Voluntary Physical Exercise Reduces Motor Dysfunction and Hampers Tumor Cell Proliferation in a Mouse Model of Glioma

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. GL261 Cells and Tumor Induction
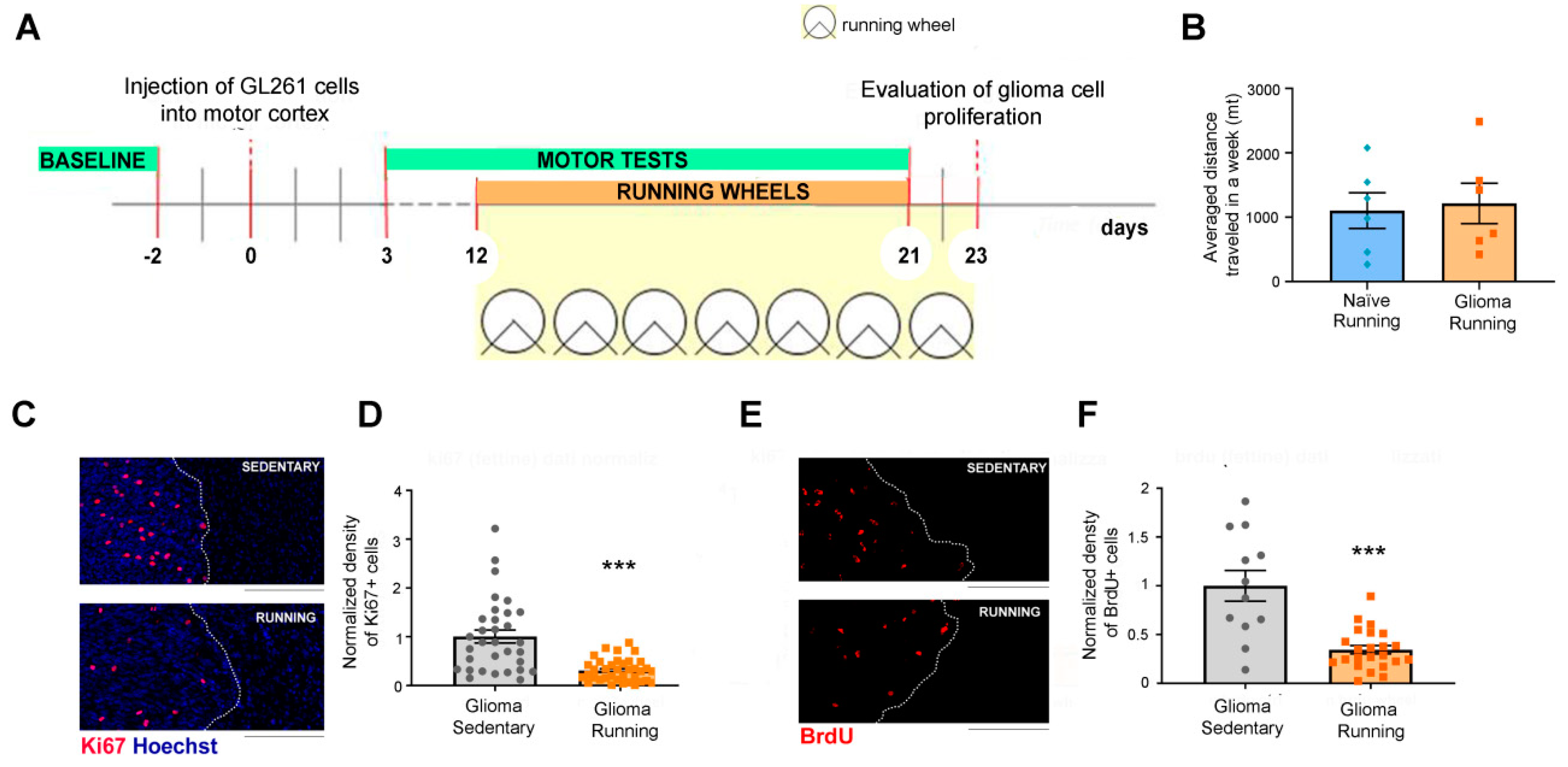
2.3. Running Wheels
2.4. Motor Tests
2.5. Grip Strength Test
2.6. Grid Walk
2.7. Three-Chamber Sociability Test
2.8. Open Field Test
2.9. Nesting Test
2.10. Stereological Reconstruction of the Tumor Volume
2.11. Immunohistochemical Analysis of Glioma Cell Proliferation
2.12. Image Acquisition and Analysis of Tumor Cell Proliferation
2.13. Statistical Analysis
3. Results
3.1. Voluntary Physical Exercise Reduced Glioma Cell Proliferation
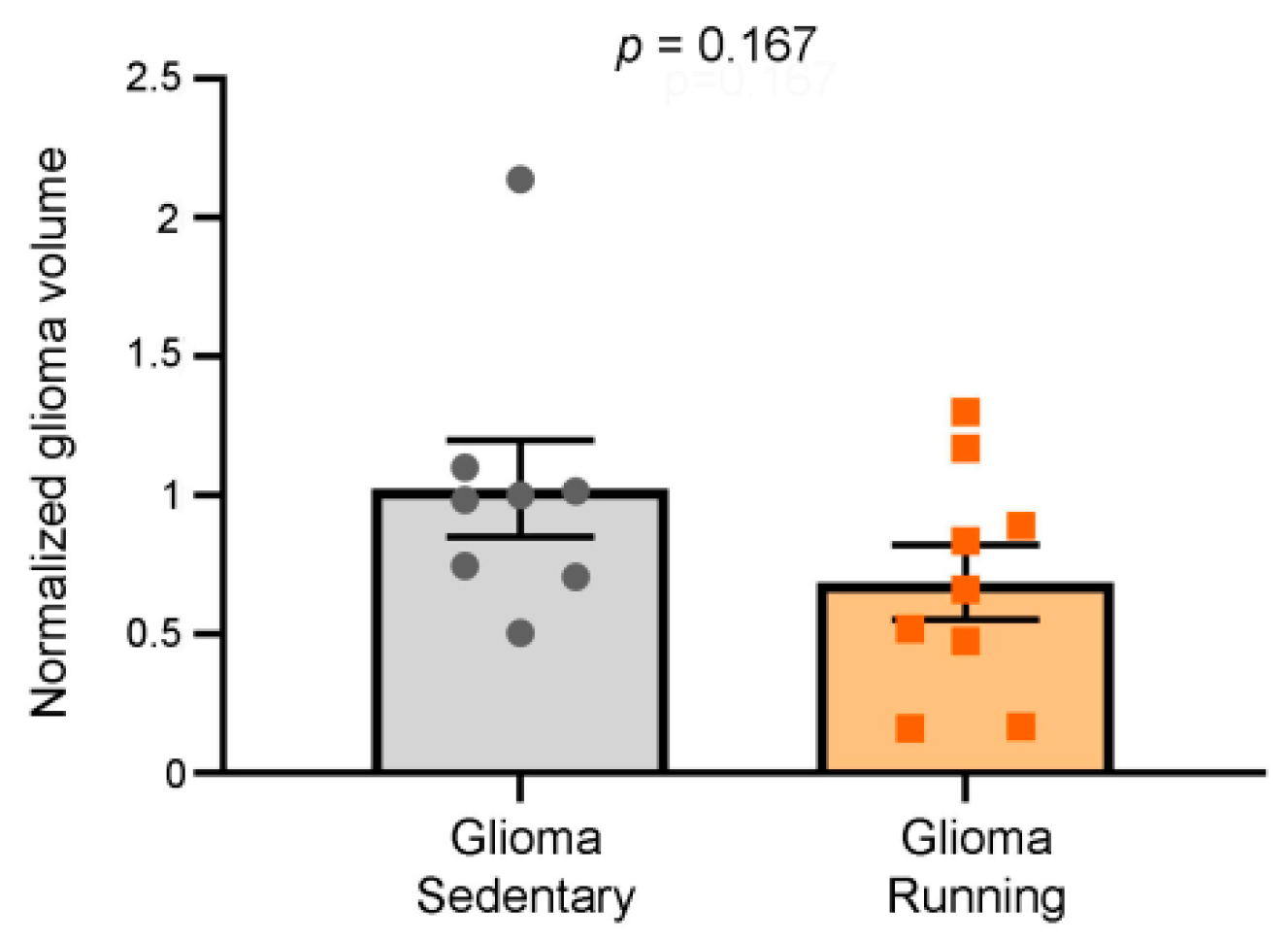
3.2. Comparison of Tumor Volumes in Running vs. Sedentary Glioma-Bearing Animals
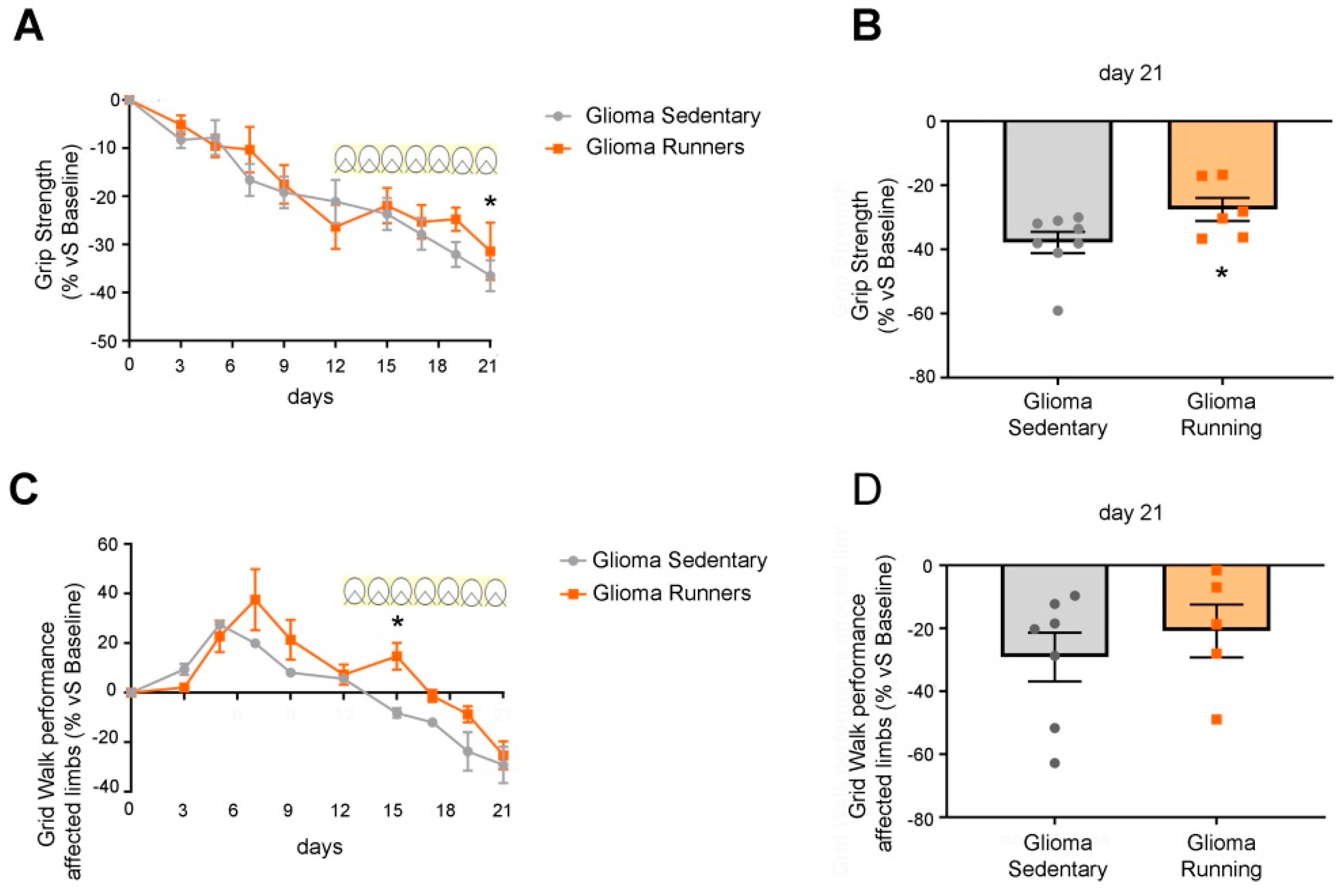
3.3. Delayed Impairment of Motor Functions in Glioma-Bearing Mice That Underwent Voluntary Physical Exercise on Running Wheels
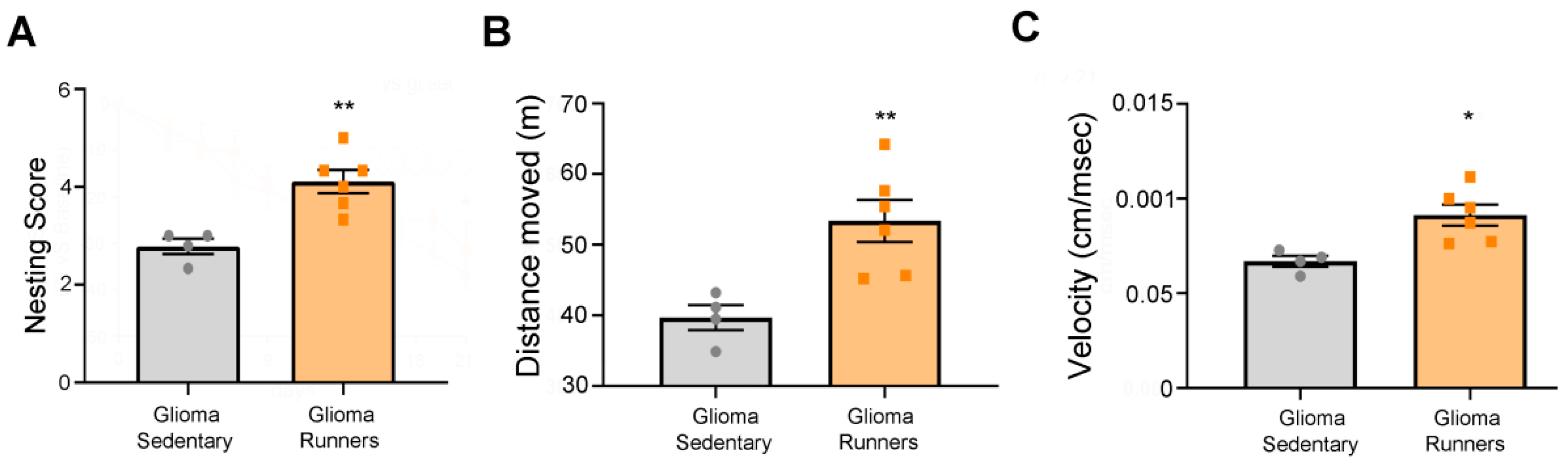
3.4. Behavioral Evaluation of the Physical Exercise Effect on Running and Sedentary Glioma-Bearing Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krech, D.; Rosenzweig, M.R.; Bennett, E.L. Relations between brain chemistry and problem-solving among rats raised in enriched and impoverished environments. J. Comp. Physiol. Psychol. 1962, 55, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.C.; Krech, D.; Rosenzweig, M.R. The effects of an enriched environment on the histology of the rat cerebral cortex. J. Comp. Neurol. 1964, 123, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Consorti, A.; Sansevero, G.; Torelli, C.; Berardi, N.; Sale, A. From basic visual science to neurodevelopmental disorders: The voyage of environmental enrichment-like stimulation. Neural Plast. 2019, 2019, 5653180. [Google Scholar] [CrossRef] [PubMed]
- Baroncelli, L.; Cenni, M.C.; Melani, R.; Deidda, G.; Landi, S.; Narducci, R.; Cancedda, L.; Maffei, L.; Berardi, N. Early IGF-1 primes visual cortex maturation and accelerates developmental switch between NKCC1 and KCC2 chloride transporters in enriched animals. Neuropharmacology 2017, 113, 167–177. [Google Scholar] [CrossRef]
- Sansevero, G.; Sale, A. Environment as therapy: Neuroscience for intellectual disability and dementia. Oncotarget 2017, 8, 5682–5683. [Google Scholar] [CrossRef]
- Baroncelli, L.; Bonaccorsi, J.; Milanese, M.; Bonifacino, T.; Giribaldi, F.; Manno, I.; Cenni, M.C.; Berardi, N.; Bonanno, G.; Maffei, L.; et al. Enriched experience and recovery from amblyopia in adult rats: Impact of motor, social and sensory components. Neuropharmacology 2012, 62, 2388–2397. [Google Scholar] [CrossRef]
- Nithianantharajah, J.; Hannan, A.J. Enriched environments, experience-dependent plasticity and disorders of the nervous system. Nat. Rev. Neurosci. 2006, 7, 697–709. [Google Scholar] [CrossRef]
- Baroncelli, L.; Braschi, C.; Spolidoro, M.; Begenisic, T.; Sale, A.; Maffei, L. Nurturing brain plasticity: Impact of environmental enrichment. Cell Death Differ. 2010, 17, 1092–1103. [Google Scholar] [CrossRef]
- Sale, A.; Berardi, N.; Maffei, L. Environment and brain plasticity: Towards an endogenous pharmacotherapy. Physiol. Rev. 2014, 94, 189–234. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, M.R.; Bennett, E.L. Cerebral changes in rats exposed individually to an enriched environment. J. Comp. Physiol. Psychol. 1972, 80, 304–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baroncelli, L.; Scali, M.; Sansevero, G.; Olimpico, F.; Manno, I.; Costa, M.; Sale, A. Experience affects critical period plasticity in the visual cortex through an epigenetic regulation of histone post-translational modifications. J. Neurosci. 2016, 36, 3430–3440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garofalo, S.; D’Alessandro, G.; Chece, G.; Brau, F.; Maggi, L.; Rosa, A.; Porzia, A.; Mainiero, F.; Esposito, V.; Lauro, C.; et al. Enriched environment reduces glioma growth through immune and non-immune mechanisms in mice. Nat. Commun. 2015, 6, 6623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobilo, T.; Liu, Q.R.; Gandhi, K.; Mughal, M.; Shaham, Y.; van Praag, H. Running is the neurogenic and neurotrophic stimulus in environmental enrichment. Learn. Mem. 2011, 18, 605–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansevero, G.; Begenisic, T.; Mainardi, M.; Sale, A. Experience-dependent reduction of soluble β-amyloid oligomers and rescue of cognitive abilities in middle-age Ts65Dn mice, a model of Down syndrome. Exp. Neurol. 2016, 283, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Downs, J.; Rodger, J.; Li, C.; Tan, X.; Hu, N.; Wong, K.; De Klerk, N.; Leonard, H. Environmental enrichment intervention for Rett syndrome: An individually randomised stepped wedge trial. Orphanet J. Rare Dis. 2018, 13, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balthazar, J.; Schöwe, N.M.; Cipolli, G.C.; Buck, H.S.; Viel, T.A. Enriched environment significantly reduced senile plaques in a transgenic mice model of Alzheimer’s disease, improving memory. Front. Aging Neurosci. 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ball, N.J.; Mercado, E.; Orduña, I. Enriched environments as a potential treatment for developmental disorders: A critical assessment. Front. Psychol. 2019, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Marques-Aleixo, I.; Beleza, J.; Sampaio, A.; Stevanovic, J.; Coxito, P.; Gonçalves, I.; Ascensão, A.; Magalhães, J. Preventive and therapeutic potential of physical exercise in neurodegenerative diseases. Antioxid. Redox Signal. 2020, ars.2020.8075. [Google Scholar] [CrossRef]
- Vivar, C.; Potter, M.C.; Van Praag, H. All About Running: Synaptic Plasticity, Growth Factors and Adult Hippocampal Neurogenesis. In Neurogenesis and Neural Plasticity; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef] [Green Version]
- Sansevero, G.; Torelli, C.; Mazziotti, R.; Consorti, A.; Pizzorusso, T.; Berardi, N.; Sale, A. Running towards amblyopia recovery. Sci. Rep. 2020, 10, 12661. [Google Scholar] [CrossRef]
- Lunghi, C.; Sframeli, A.T.; Lepri, A.; Lepri, M.; Lisi, D.; Sale, A.; Morrone, M.C. A new counterintuitive training for adult amblyopia. Ann. Clin. Transl. Neurol. 2019, 6, 274–284. [Google Scholar] [CrossRef] [Green Version]
- TAMIMI, A.F.; JUWEID, M. Epidemiology and Outcome of Glioblastoma. In Glioblastoma; Codon Publications: Singapore, 2017; pp. 143–153. ISBN 9780994438126. [Google Scholar]
- Ostrom, Q.T.; Cioffi, G.; Gittleman, H.; Patil, N.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2012–2016. Neuro. Oncol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Laub, C.K.; Stefanik, J.; Doherty, L. Approved Treatments for Patients with Recurrent High-grade Gliomas. Semin. Oncol. Nurs. 2018, 34, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Wick, W.; Osswald, M.; Wick, A.; Winkler, F. Treatment of glioblastoma in adults. Ther. Adv. Neurol. Disord. 2018, 11, 175628641879045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannini, E.; Olimpico, F.; Middei, S.; Ammassari-Teule, M.; De Graaf, E.L.; McDonnell, L.; Schmidt, G.; Fabbri, A.; Fiorentini, C.; Baroncelli, L.; et al. Electrophysiology of glioma: A Rho GTPase-activating protein reduces tumor growth and spares neuron structure and function. Neuro. Oncol. 2016, 18, 1634–1643. [Google Scholar] [CrossRef] [Green Version]
- Vannini, E.; Maltese, F.; Olimpico, F.; Fabbri, A.; Costa, M.; Caleo, M.; Baroncelli, L. Progression of motor deficits in glioma-bearing mice: Impact of CNF1 therapy at symptomatic stages. Oncotarget 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tantillo, E.; Vannini, E.; Cerri, C.; Spalletti, C.; Colistra, A.; Mazzanti, C.M.; Costa, M.; Caleo, M. Differential roles of pyramidal and fast-spiking, GABAergic neurons in the control of glioma cell proliferation. Neurobiol. Dis. 2020, 141, 104942. [Google Scholar] [CrossRef]
- Spalletti, C.; Alia, C.; Lai, S.; Panarese, A.; Conti, S.; Micera, S.; Caleo, M. Combining robotic training and inactivation of the healthy hemisphere restores pre-stroke motor patterns in mice. Elife 2017. [Google Scholar] [CrossRef]
- Kaidanovich-Beilin, O.; Lipina, T.; Vukobradovic, I.; Roder, J.; Woodgett, J.R. Assessment of social interaction behaviors. J. Vis. Exp. 2010, 2473. [Google Scholar] [CrossRef]
- Yang, M.; Silverman, J.L.; Crawley, J.N. Automated three-chambered social approach task for mice. Curr. Protoc. Neurosci. 2011. Chapter 8. [Google Scholar] [CrossRef]
- Narducci, R.; Baroncelli, L.; Sansevero, G.; Begenisic, T.; Prontera, C.; Sale, A.; Cenni, M.C.; Berardi, N.; Maffei, L. Early impoverished environment delays the maturation of cerebral cortex. Sci. Rep. 2018, 8, 1187. [Google Scholar] [CrossRef] [Green Version]
- Seibenhener, M.L.; Wooten, M.C. Use of the open field maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 52434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinuesa, A.; Pomilio, C.; Menafra, M.; Bonaventura, M.M.; Garay, L.; Mercogliano, M.F.; Schillaci, R.; Lux Lantos, V.; Brites, F.; Beauquis, J.; et al. Juvenile exposure to a high fat diet promotes behavioral and limbic alterations in the absence of obesity. Psychoneuroendocrinology 2016, 72, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M.J. Assessing nest building in mice. Nat. Protoc. 2006, 1, 1117–1119. [Google Scholar] [CrossRef] [PubMed]
- Vannini, E.; Panighini, A.; Cerri, C.; Fabbri, A.; Lisi, S.; Pracucci, E.; Benedetto, N.; Vannozzi, R.; Fiorentini, C.; Caleo, M.; et al. The bacterial protein toxin, cytotoxic necrotizing factor 1 (CNF1) provides long-term survival in a murine glioma model. BMC Cancer 2014, 14, 1–10. [Google Scholar] [CrossRef]
- Moy, S.S.; Nadler, J.J.; Perez, A.; Barbaro, R.P.; Johns, J.M.; Magnuson, T.R.; Piven, J.; Crawley, J.N. Sociability and preference for social novelty in five inbred strains: An approach to assess autistic-like behavior in mice. Genes, Brain Behav. 2004, 3, 287–302. [Google Scholar] [CrossRef]
- Lemke, D.; Pledl, H.; Zorn, M.; Jugold, M.; Blaes, J.; Löw, S.; Hertenstein, A.; Ott, M.; Sahm, F.; Steffen, A.-C.; et al. Slowing down glioblastoma progression in mice by running or the anti-malarial drug dihydroartemisinin? Induction of oxidative stress in murine glioblastoma therapy. Oncotarget 2016, 7, 56713–56725. [Google Scholar] [CrossRef]
- Ruden, E.; Reardon, D.A.; Coan, A.D.; Herndon, J.E.; Hornsby, W.E.; West, M.; Fels, D.R.; Desjardins, A.; Vredenburgh, J.J.; Waner, E.; et al. Exercise behavior, functional capacity, and survival in adults with malignant recurrent glioma. J. Clin. Oncol. 2011, 29, 2918–2923. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, S.; Porzia, A.; Mainiero, F.; Di Angelantonio, S.; Cortese, B.; Basilico, B.; Pagani, F.; Cignitti, G.; Chece, G.; Maggio, R.; et al. Environmental stimuli shape microglial plasticity in glioma. Elife 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Carson, K.A.; Grossman, S.A.; Fisher, J.D.; Shaw, E.G. Prognostic factors for survival in adult patients with recurrent glioma enrolled onto the new approaches to brain tumor therapy CNS Consortium phase I and II clinical trials. J. Clin. Oncol. 2007, 25, 2601–2606. [Google Scholar] [CrossRef]
- Wu, W.; Lamborn, K.R.; Buckner, J.C.; Novotny, P.J.; Chang, S.M.; O’Fallon, J.R.; Jaeckle, K.A.; Prados, M.D. Joint NCCTG and NABTC prognostic factors analysis for high-grade recurrent glioma. Neuro. Oncol. 2010, 12, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Cormie, P.; Nowak, A.K.; Chambers, S.K.; Galvão, D.A.; Newton, R.U. The potential role of exercise in neuro-oncology. Front. Oncol. 2015, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoladz, J.A.; Pilc, A. The effect of physical activity on the brain derived neurotrophic factor: From animal to human studies. J. Physiol. Pharmacol. 2010, 61, 533–541. [Google Scholar] [PubMed]
- Hamilton, G.F.; Rhodes, J.S. Exercise Regulation of Cognitive Function and Neuroplasticity in the Healthy and Diseased Brain. In Progress in Molecular Biology and Translational Science; Academic Press: Cambridge, MA, USA, 2015; pp. 135, 381–406. [Google Scholar]
- Baek, S.-S. Role of exercise on the brain. J. Exerc. Rehabil. 2016, 12, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Accattato, F.; Greco, M.; Pullano, S.A.; Caré, I.; Fiorillo, A.S.; Pujia, A.; Montalcini, T.; Foti, D.P.; Brunetti, A.; Gulletta, E. Effects of acute physical exercise on oxidative stress and inflammatory status in young, sedentary obese subjects. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simioni, C.; Zauli, G.; Martelli, A.M.; Vitale, M.; Sacchetti, G.; Gonelli, A.; Neri, L.M. Oxidative stress: Role of physical exercise and antioxidant nutraceuticals in adulthood and aging. Oncotarget 2018, 9, 17181–17198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radak, Z.; Marton, O.; Nagy, E.; Koltai, E.; Goto, S. The complex role of physical exercise and reactive oxygen species on brain. J. Sport Heal. Sci. 2013, 2, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Barnes, J.N.; Corkery, A.T. Exercise Improves Vascular Function, but does this Translate to the Brain? Brain Plast. 2018, 4, 65–79. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tantillo, E.; Colistra, A.; Baroncelli, L.; Costa, M.; Caleo, M.; Vannini, E. Voluntary Physical Exercise Reduces Motor Dysfunction and Hampers Tumor Cell Proliferation in a Mouse Model of Glioma. Int. J. Environ. Res. Public Health 2020, 17, 5667. https://doi.org/10.3390/ijerph17165667
Tantillo E, Colistra A, Baroncelli L, Costa M, Caleo M, Vannini E. Voluntary Physical Exercise Reduces Motor Dysfunction and Hampers Tumor Cell Proliferation in a Mouse Model of Glioma. International Journal of Environmental Research and Public Health. 2020; 17(16):5667. https://doi.org/10.3390/ijerph17165667
Chicago/Turabian StyleTantillo, Elena, Antonella Colistra, Laura Baroncelli, Mario Costa, Matteo Caleo, and Eleonora Vannini. 2020. "Voluntary Physical Exercise Reduces Motor Dysfunction and Hampers Tumor Cell Proliferation in a Mouse Model of Glioma" International Journal of Environmental Research and Public Health 17, no. 16: 5667. https://doi.org/10.3390/ijerph17165667