
Associations among Human-Associated Fecal Contamination, Microcystis aeruginosa, and Microcystin at Lake Erie Beaches



Abstract
:1. Introduction
2. Experimental Section
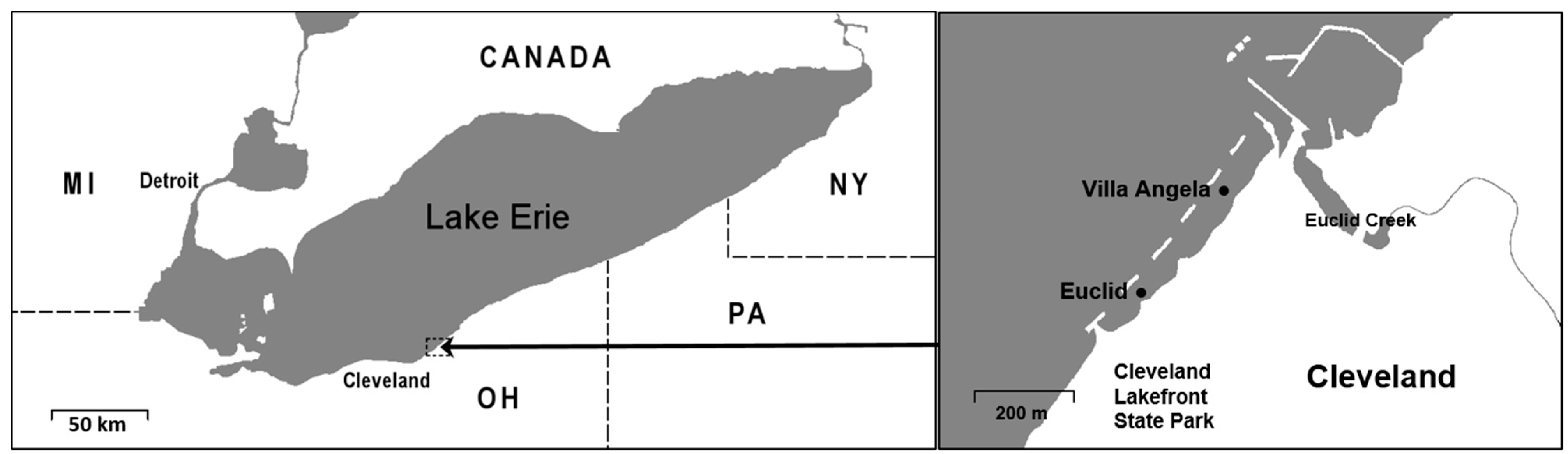
2.1. Site Description and Water Sampling Collection

2.2. Measurement of Nutrients
2.3. Measurement of Microcystin
2.4. DNA Extraction
2.5. Real-Time qPCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay Name | Primer | Sequence (5′-3′) | Amplicon Size (bp) | Gene | Target | Reference |
|---|---|---|---|---|---|---|
| PC-IGS | 188F | GCTACTTCGACCGCGCC | 67 | phycocyanin intergenic spacer | Microcystis aeruginosa | [23] |
| 254R | TCCTACGGTTTAATTGAGACTAGCC | |||||
| mcyA | M1rF | AGCGGTAGTCATTGCATCGG | 107 | mcyA gene | [18] | |
| M1rR | GCCCTTTTTCTGAAGTCGCC | |||||
| g-Bfra | g-Bfra-F | ATAGCCTTTCGAAAGRAAGAT | 501 | 16S rRNA gene | Bacteroides fragilis | [22] |
| g-Bfra-R | CCAGTATCAACTGCAATTTTA |
2.6. Determination of PCR Inhibition in Beach Water Samples
2.7. Statistical Analysis
3. Results
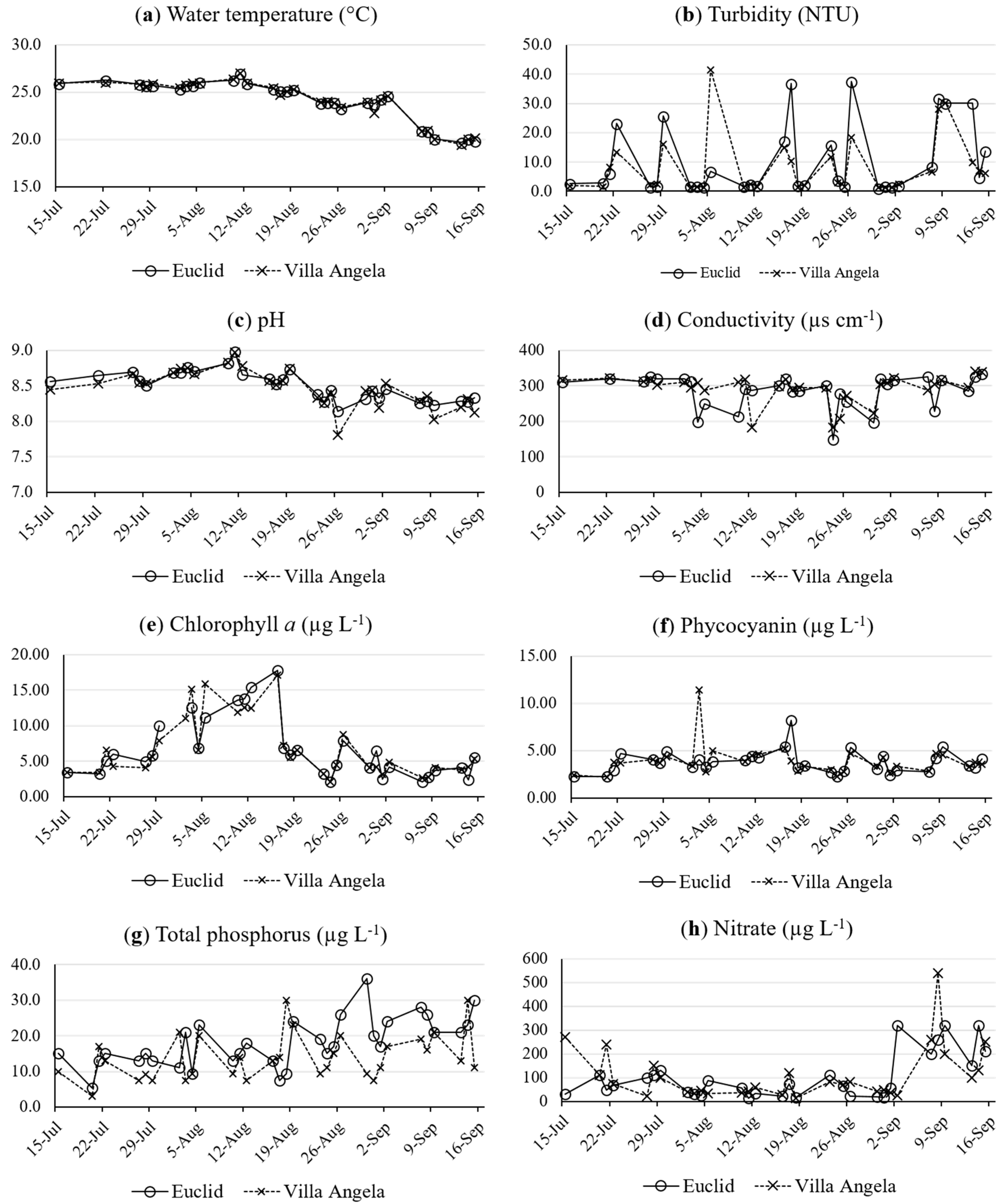
3.1. Water Quality Parameters
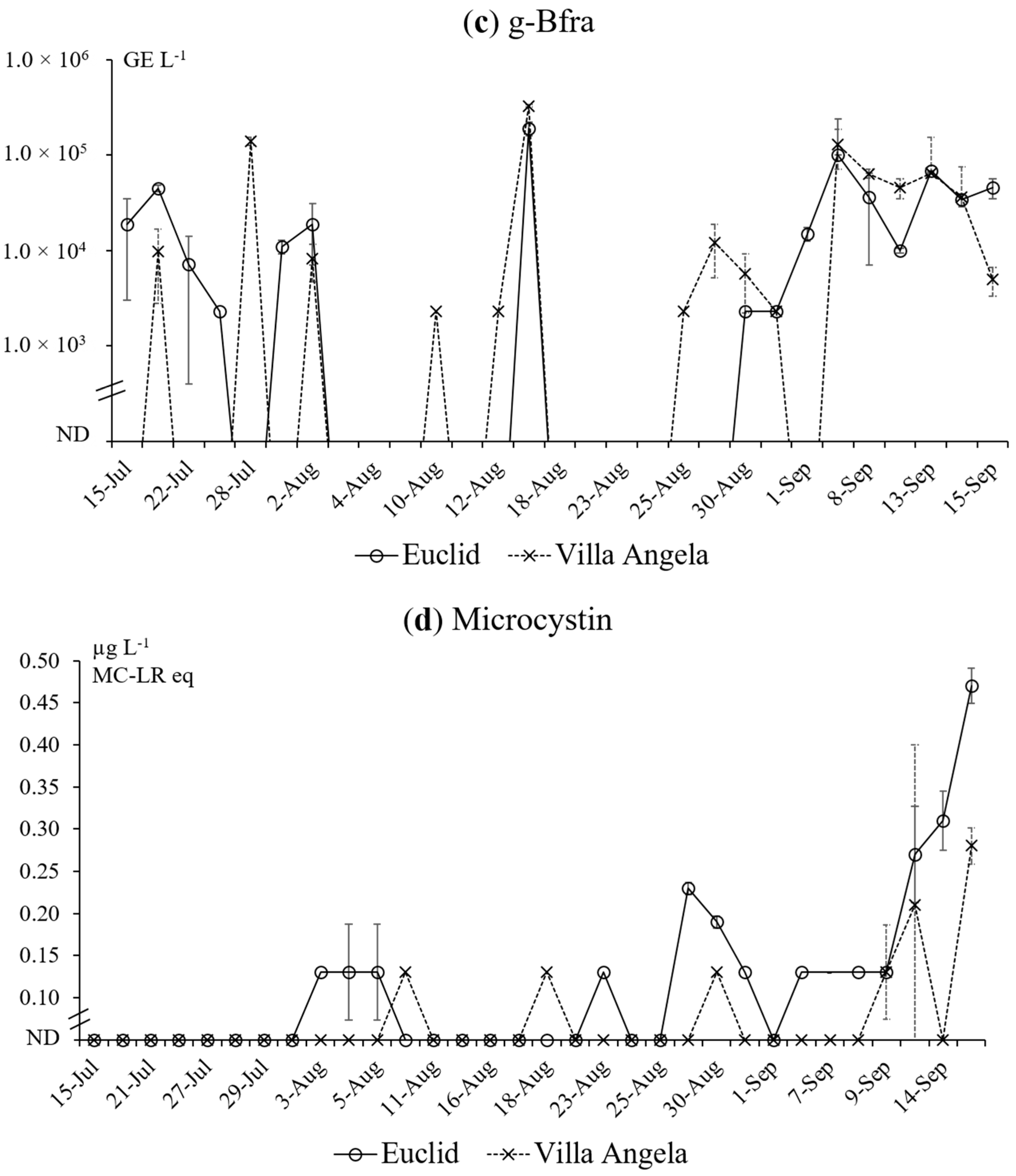
3.2. qPCR Results

| Beach | Statistic | PC-IGS (GE L−1) | mcyA (GE L−1) | g-Bfra (GE·L−1) | Microcystin (μg L−1 MC-LR eq) |
|---|---|---|---|---|---|
| Euclid | Occurrence a | 13/32 (40.6) | 12/32 (37.5) | 16/29 (55.2) | 13/31 (41.9) |
| Mean ± SD b (range) | 1.7 × 104 ± 2.5 × 104 (ND-1.8 × 105 c) | 5.6 × 103 ± 7.7 × 103 (ND-5.9 × 104) | 2.1 × 104 ± 4.0 × 104 (ND-1.9 × 105) | 0.11 ± 0.10 (ND-0.47) | |
| Villa Angela | Occurrence | 19/32 (59.4) | 13/32 (40.6) | 16/29 (55.2) | 6/32 (18.8) |
| Mean ± SD (range) | 3.9 × 104 ± 7.5 × 104 (ND-3.1 × 105) | 1.3 × 104 ± 2.9 × 104 (ND-1.6 × 105) | 3.0 × 104 ± 6.9 × 104 (ND-3.3 × 105) | 0.07 ± 0.053 (ND-0.28) |
3.3. Microcystin Measurement
3.4. Relationships among QPCR Results, Microcystin, and Water Quality Parameters


| Variable | PC-IGS | mcyA | g-Bfra | Microcystin |
|---|---|---|---|---|
| PC-IGS | − | 0.716 *** | 0.411 ** | 0.414 ** |
| mcyA | 0.716 *** | − | 0.535 *** | 0.413 ** |
| g-Bfra | 0.411 ** | 0.535 *** | − | NC |
| Microcystin | 0.414 ** | 0.413 ** | NC | − |
| Water temperature | −0.701 *** | −0.750 *** | −0.542 *** | −0.470 *** |
| Turbidity | NC | 0.384 ** | 0.341 ** | NC |
| pH | −0.632 *** | −0.673 *** | −0.431 ** | −0.369 ** |
| Conductivity | NC | NC | 0.363 ** | NC |
| Chlorophyll a | −0.386 ** | −0.407 ** | −0.325 * | NC |
| Phycocyanin | NC | NC | NC | NC |
| Total phosphorus | NC | 0.336 ** | NC | 0.628 *** |
| Nitrate | 0.462 *** | 0.586 *** | 0.547 *** | NC |
| Dependent Variable | Water Quality Explanatory Variable | Regression Coefficient (β) | Standard Error of β | Standardized Regression Coefficient (β) | t Test Value | Sig. (p) | R2 | Adjusted R2 | Model Sig. |
|---|---|---|---|---|---|---|---|---|---|
| PC-IGS (log) | Constant | 50.743 | 6.387 | 7.945 | <0.001 | 0.519 | 0.511 | <0.001 | |
| Water temperature (log) | −35.320 | 4.623 | −0.721 | −7.640 | <0.001 | ||||
| mcyA (log) | Constant | 43.662 | 6.486 | 6.731 | <0.001 | 0.683 | 0.671 | <0.001 | |
| Water temperature (log) | −31.649 | 4.318 | −0.708 | −7.330 | <0.001 | ||||
| Nitrate (log) | 0.825 | 0.448 | 0.178 | 1.843 | 0.071 | ||||
| g-Bfra (log) | Constant | 29.747 | 10.839 | 2.745 | 0.008 | 0.336 | 0.322 | <0.001 | |
| Water temperature (log) | −21.505 | 7.166 | −0.433 | −3.001 | 0.004 | ||||
| Nitrate (log) | 1.287 | 0.784 | 0.237 | 1.642 | 0.107 | ||||
| Microcystin (log) | Constant | 2.028 | 0.909 | 2.230 | 0.003 | 0.554 | 0.538 | <0.001 | |
| Water temperature (log) | −2.520 | 0.637 | −0.388 | −3.957 | <0.001 | ||||
| Total phosphorus | 0.020 | 0.004 | 0.494 | 5.035 | <0.001 |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bolsenga, S.J.; Herdendorf, C.E. Lake Erie and Lake St. Clair Handbook; Wayne State University Press: Detroit, MI, USA, 1993. [Google Scholar]
- Cordell, K.; Betz, C.J.; Green, G.T.; Mou, S.; Leeworthy, V.R.; Wiley, P.C.; Berry, J.J.; Hellerstein, D. Outdoor Recreation for 21st Century America: A Report to the Nation: The National Survey on Recreation and the Environment; Venture Publishing, Inc.: State College, PA, USA, 2004. [Google Scholar]
- Natural Resources Defense Council. Testing the Waters: A Guide to Water Quality at Vacation Beaches. Available online: http://www.nrdc.org/water/oceans/ttw/oh.asp (accessed on 17 June 2015).
- Ohio Department of Natural Resources. Ohio State Parks 2010 Annual Report. Available online: http://parks.ohiodnr.gov/portals/parks/pdfs/about/2010-annual-report.pdf (accessed on 17 June 2015).
- Ohio Department of Health. Beach monitoring. Available online: http://www.odh.ohio.gov/odhPrograms/eh/bbeach/beachmon.aspx (accessed on 17 June 2015).
- Lee, C.; Agidi, S.; Marion, J.W.; Lee, J. Arcobacter in Lake Erie beach waters: An emerging gastrointestinal pathogen linked with human-associated fecal contamination. Appl. Environ. Microbiol. 2012, 78, 5511–5519. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Marion, J.W.; Lee, J. Development and application of a quantitative PCR assay targeting Catellicoccus marimammalium for assessing gull-associated fecal contamination at Lake Erie beaches. Sci. Total Environ. 2013, 454, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Horst, G.P.; Sarnelle, O.; White, J.D.; Hamilton, S.K.; Kaul, R.B.; Bressie, J.D. Nitrogen availability increases the toxin quota of a harmful cyanobacterium, Microcystis aeruginosa. Water Res. 2014, 54, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Michalak, A.M.; Anderson, E.J.; Beletsky, D.; Boland, S.; Bosch, N.S.; Bridgeman, T.B.; Chaffin, J.D.; Cho, K.; Confesor, R.; Daloglu, I.; et al. Record-setting algal bloom in Lake Erie caused by agricultural and meteorological trends consistent with expected future conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6448–6452. [Google Scholar] [CrossRef] [PubMed]
- Ouellette, A.J.A.; Handy, S.M.; Wilhelm, S.W. Toxic Microcystis is widespread in Lake Erie: PCR detection of toxin genes and molecular characterization of associated cyanobacterial communities. Microb. Ecol. 2006, 51, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Rinta-Kanto, J.M.; Ouellette, A.J.A.; Boyer, G.L.; Twiss, M.R.; Bridgeman, T.B.; Wilhelm, S.W. Quantification of toxic Microcystis spp. during the 2003 and 2004 blooms in western Lake Erie using quantitative real-time PCR. Environ. Sci. Technol. 2005, 39, 4198–4205. [Google Scholar] [CrossRef] [PubMed]
- Budd, J.W.; Beeton, A.M.; Stumpf, R.P.; Culver, D.A.; Kerfoot, W.C. Satellite observations of Microcystis blooms in western Lake Erie. Verh. Internat. Verein. Limnol. 2002, 27, 3787–3793. [Google Scholar]
- Guzman, R.E.; Solter, P.F. Hepatic oxidative stress following prolonged sublethal microcystin LR exposure. Toxicol. Pathol. 1999, 27, 582–588. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Guidelines for Safe Recreational Water Environments—Volume 1: Coastal and Fresh Waters; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- Ohio Environmental Protection Agency. Ohio Algae Information for Recreational Waters. Available online: http://epa.ohio.gov/habalgae.aspx (accessed on 17 June 2015).
- Lehman, P.W.; Teh, S.J.; Boyer, G.L.; Nobriga, M.L.; Bass, E.; Hogle, C. Initial impacts of Microcystis aeruginosa blooms on the aquatic food web in the San Francisco Estuary. Hydrobiologia 2010, 637, 229–248. [Google Scholar] [CrossRef]
- Rinta-Kanto, J.M.; Konopko, E.A.; DeBruyn, J.M.; Bourbonniere, R.A.; Boyer, G.L.; Wilhelm, S.W. Lake Erie Microcystis: Relationship between microcystin production, dynamics of genotypes and environmental parameters in a large lake. Harmful Algae 2009, 8, 665–673. [Google Scholar] [CrossRef]
- Yoshida, M.; Yoshida, T.; Takashima, Y.; Hosoda, N.; Hiroishi, S. Dynamics of microcystin-producing and non-microcystin-producing Microcystis populations is correlated with nitrate concentration in a Japanese lake. FEMS Microbiol. Lett. 2007, 266, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Hong, P.-Y.; Wu, J.-H.; Liu, W.-T. Relative abundance of Bacteroides spp. in stools and wastewaters as determined by hierarchical oligonucleotide primer extension. Appl. Environ. Microbiol. 2008, 74, 2882–2893. [Google Scholar] [CrossRef] [PubMed]
- Ballesté, E.; Blanch, A.R. Persistence of Bacteroides species populations in a river as measured by molecular and culture techniques. Appl. Environ. Microbiol. 2010, 76, 7608–7616. [Google Scholar] [CrossRef] [PubMed]
- Layton, A.; Mckay, L.; Williams, D.; Garrett, V.; Gentry, R.; Sayler, G. Development of Bacteroides 16S rRNA gene TaqMan-based real-time PCR assays for estimation of total, human, and bovine fecal pollution in water. Appl. Environ. Microbiol. 2006, 72, 4214–4224. [Google Scholar] [CrossRef] [PubMed]
- Matsuki, T.; Watanabe, K.; Fujimoto, J.; Miyamoto, Y.; Takada, T.; Matsumoto, K.; Oyaizu, H.; Tanaka, R. Development of 16S rRNA-gene-targeted group-specific primers for the detection and identification of predominant bacteria in human feces. Appl. Environ. Microbiol. 2002, 68, 5445–5451. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Kutzenberger, T. Application of real-time PCR for quantification of microcystin genotypes in a population of the toxic cyanobacterium Microcystis sp. Appl. Environ. Microbiol. 2003, 69, 6723–6730. [Google Scholar] [CrossRef] [PubMed]
- Bertke, E.E. Composite analysis for Escherichia coli at coastal beaches. J. Great Lakes Res. 2007, 33, 335–341. [Google Scholar] [CrossRef]
- Francy, D.S.; Gifford, A.M.; Darner, R.A. Escherichia coli at Ohio Bathing Beaches—Distribution, Sources, Wastewater Indicators, and Predictive Modeling; U.S. Geological Survey: Columbus, OH, USA, 2003.
- National Oceanic and Atmospheric Administration; National Centers for Environmental Information. Daily Summaries Station Details. Available online: http://www.webcitation.org/6bRGTqK0q (accessed on 10 September 2015).
- Marion, J.W.; Lee, J.; Wilkins, J.R., III; Lemeshow, S.; Lee, C.; Waletzko, E.J.; Buckley, T.J. In vivo phycocyanin flourometry as a potential rapid screening tool for predicting elevated microcystin concentrations at eutrophic lakes. Environ. Sci. Technol. 2012, 46, 4523–4531. [Google Scholar] [CrossRef] [PubMed]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Lehman, E.M. Seasonal occurrence and toxicity of Microcystis in impoundments of the Huron River, Michigan, USA. Water Res. 2007, 41, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Neilan, B.A. Xanthogenate acid isolation from cultured and environmental cyanobacteria. J. Phycol. 2000, 36, 251–258. [Google Scholar] [CrossRef]
- Lee, C.S.; Lee, C.; Marion, J.; Wang, Q.; Saif, L.; Lee, J. Occurrence of human enteric viruses at freshwater beaches during swimming season and its link to water inflow. Sci. Total Environ. 2014, 472, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Kephart, C.M.; Bushon, R.N. Utility of Microbial Source-Tracking Markers for Assessing Fecal Contamination in the Portage River Watershed, Northwestern Ohio, 2008; U.S. Geological Survey: Reston, VA, USA, 2010.
- Haugland, R.A.; Siefring, S.C.; Wymer, L.J.; Brenner, K.P.; Dufour, A.P. Comparison of Enterococcus measurements in freshwater at two recreational beaches by quantitative polymerase chain reaction and membrane filter culture analysis. Water Res. 2005, 39, 559–568. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency. Method A: Enterococci in Water by TaqMan® Quantitative Polymerase Chain Reaction (qPCR) Assay; U.S. Environmental Protection Agency, Office of Water (4303T): Washington, DC, USA, 2010.
- Bendel, R.B.; Afifi, A.A. Comparison of stopping rules in forward stepwise regression. J. Am. Stat. Assoc. 1977, 72, 46–53. [Google Scholar]
- Rantala, A.; Rajaniemi-Wacklin, P.; Lyra, C.; Lepistö, L.; Rintala, J.; Mankiewicz-Boczek, J.; Sivonen, K. Detection of microcystin-producing cyanobacteria in Finnish lakes with genus-specific microcystin synthetase gene E (mcyE) PCR and associations with environmental factors. Appl. Environ. Microbiol. 2006, 72, 6101–6110. [Google Scholar] [CrossRef] [PubMed]
- Te, S.H.; Gin, K.Y.-H. The dynamics of cyanobacteria and microcystin production in a tropical reservoir of Singapore. Harmful Algae 2011, 10, 319–329. [Google Scholar]
- Carlson, R.E.; Simpson, J. A Coordinator’s Guide to Volunteer Lake Monitoring Methods; North American Lake Management Society: Madison, WI, USA, 1996. [Google Scholar]
- Chaffin, J.D.; Bridgeman, T.B. Organic and inorganic nitrogen utilization by nitrogen-stressed cyanobacteria during bloom conditions. J. Appl. Phycol. 2014, 26, 299–309. [Google Scholar] [CrossRef]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Farnsley, S.E.; LeCleir, G.R.; Layton, A.C.; Satchwell, M.F.; DeBruyn, J.M.; Boyer, G.L.; Zhu, G.; Paerl, H.W. The relationships between nutrients, cyanobacterial toxins and the microbial community in Taihu (Lake Tai), China. Harmful Algae 2011, 10, 207–215. [Google Scholar] [CrossRef]
- Robarts, R.D.; Zohary, T. Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. N. Z. J. Mar. Freshw. Res. 1987, 21, 391–399. [Google Scholar] [CrossRef]
- Kardinaal, W.E.A.; Visser, P.M. Dynamics of cyanobacterial toxins: Sources of variability in microcystin concentrations. In Harmful Cyanobacteria; Huisman, J., Matthijs, H.C.P., Visser, P.M., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 41–63. [Google Scholar]
- Li, S.; Xie, P.; Xu, J.; Zhang, X.; Qin, J.; Zheng, L.; Liang, G. Factors shaping the pattern of seasonal variations of microcystins in Lake Xingyun, a subtropical plateau lake in China. Bull. Environ. Contam. Toxicol. 2007, 78, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Van der Westhuizen, A.J.; Eloff, J.N. Effect of temperature and light on the toxicity and growth of the blue-green alga Microcystis aeruginosa (UV-006). Planta 1985, 163, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Kong, F.; Zhang, M.; Yang, Z.; Shi, X.; Du, M. The dynamics of Microcystis genotypes and microcystin production and associations with environmental factors during blooms in Lake Chaohu, China. Toxins 2014, 6, 3238–3257. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Tsuji, K.; Watanabe, M.F.; Kondo, F. Stability of microcystins from cyanobacteria—III. Effect of pH and temperature. Phycologia 1996, 35, 83–88. [Google Scholar] [CrossRef]
- Kotak, B.G.; Lam, A.K.; Prepas, E.E.; Hrudey, S.E. Role of chemical and physical variables in regulating microcystin-LR concentration in phytoplankton of eutrophic lakes. Can. J. Fish. Aquat. Sci. 2000, 57, 1584–1593. [Google Scholar] [CrossRef]
- Gągała, I.; Izydorczyk, K.; Jurczak, T.; Pawełczyk, J.; Dziadek, J.; Wojtal-Frankiewicz, A.; Jóźwik, A.; Jaskulska, A.; Mankiewicz-Boczek, J. Role of environmental factors and toxic genotypes in the regulation of microcystins-producing cyanobacterial blooms. Microb. Ecol. 2014, 67, 465–479. [Google Scholar]
- Makarewicz, J.C.; Boyer, G.L.; Lewis, T.W.; Guenther, W.; Atkinson, J.; Arnold, M. Spatial and temporal distribution of the cyanotoxin microcystin-LR in the Lake Ontario ecosystem: coastal embayments, rivers, nearshore and offshore, and upland lakes. J. Great Lakes Res. 2009, 35, 83–89. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.; Marion, J.W.; Cheung, M.; Lee, C.S.; Lee, J. Associations among Human-Associated Fecal Contamination, Microcystis aeruginosa, and Microcystin at Lake Erie Beaches. Int. J. Environ. Res. Public Health 2015, 12, 11466-11485. https://doi.org/10.3390/ijerph120911466
Lee C, Marion JW, Cheung M, Lee CS, Lee J. Associations among Human-Associated Fecal Contamination, Microcystis aeruginosa, and Microcystin at Lake Erie Beaches. International Journal of Environmental Research and Public Health. 2015; 12(9):11466-11485. https://doi.org/10.3390/ijerph120911466
Chicago/Turabian StyleLee, Cheonghoon, Jason W. Marion, Melissa Cheung, Chang Soo Lee, and Jiyoung Lee. 2015. "Associations among Human-Associated Fecal Contamination, Microcystis aeruginosa, and Microcystin at Lake Erie Beaches" International Journal of Environmental Research and Public Health 12, no. 9: 11466-11485. https://doi.org/10.3390/ijerph120911466