEG spectrum in mice: Role of the alpha3-GABA(A) receptor subtype. Eur. J. Neurosci 2003, 17, 2226–2230. [Google Scholar] [CrossRef] [PubMed]
- van Lier, H.; Drinkenburg, W.H.; van Eeten, Y.J.; Coenen, A.M. Effects of diazepam and zolpidem on EEG beta frequencies are behavior-specific in rats. Neuropharmacology 2004, 47, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Scheffzuk, C.; Kukushka, V.I.; Vyssotski, A.L.; Draguhn, A.; Tort, A.B.; Brankack, J. Global slowing of network oscillations in mouse neocortex by diazepam. Neuropharmacology 2013, 65, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Drexler, B.; Zinser, S.; Huang, S.; Poe, M.M.; Rudolph, U.; Cook, J.M.; Antkowiak, B. Enhancing the function of alpha5-subunit-containing GABAA receptors promotes action potential firing of neocortical neurons during up-states. Eur. J. Pharmacol. 2013, 703, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanos, P.; Nelson, M.E.; Highland, J.N.; Krimmel, S.R.; Georgiou, P.; Gould, T.D.; Thompson, S.M. A Negative Allosteric Modulator for alpha5 Subunit-Containing GABA Receptors Exerts a Rapid and Persistent Antidepressant-Like Action without the Side Effects of the NMDA Receptor Antagonist Ketamine in Mice. eNeuro 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Walters, R.J.; Hadley, S.H.; Morris, K.D.; Amin, J. Benzodiazepines act on GABAA receptors via two distinct and separable mechanisms. Nat. Neurosci. 2000, 3, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Drexler, B.; Zinser, S.; Hentschke, H.; Antkowiak, B. Diazepam Decreases Action Potential Firing of Neocortical Neurons via Two Distinct Mechanisms. Anesth. Analg. 2010, 111, 1394–1399. [Google Scholar] [CrossRef]
- Tokuda, K.; O’Dell, K.A.; Izumi, Y.; Zorumski, C.F. Midazolam inhibits hippocampal long-term potentiation and learning through dual central and peripheral benzodiazepine receptor activation and neurosteroidogenesis. J. Neurosci. 2010, 30, 16788–16795. [Google Scholar] [CrossRef]
- Rupprecht, R.; Rammes, G.; Eser, D.; Baghai, T.C.; Schule, C.; Nothdurfter, C.; Troxler, T.; Gentsch, C.; Kalkman, H.O.; Chaperon, F.; et al. Translocator protein (18 kD) as target for anxiolytics without benzodiazepine-like side effects. Science 2009, 325, 490–493. [Google Scholar] [CrossRef]
- Drexler, B.; Jurd, R.; Rudolph, U.; Antkowiak, B. Distinct actions of etomidate and propofol at beta3-containing gamma-aminobutyric acid type A receptors. Neuropharmacology 2009, 57, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Shen, B.; Stewart, L.S.; Herrick, I.A.; Leung, L.S. The septohippocampal system participates in general anesthesia. J. Neurosci. 2002, 22, RC200. [Google Scholar] [CrossRef] [PubMed]
- Engin, E.; Benham, R.S.; Rudolph, U. An Emerging Circuit Pharmacology of GABAA Receptors. Trends Pharmacol. Sci. 2018, 39, 710–732. [Google Scholar] [CrossRef] [PubMed]
- Shields, B.C.; Kahuno, E.; Kim, C.; Apostolides, P.F.; Brown, J.; Lindo, S.; Mensh, B.D.; Dudman, J.T.; Lavis, L.D.; Tadross, M.R. Deconstructing behavioral neuropharmacology with cellular specificity. Science 2017, 356. [Google Scholar] [CrossRef] [PubMed]
- Haiss, F.; Butovas, S.; Schwarz, C. A miniaturized chronic microelectrode drive for awake behaving head restrained mice and rats. J. Neurosci. Methods 2010, 187, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, C.; Hentschke, H.; Butovas, S.; Haiss, F.; Stuttgen, M.C.; Gerdjikov, T.V.; Bergner, C.G.; Waiblinger, C. The head-fixed behaving rat—Procedures and pitfalls. Somatosens. Mot. Res. 2010, 27, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Hentschke, H.; Stuttgen, M.C. Computation of measures of effect size for neuroscience data sets. Eur. J. Neurosci. 2011, 34, 1887–1894. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hofmann, J.I.; Schwarz, C.; Rudolph, U.; Antkowiak, B. Effects of Diazepam on Low-Frequency and High-Frequency Electrocortical γ-Power Mediated by α1- and α2-GABAA Receptors. Int. J. Mol. Sci. 2019, 20, 3486. https://doi.org/10.3390/ijms20143486
Hofmann JI, Schwarz C, Rudolph U, Antkowiak B. Effects of Diazepam on Low-Frequency and High-Frequency Electrocortical γ-Power Mediated by α1- and α2-GABAA Receptors. International Journal of Molecular Sciences. 2019; 20(14):3486. https://doi.org/10.3390/ijms20143486
Chicago/Turabian StyleHofmann, Julian I., Cornelius Schwarz, Uwe Rudolph, and Bernd Antkowiak. 2019. "Effects of Diazepam on Low-Frequency and High-Frequency Electrocortical γ-Power Mediated by α1- and α2-GABAA Receptors" International Journal of Molecular Sciences 20, no. 14: 3486. https://doi.org/10.3390/ijms20143486