
Hypermethylation of the 16q23.1 Tumor Suppressor Gene ADAMTS18 in Clear Cell Renal Cell Carcinoma

Abstract
:1. Introduction
2. Results
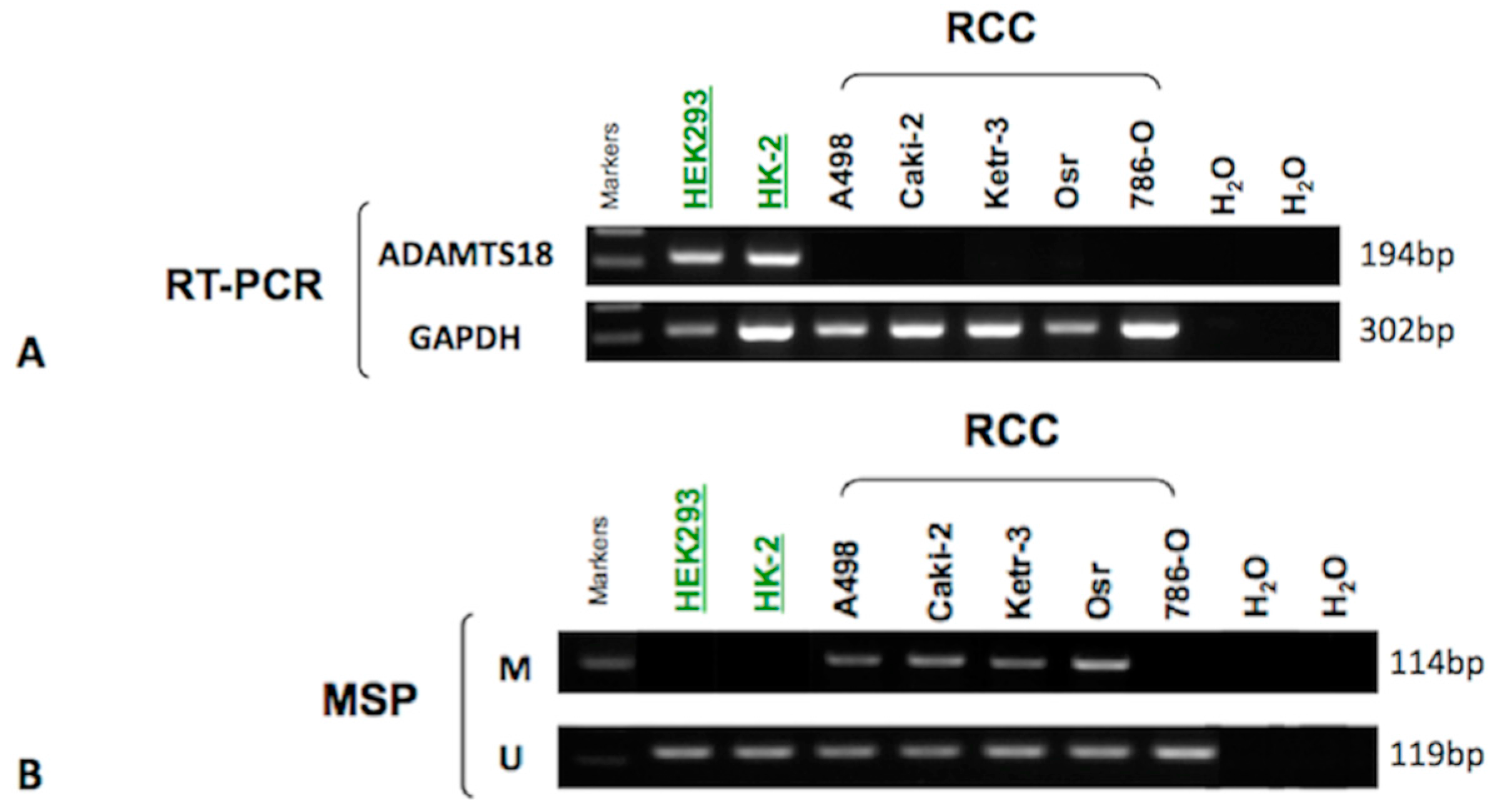
2.1. Down-Regulation of ADAMTS18 Gene Expression in Clear Cell Renal Cell Carcinoma (ccRCC)-Derived Cell Lines

2.2. Hypermethylation of ADAMTS18 in ccRCC-Derived Cell Lines
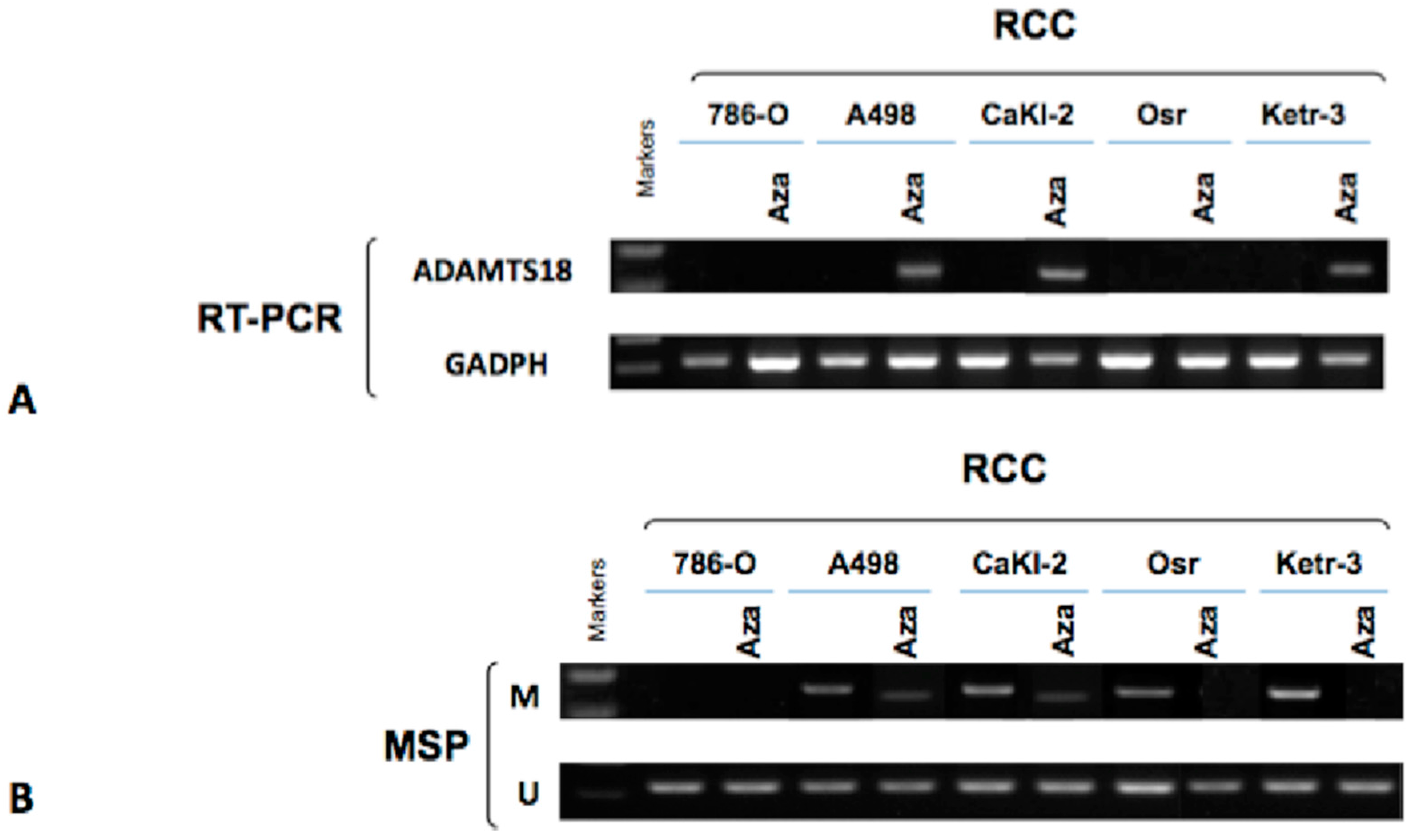
2.3. Activation of ADAMTS18 Gene Expression after Demethylation via Drug Treatment in ccRCC-Derived Cell Lines

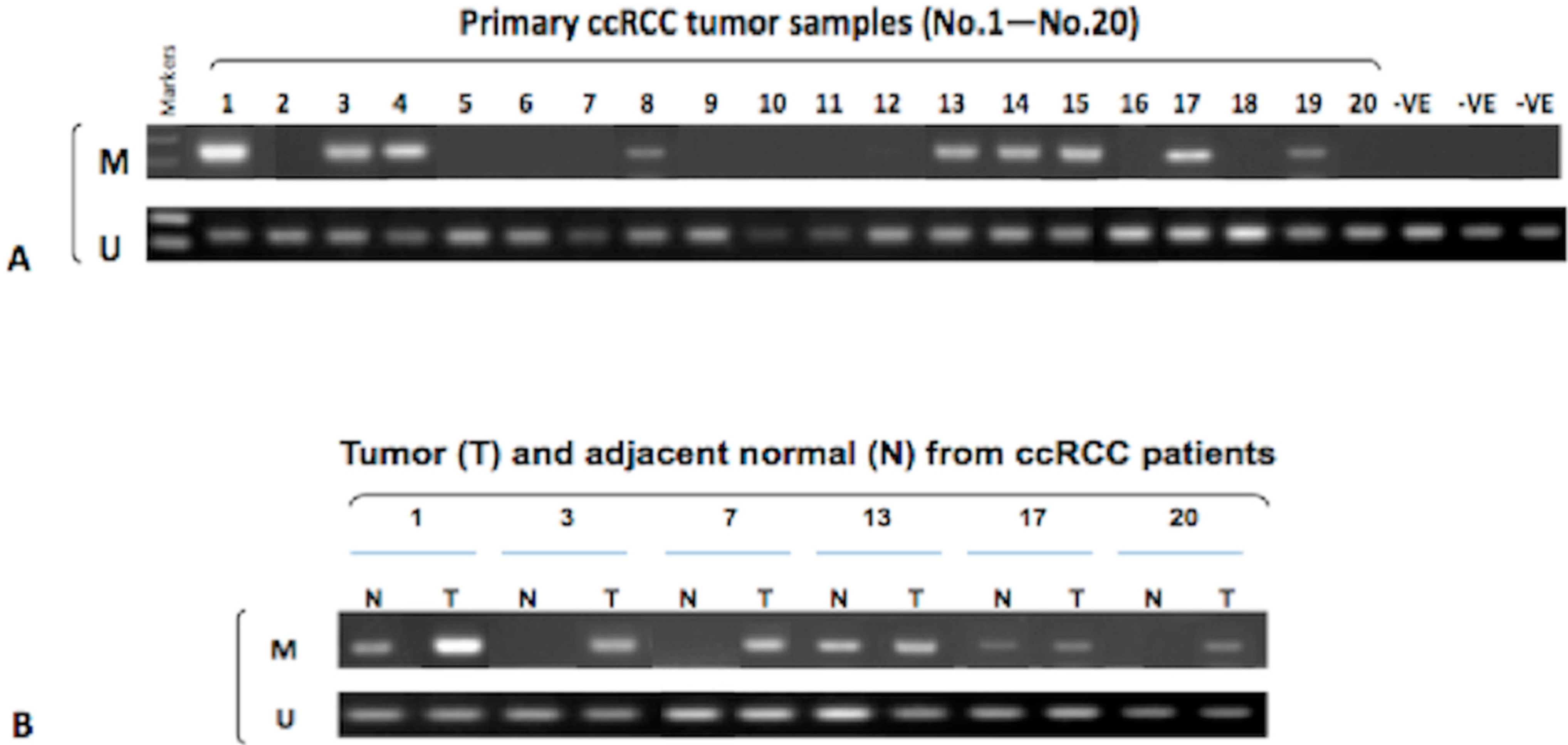
2.4. Hypermethylation of the ADAMTS18 Gene in ccRCC Primary Tumors and Adjacent Normal Tissues

2.5. Bisulfite Genomic Sequencing (BGS) Analysis

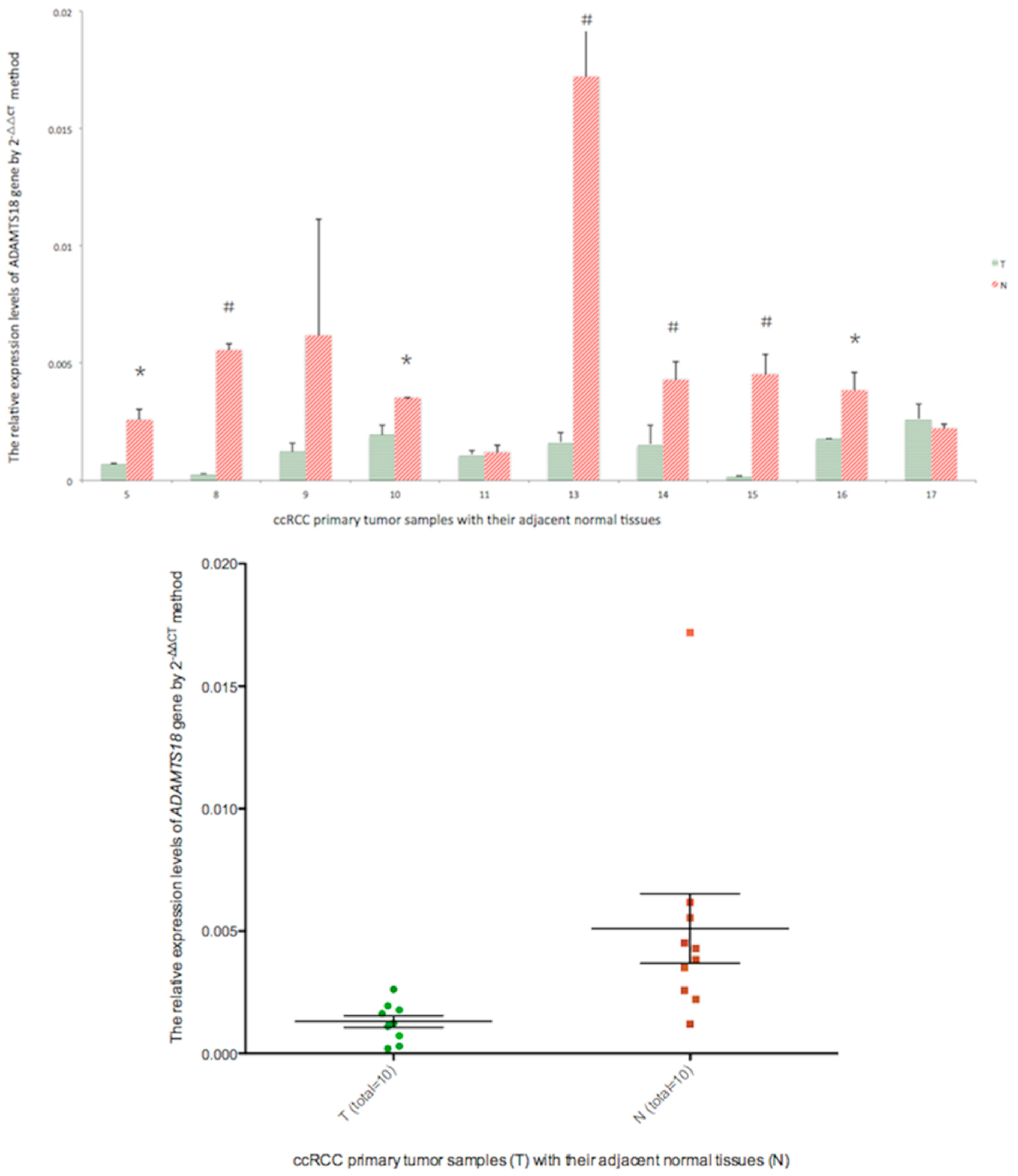
2.6. Real-Time PCR Analysis

2.7. Analysis of Clinicopathological Features

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Features | No. Methylated (%) | No. Unmethylated (%) | p Value | |
|---|---|---|---|---|
| Overall | 44 (43.6%) | 57 (56.4%) | – | |
| Age (years) | 60.6 ± 9.6 | 56.9 ± 12.1 | 0.092 | |
| Gender | M | 35 (49.3%) | 36 (50.7%) | 0.074 |
| F | 9 (30.0%) | 21 (70.0%) | ||
| Side | Left | 27 (45.8%) | 32 (54.2%) | 0.597 |
| Right | 17 (40.5%) | 25 (59.5%) | ||
| Tumor diameters (cm) | 5.9 ± 1.6 | 5.2 ± 2.2 | 0.085 | |
| AJCC pathological stage | I | 27 (39.1%) | 42 (60.9%) | 0.400 |
| II | 3 (60.0%) | 2 (40.0%) | ||
| III | 14 (51.9%) | 13 (48.1%) | ||
| Nuclear grade | G1 | 11 (41.9%) | 17 (58.1%) | 0.502 |
| G2 | 30 (45.9%) | 33 (54.1%) | ||
| G3 | 3 (33.3%) | 7 (66.7%) | ||
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. Cell Line Preparation and Treatment of ccRCC-Derived Cell Lines with a Demethylating Drug
4.3. Semi-Quantitative Reverse Transcription PCR (RT-PCR)
| Gene | PCR | Primer Sequences (5'→3') | Annealing Temperature (°C) | Cycles |
|---|---|---|---|---|
| ADAMTS18 | RT-PCR (real-time PCR) | F: TAGCCAGTGACAGCAGCAG | 55 | 37 |
| R: CTAAGTGCAGTTCCTGTCCA | 55 | 37 | ||
| MSP | m1: TTGTAGTTCGGTAGGTTCGC | 60 | 40 | |
| m2: ACTCCAAATAAAAACCGCCG | ||||
| u1: AAATTGTAGTTTGGTAGGTTTGT | 58 | 38 | ||
| u2: CAACTCCAAATAAAAACCACCA | ||||
| BGS | BGS1: GTTTTAGTTTYGGTTTAGGGAGTT | 60 | 40 | |
| BGS2: AACRCACTCCATAATCAAATAC |
4.4. DNA Extraction from ccRCC Tissue Samples
4.5. Bisulfite Modification of DNA and Methylation-Specific PCR (MSP)
4.6. Bisulfite Genomic Sequencing (BGS) of ADAMTS18 in Cell Lines and Specimens
4.7. Quantitative Real-Time PCR
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Nagata, M.; Sakurai-Yageta, M.; Yamada, D.; Goto, A.; Ito, A.; Fukuhara, H.; Kume, H.; Morikawa, T.; Fukayama, M.; Homma, Y.; et al. Aberrations of a cell adhesion molecule CADM4 in renal clear cell carcinoma. Int. J. Cancer 2012, 130, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.R.; Ricketts, C.; Gentle, D.; Abdulrahman, M.; Clarke, N.; Brown, M.; Kishida, T.; Yao, M.; Latif, F.; Maher, E.R. Identification of candidate tumor suppressor genes frequently methylated in renal cell carcinoma. Oncogene 2010, 29, 2104–2117. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Epigenetics in cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Onay, H.; Pehlivan, S.; Koyuncuoglu, M.; Kirkali, Z.; Ozkinay, F. Multigene methylation analysis of conventional renal cell carcinoma. Urol. Int. 2009, 83, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Sebova, K.; Zmetakova, I.; Bella, V.; Kajo, K.; Stankovicova, I.; Kajabova, V.; Krivulcik, T.; Lasabova, Z.; Tomka, M.; Galbavy, S.; et al. RASSF1A and CDH11 hypermethylation as potential epimarkers in breast cancer. Cancer Biomark. 2011, 10, 13–26. [Google Scholar] [PubMed]
- Carmona, F.J.; Villanueva, A.; Vidal, A.; Muñoz, C.; Puertas, S.; Penin, R.M.; Gomà, M.; Lujambio, A.; Piulats, J.M.; Mesía, R.; et al. Epigenetic disruption of cadherin-11 in human cancer metastasis. J. Pathol. 2012, 228, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, C.J.; Morris, M.R.; Gentle, D.; Brown, M.; Wake, N.; Woodward, E.R.; Clarke, N.; Latif, F.; Maher, E.R. Genome-wide CpG island methylation analysis implicates novel genes in the pathogenesis of renal cell carcinoma. Epigenetics 2012, 7, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Ibragimova, I.; Maradeo, M.E.; Dulaimi, E.; Cairns, P. Aberrant promoter hypermathylation of PBRM1, BAP1, SETD2, KDM6A and other chromatin-modifying genes is absent or rare in clear cell RCC. Epigenetics 2013, 8, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.R.; Maher, E.R. Epigenetics of renal cell carcinoma: The path towards new diagnostics and therapeutics. Genome Med. 2010, 2, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ying, J.; Li, J.; Fan, Y.; Poon, F.F.; Ng, K.M.; Tao, Q.; Jin, J. Aberrant promoter methylation of DLEC1, a critical 3p22 tumor suppressor for renal cell carcinoma, is associated with more advanced tumor stage. J. Urol. 2010, 184, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ying, J.; Zhang, K.; Li, H.; Ng, K.M.; Zhao, Y.; He, Q.; Yang, X.; Xin, D.; Liao, S.K.; et al. Aberrant methylation of the 8p22 tumor suppressor gene DLC1 in renal cell carcinoma. Cancer Lett. 2007, 249, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.R.; Ricketts, C.J.; Gentle, D.; McRonald, F.; Carli, N.; Khalili, H.; Brown, M.; Kishida, T.; Yao, M.; Banks, R.E.; et al. Genome-wide methylation analysis identifies epigenetically inactivated candidate tumor suppressor genes in renal cell carcinoma. Oncogene 2011, 30, 1390–1401. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.; Clark, I.M.; Kevorkian, L.; Edwards, D.R. The ADAMTS metalloproteinases. Biochem. J. 2005, 386, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Wang, X.; Ying, J.; Wong, A.H.; Li, H.; Lee, K.Y.; Srivastava, G.; Chan, A.T.; Yeo, W.; Ma, B.B.; et al. Epigenetic identification of ADAMTS18 as a novel 16q23.1 tumor suppressor frequently silenced in esophageal, nasopharyngeal and multiple other carcinomas. Oncogene 2007, 26, 7490–7498. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, W.; Shao, Y.; Zhang, C.; Wu, Q.; Yang, H.; Wan, X.; Zhang, J.; Guan, M.; Wan, J.; et al. High-resolution melting analysis of ADAMTS18 methylation levels in gastric, colorectal and pancreatic cancers. Med. Oncol. 2010, 27, 998–1004. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, K.; Minagawa, M.; Takatani, T.; Takatani, R.; Ohashi, M.; Kohno, Y. Establishment of diagnosis by bisulfite-treated methylation-specific PCR method and analysis of clinical characteristics of pseudohypoparathyoidism type 1b. Endocr. J. 2011, 58, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, C.J.; Morris, M.R.; Gentle, D.; Shuib, S.; Brown, M.; Clarke, N.; Wei, W.; Nathan, P.; Latif, F.; Maher, E.R. Methylation profiling and evaluation of demethylating therapy in renal cell carcinoma. Clin. Epigenet. 2013, 5, 16. [Google Scholar] [CrossRef]
- Peters, I.; Eggers, H.; Atschekzei, F.; Hennenlotter, J.; Waalkes, S.; Tränkenschuh, W.; Grosshennig, A.; Merseburger, A.S.; Stenzl, A.; Kuczyk, M.A.; et al. GATA5 CpG island methylation in renal cell cancer: A potential biomarker for metastasis and disease progression. BJU Int. 2012, 110, e144–e152. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Wang, Y.L.; Fu, X.L.; Ma, J.G. Aberrant methylation of PCDH8 is a potential prognostic biomarker for patients with clear cell renal cell carcinoma. Med. Sci. Monit. 2014, 20, 2380–2385. [Google Scholar] [CrossRef] [PubMed]
- Kwabi-Addo, B.; Chung, W.; Shen, L.; Ittmann, M.; Wheeler, T.; Jelinek, J.; Issa, J.P. Age-related DNA methylation changes in normal human prostate tissues. Clin. Cancer Res. 2007, 13, 3796–3802. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, R.M.; Snijders, P.J.; Grünberg, K.; Kooi, C.; Spreeuwenberg, M.D.; Meijer, C.J.; Postmus, P.E.; Smit, E.F.; Steenbergen, R.D. Comprehensive CADM1 promoter methylation analysis in NSCLC and normal lung specimens. Lung Cancer 2011, 72, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Wistuba, I.I. Genetics of preneoplasia: Lessons from lung cancer. Curr. Mol. Med. 2007, 7, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.G.; Fan, Y.; Davies, G.; Ying, J.; Geng, H.; Ng, K.M.; Li, H.; Gao, Z.; Wei, W.; Bose, S.; et al. Epigenetic silencing of a proapoptotic cell adhesion molecule, the immunoglobulin superfamily member IGSF4, by promoter CpG methylation protects Hodgkin lymphoma cells from apoptosis. Am. J. Pathol. 2010, 177, 1480–1490. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Li, X.; Xu, S.; Ai, J.; Gong, Y.; Gregg, J.L.; Guan, R.; Qiu, W.; Xin, D.; Gingrich, J.R.; et al. Aberrant methylation and loss of CADM2 tumor suppressor expression is associated with human renal cell carcinoma tumor progression. Biochem. Biophys. Res. Commun. 2013, 435, 526–532. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, B.; Zhang, L.; Luo, C.; Qi, Y.; Cui, Y.; Ying, J.-M.; Zhang, Q.; Jin, J. Hypermethylation of the 16q23.1 Tumor Suppressor Gene ADAMTS18 in Clear Cell Renal Cell Carcinoma. Int. J. Mol. Sci. 2015, 16, 1051-1065. https://doi.org/10.3390/ijms16011051
Xu B, Zhang L, Luo C, Qi Y, Cui Y, Ying J-M, Zhang Q, Jin J. Hypermethylation of the 16q23.1 Tumor Suppressor Gene ADAMTS18 in Clear Cell Renal Cell Carcinoma. International Journal of Molecular Sciences. 2015; 16(1):1051-1065. https://doi.org/10.3390/ijms16011051
Chicago/Turabian StyleXu, Ben, Lian Zhang, Cheng Luo, Yan Qi, Yun Cui, Jian-Ming Ying, Qian Zhang, and Jie Jin. 2015. "Hypermethylation of the 16q23.1 Tumor Suppressor Gene ADAMTS18 in Clear Cell Renal Cell Carcinoma" International Journal of Molecular Sciences 16, no. 1: 1051-1065. https://doi.org/10.3390/ijms16011051