Heterologous Expression of the DREB Transcription Factor AhDREB in Populus tomentosa Carrière Confers Tolerance to Salt without Growth Reduction under Greenhouse Conditions

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Salt Treatments
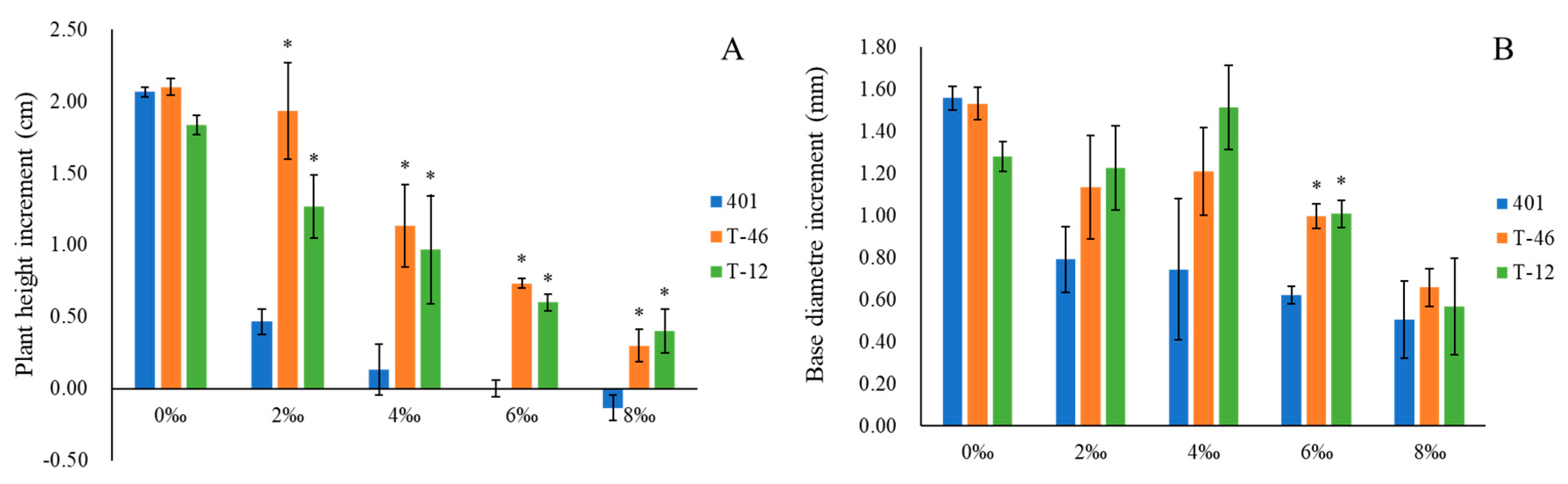
2.2. Determination of the Increment of Growth Parameters
2.3. Determination of Electrolyte Leakage and Malondialdehyde (MDA) Concentration
2.4. Measurement of Leaf Gas Exchange
2.5. Measurement of Proline, SOD, POD and Chl Content
2.6. Cdna Library Preparation and Illumina Transcriptomic Sequencing
2.7. Analysis of Illumina Transcriptomic Sequencing Results
2.8. Differential Expression Analysis and Function Enrichment
3. Results
3.1. Growth under Increasing Levels of Soil Salinity
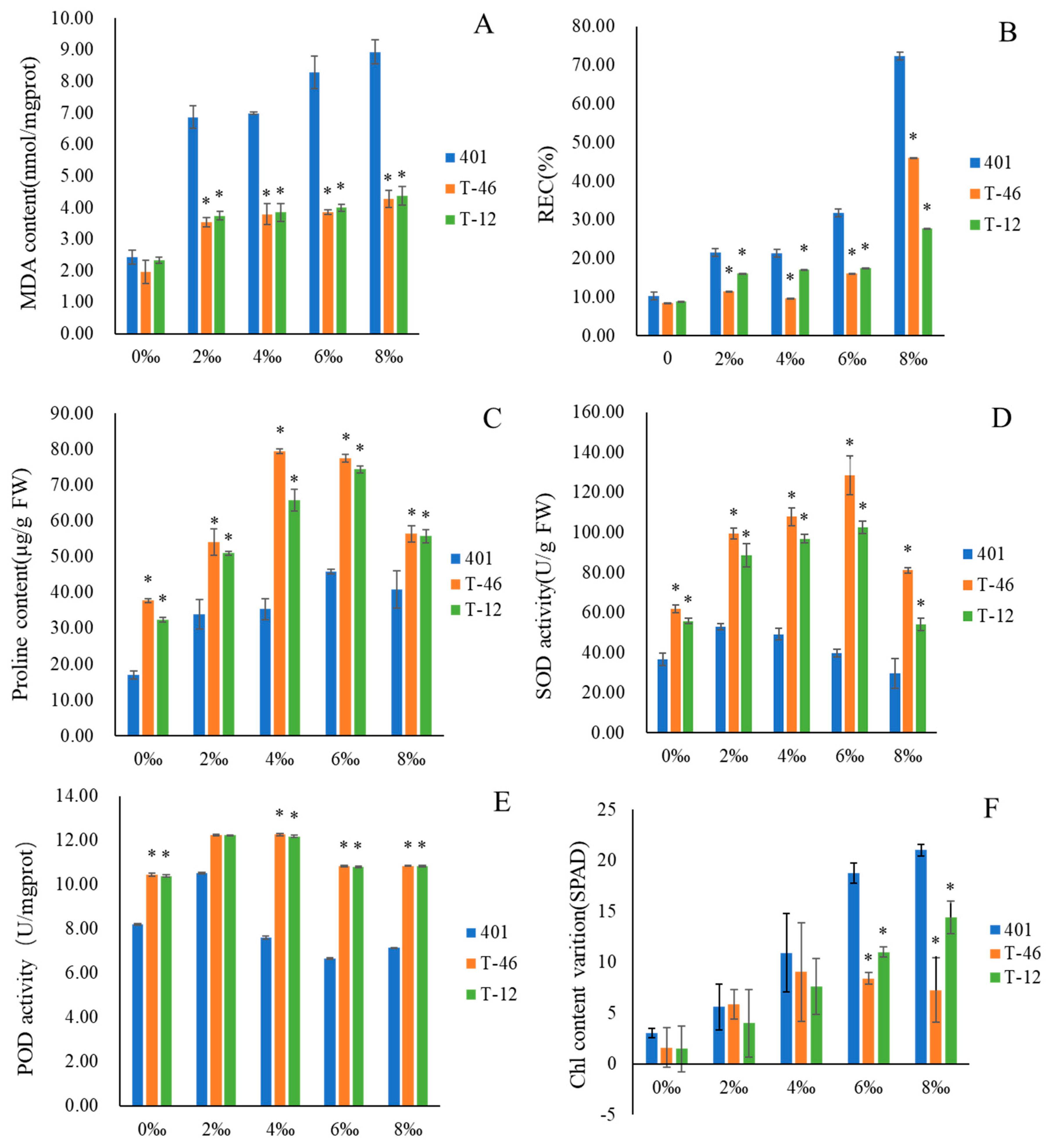
3.2. Effect of Ahdreb Expression on Rate of Electrolyte Penetration (REC) and MDA Contents
3.3. Effect of Ahdreb Expression on Chl Content Variation, SOD and POD Activities And Proline Content
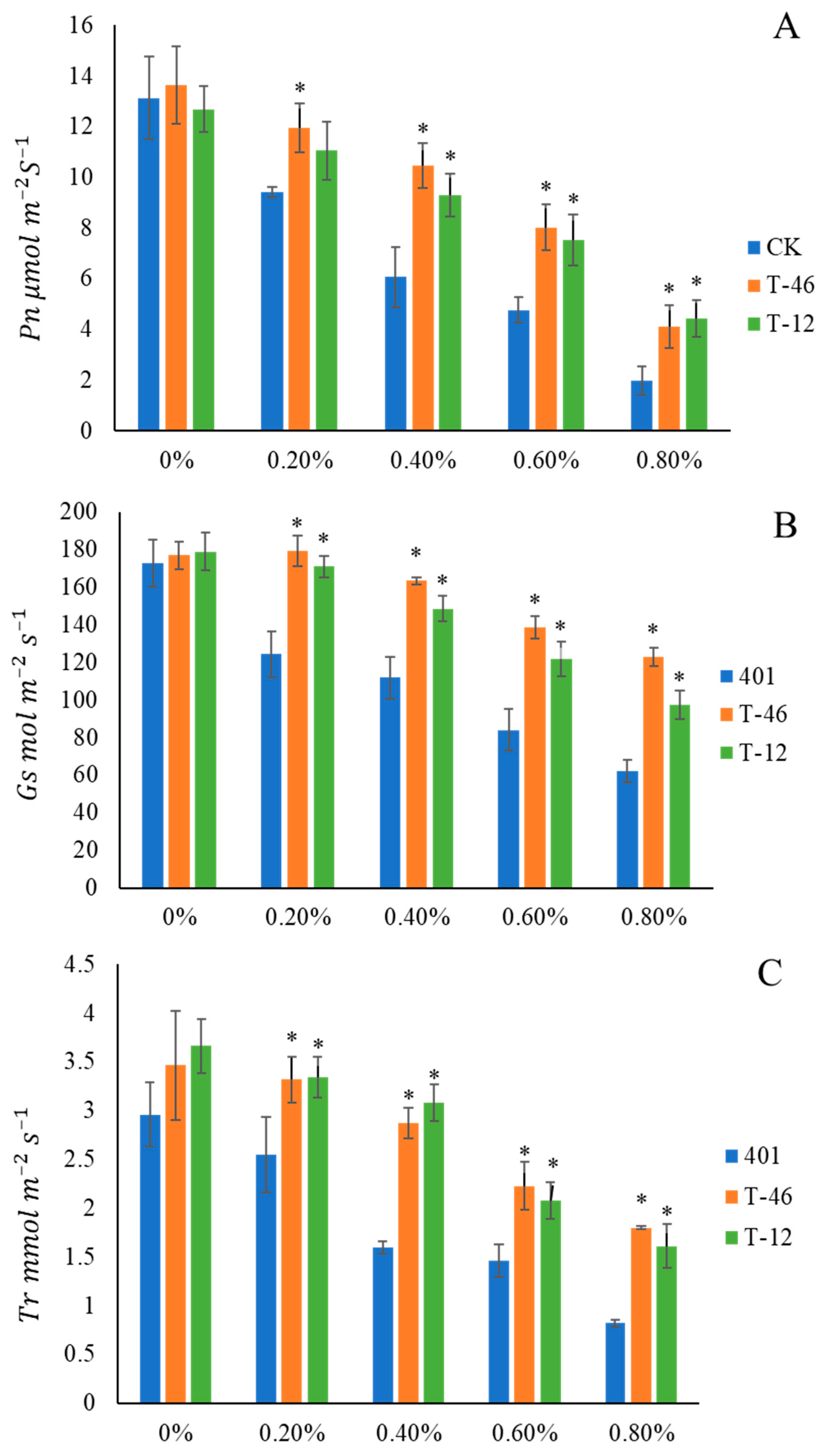
3.4. Photosynthetic Capacity of Transgenic Plants Under Salt Stress Conditions
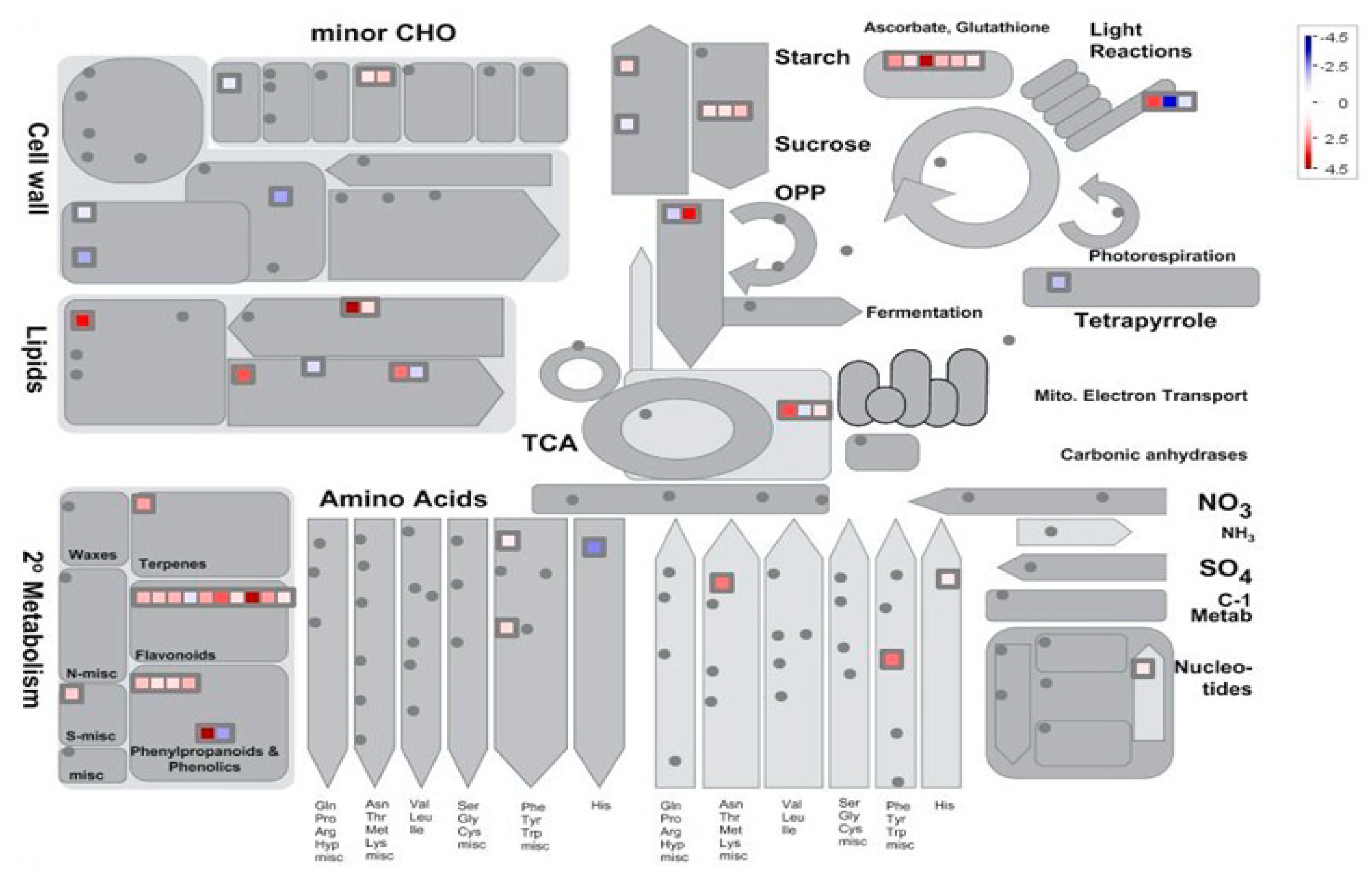
3.5. Transcriptomic Analysis of Ahdreb-Overexpressing Populus Tomentosa Under Salt Stress
3.6. Effect of Ahdreb Overexpression on the Transcriptome in the Absence of Salt
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Zhu, Z.T.; Zhang, Z.Y. The status and advances of genetic improvement of Populus tomentosa Carr. J. Beijing For. Univ. 1997, 6, 1–7. [Google Scholar]
- Li, J.G.; Pu, L.J.; Han, M.J.; Zhu, M.; Zhang, R.S.; Xiang, Y.Z. Soil salinization research in China: Advances and prospects. J. Geogr. Sci. 2014, 24, 943–960. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Li, Z.G.; Hoque, T.S.; Burritt, D.J.; Fujita, M.; Munné-Bosch, S. Heat or cold priming-induced cross-tolerance to abiotic stresses in plants: key regulators and possible mechanisms. Protoplasma 2017, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.Q.; Hou, P.C.; Shen, X.; Wang, M.J.; Deng, S.R.; Sun, J.; Xiao, F.; Wang, R.G.; Zhou, X.Y.; Lu, C.F.; et al. Salt-induced expression of genes related to Na+/K+ and ROS homeostasis in leaves of salt-resistant and salt-sensitive poplar species. Plant Mol. Biol. 2010, 73, 251–269. [Google Scholar] [CrossRef] [PubMed]
- Hmida-Sayari, A.; Gargouri-Bouzid, R.; Bidani, A.; Jaoua, L.; Savoure, A.; Jaoua, S. Overexpression of delta (1)-pyrroline-5-carboxylate synthetase increases proline production and confers salt tolerance in transgenic potato plants. Plant Sci. 2005, 169, 746–752. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chen, Q.J.; Wang, W.L.; Wang, X.C.; Lu, M.Z. Salt tolerance conferred by over-expression of OsNHX1 gene in Poplar 84K. Chin. Sci. Bull. 2005, 50, 225–229. [Google Scholar] [CrossRef]
- Shih, M.D.; Hoekstra, F.A.; Hsing, Y.I.C. Late embryogenesis abundant proteins. Adv. Bot. Res. 2008, 48, 211–255. [Google Scholar] [CrossRef]
- Kumar, V.; Shriram, V.; Kishor, P.B.K.; Narendra, J.L.; Shitole, M.G. Enhanced proline accumulation and salt stress tolerance of transgenic indica rice by over-expressing P5CSF129A gene. Plant Biotechnol. Rep. 2010, 4, 37–48. [Google Scholar] [CrossRef]
- Lan, T.; Gao, J.; Zeng, Q.Y. Genome-wide analysis of the LEA (late embryogenesis abundant) protein gene family in Populus trichocarpa. Tree Genet. Genomes 2013, 9, 253–264. [Google Scholar] [CrossRef]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Lu, H.J.; Wang, X.X.; Cai, X.Y.; Zhou, Z.L.; Zhang, Z.M.; Salih, H.; Wang, K.B.; et al. Characterization of the late embryogenesis abundant (LEA) proteins family and their role in drought stress tolerance in upland cotton. BMC Genet. 2018, 19, 6. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, T.; Shinozaki, K. Research on plant abiotic stress responses in the post-genome era: Past, present and future. Plant J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Jain, D.; Chattopadhyay, D. Role of DREB-like proteins in improving stress tolerance of transgenic crops. In Plant Acclimation to Environmental Stress; Springer: New York, NY, USA, 2013; pp. 147–161. [Google Scholar] [CrossRef]
- Hu, L.; Lu, H.; Liu, Q.; Chen, X.M.; Jiang, X.N. Overexpression of mtlD gene in transgenic Populus tomentosa improves salt tolerance through accumulation of mannitol. Tree Physiol. 2005, 25, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Qu, G.Z.; Li, H.Y.; Wu, Y.J.; Wang, C.; Liu, G.F.; Yang, C.P. Enhanced salt tolerance of transgenic poplar plants expressing a manganese superoxide dismutase from Tamarix androssowii. Mol. Biol. Rep. 2010, 37, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Wang, S.; Zhou, B.; Zhou, B.R.; Jiang, T.B. Transgenic poplar overexpressing the endogenous transcription factor ERF76 gene improves salinity tolerance. Tree Physiol. 2016, 36, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration-and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt-and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Li, X.P.; Tian, A.G.; Luo, G.Z.; Gong, Z.Z.; Zhang, J.S.; Chen, S.Y. Soybean DRE-binding transcription factors that are responsive to abiotic stresses. Theor. Appl. Genet. 2005, 110, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.X.; Wang, X.L.; Wang, H.W.; Xin, H.B.; Yang, X.H.; Yan, J.B.; Li, J.S.; Tran, L.P.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; et al. Genome-wide analysis of ZmDREB genes and their association with natural variation in drought tolerance at seedling stage of Zea mays L. PloS Genet. 2013, 9, e1003790. [Google Scholar] [CrossRef] [PubMed]
- Artlip, T.S.; Wisniewski, M.E.; Bassett, C.L.; Norelli, J.L. CBF gene expression in peach leaf and bark tissues is gated by a circadian clock. Tree Physiol. 2013, 33, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.L.; Lv, H.; Li, L.; Mu, S.H.; Li, X.P.; Gao, J. Genome-wide analysis of the AP2/ERF transcription factors family and the expression patterns of DREB genes in Moso Bamboo (Phyllostachys edulis). PLoS ONE 2015, 10, e0126657. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Guo, X.; Wang, Q.; Wang, Y.T.; Zhao, D.; Yao, L.P.; Wang, S.; Liu, G.J.; Li, T.H. Overexpression of MsDREB6.2 results in cytokinin-deficient developmental phenotypes and enhances drought tolerance in transgenic apple plants. Plant J. 2017, 89, 510–526. [Google Scholar] [CrossRef] [PubMed]
- Atia, A.; Barhoumi, Z.; Debez, A.; Hkiri, S.; Abdelly, C.; Smaoui, A.; Chaffei, C.; Gouia, H. Plant Hormones: Potent Targets for Engineering Salinity Tolerance in Plants. In Salinity Responses and Tolerance in Plants; Springer: New York, NY, USA, 2018; Volume 1, pp. 159–184. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Agarwal, P.; Reddy, M.K.; Sopory, S.K. Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant Cell Rep. 2006, 25, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol. 2018, 149, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.Y.; Niu, F.J.; Sun, X.J.; Hu, Z.; Li, X.H.; Ma, Y.Z.; Zhang, H. RNA-seq analysis of unintended effects in transgenic wheat overexpressing the transcription factor GmDREB1. Crop J. 2017, 5, 207–218. [Google Scholar] [CrossRef]
- Miki, B.; Abdeen, A.; Manabe, Y.; MacDonald, P. Selectable marker genes and unintended changes to the plant transcriptome. Plant Biotechnol. J. 2009, 7, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Abdeen, A.; Schnell, J.; Miki, B. Transcriptome analysis reveals absence of unintended effects in drought-tolerant transgenic plants overexpressing the transcription factor ABF3. BMC Genom. 2010, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Quan, R.D.; Shang, M.; Zhang, H.; Zhao, Y.X.; Zhang, J.R. Improved chilling tolerance by transformation with betA gene for the enhancement of glycinebetaine synthesis in maize. Plant Sci. 2004, 166, 141–149. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Becana, M.; Aparicio-Tejo, P.; Irigoyen, J.J.; Sanchez-Diaz, M. Some enzymes of hydrogen peroxide metabolism in leaves and root nodules of Medicago sativa. Plant Physiol. 1986, 82, 1169–1171. [Google Scholar] [CrossRef] [PubMed]
- Civello, P.M.; Martinez, G.A.; Chaves, A.R.; Anon, M.C. Peroxidase from strawberry fruit (Fragaria ananassa Duch.): partial purification and determination of some properties. J. Agric. Food Chem. 1995, 43. [Google Scholar] [CrossRef]
- Sun, H.P.; Li, F.; Xu, Z.J.; Sun, M.L.; Cong, H.Q.; Qiao, F.; Zhong, X.H. De novo leaf and root transcriptome analysis to identify putative genes involved in triterpenoid saponins biosynthesis in Hedera helix L. PLoS ONE 2017, 12, e0182243. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Wang, L.; Yang, X.; Gong, C.; Zhang, D. Populus endo-β-1,4-glucanases gene family: genomic organization, phylogenetic analysis, expression profiles and association mapping. Planta 2015, 241, 1417–1434. [Google Scholar] [CrossRef] [PubMed]
- Moud, A.M.; Maghsoudi, K. Salt stress effects on respiration and growth of germinated seeds of different wheat (Triticum aestivum L.) cultivars. World J. Agric. Sci. 2008, 4, 351–358. [Google Scholar]
- Moradi, F.; Ismail, A.M. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Liu, H.; Zhao, L.; Wang, X. Effects of exogenous hydrogen sulfide on the redox states of ascorbate and glutathione in maize leaves under salt stress. Biol. Plantarum 2014, 58, 169–173. [Google Scholar] [CrossRef]
- Ramanjulu, S.; Bartels, D. Drought-and desiccation-induced modulation of gene expression in plants. Plant Cell Environ. 2002, 25, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Bao, A.K.; Du, B.Q.; Touil, L.; Kang, P.; Wang, Q.L.; Wang, S.M. Co-expression of tonoplast Cation/H+ antiporter and H+-pyrophosphatase from xerophyte Zygophyllum xanthoxylum improves alfalfa plant growth under salinity, drought and field conditions. Plant Biotechnol. J. 2016, 14, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview. In Plant Responses to Drought Stress; Springer: New York, NY, USA, 2012; pp. 1–33. [Google Scholar] [CrossRef]
- Diaz-Vivancos, P.; Barba-Espín, G.; Hernández, J.A. Elucidating hormonal/ROS networks during seed germination: insights and perspectives. Plant Cell Rep. 2013, 32, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.B.; Luo, J.; Yao, D.W.; Zhu, J.J.; Xu, H.F.; Shi, H.P.; Loor, J.J. Peroxisome proliferator-activated receptor-γ stimulates the synthesis of monounsaturated fatty acids in dairy goat mammary epithelial cells via the control of stearoyl-coenzyme A desaturase. J. Dairy Sci. 2013, 96, 7844–7853. [Google Scholar] [CrossRef] [PubMed]
- Hajiboland, R. Reactive oxygen species and photosynthesis. In Oxidative Damage to Plants; Elsevier: Amsterdam, Netherlands, 2014; pp. 1–63. [Google Scholar] [CrossRef]
- Bompy, F.; Lequeue, G.; Imbert, D.; Dulormne, M. Increasing fluctuations of soil salinity affect seedling growth performances and physiology in three Neotropical mangrove species. Plant Soil 2014, 380, 399–413. [Google Scholar] [CrossRef]
- Szabados, L.; Savoure, A. Proline: a multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Kasuga, M.; Liu, Q.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nat. Biotechnol. 1999, 17, 287. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Sakuma, Y.; Kasuga, M.; Ito, Y.; Seki, M.; Goda, H.; Shimada, Y.; Yoshida, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Identification of cold-inducible downstream genes of the Arabidopsis DREB1A/CBF3 transcriptional factor using two microarray systems. Plant J. 2004, 38, 982–993. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.G.; Tian, Q.Y.; Zhang, W.H. Nitric oxide synthase-dependent nitric oxide production is associated with salt tolerance in Arabidopsis. Plant Physiol. 2007, 144, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Tang, Y.J.; Ma, Q.B.; Yang, C.Y.; Mu, Y.H.; Suo, H.C.; Luo, L.H.; Nian, H. OsDREB2A, a rice transcription factor, significantly affects salt tolerance in transgenic soybean. PLoS ONE 2013, 8, e83011. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene expression and signal transduction in water-stress response. Plant Physiol. 1997, 115, 327. [Google Scholar] [CrossRef] [PubMed]
- Mondini, L.; Nachit, M.M.; Pagnotta, M.A. Allelic variants in durum wheat (Triticum turgidum L. Szabados var. durum) DREB genes conferring tolerance to abiotic stresses. Mol. Genet. Genom. 2015, 290, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Artlip, T.S.; Wisniewski, M.E.; Norelli, J.L. Field evaluation of apple overexpressing a peach CBF gene confirms its effect on cold hardiness, dormancy, and growth. Environ. Exp. Bot. 2014, 106, 79–86. [Google Scholar] [CrossRef]
- Li, J.; Sima, W.; Ouyang, B.; Wang, T.; Ziaf, K.; Luo, Z.; Liu, L.; Li, H.; Chen, M.; Huang, Y.; et al. Tomato SlDREB gene restricts leaf expansion and internode elongation by downregulating key genes for gibberellin biosynthesis. J. Exp. Bot. 2012, 63, 6407–6420. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Xia, X.L.; Yin, W.L. Expression profiling and functional characterization of a DREB2-type gene from Populus euphratica. Biochem. Biophys. Res. Commun. 2009, 378, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Chan, Z.; Bigelow, P.J.; Loescher, W.; Grumet, R. Comparison of salt stress resistance genes in transgenic Arabidopsis thaliana indicates that extent of transcriptomic change may not predict secondary phenotypic or fitness effects. Plant Biotechnol. J. 2012, 10, 284–300. [Google Scholar] [CrossRef] [PubMed]
- Suo, H.; Ma, Q.; Ye, K.; Yang, C.; Tang, Y.; Hao, J.; Zhang, Z.Y.J.; Nian, H. Overexpression of AtDREB1A causes a severe dwarf phenotype by decreasing endogenous gibberellin levels in soybean [Glycine max (L.) Merr.]. PLoS ONE 2012, 7, e45568. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
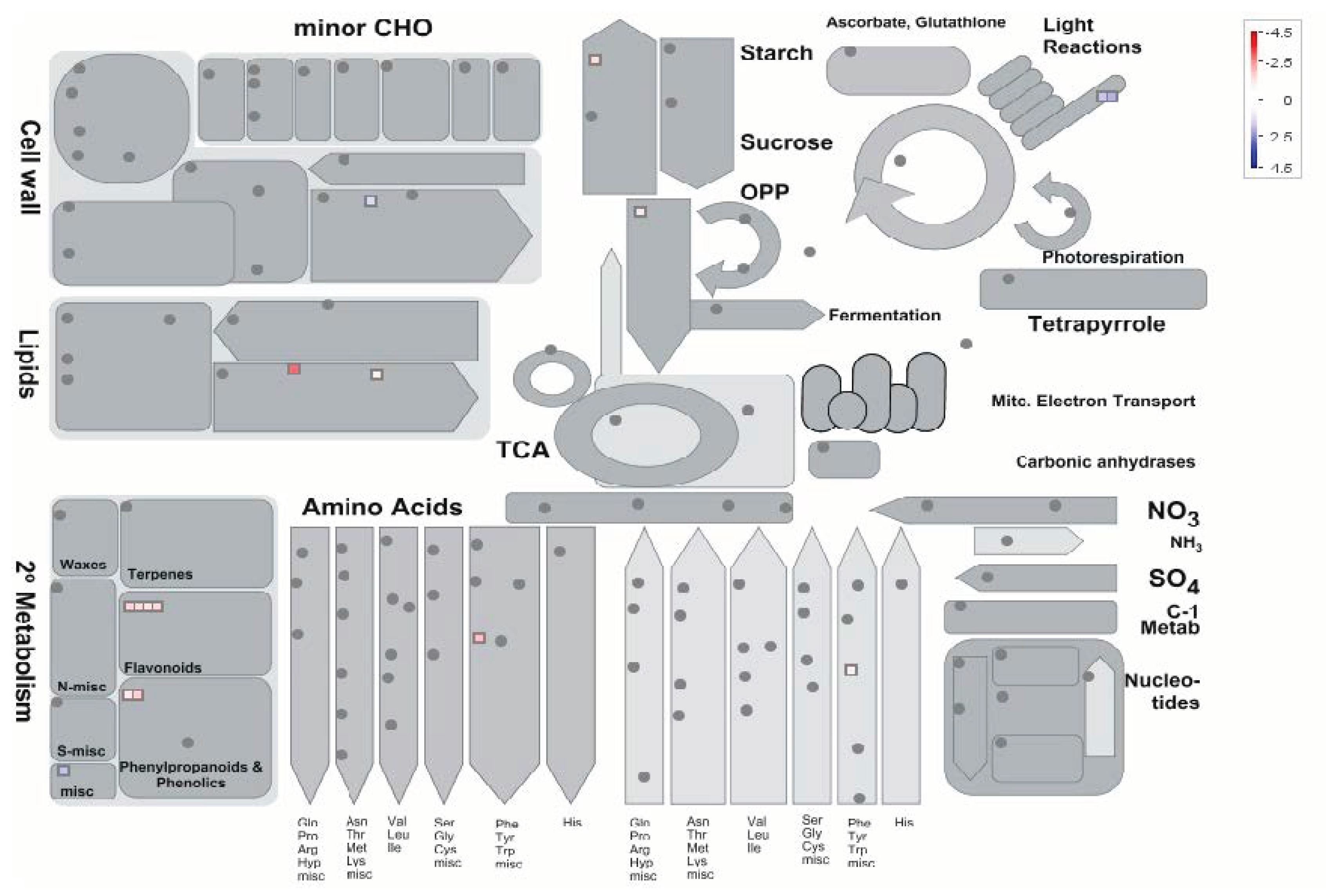{kind=link}
| Gene ID | Function Description | Tair ID | log2FC | p-Value |
|---|---|---|---|---|
| Genes have cri-regulatory element “CCGAC” in promotor regions | ||||
| Potri.002G186600 | WRKY transcription factor | AT4G01720.1 | Inf | 2.42 × 10−4 |
| Potri.003G182200 | WRKY transcription factor | AT1G80840.1 | 1.99 | 1.25 × 10−3 |
| Potri.004G002800 | Phosphotyrosine protein phosphatases superfamily protein | AT1G05000.1 | Inf | 1.27 × 10−3 |
| Potri.011G021100 | Phosphotyrosine protein phosphatases superfamily protein | AT4G03960.1 | 1.22 | 1.54 × 10−3 |
| Potri.013G083600 | Peroxidase superfamily protein | AT5G05340.1 | 6.23 | 3.52 × 10−45 |
| Potri.005G135300 | Peroxidase superfamily protein | AT1G49570.1 | Inf | 1.87 × 10−4 |
| Potri.001G107800 | osmotin 34 | AT4G11650.1 | 3.81 | 1.89 × 10−9 |
| Potri.018G003800 | NAC transcription factor | AT2G24430.2 | 4.27 | 8.08 × 10−5 |
| Potri.005G164900 | myb transcription factor | AT1G34670.1 | 4.04 | 4.48 × 10−4 |
| Potri.008G073700 | laccase 5 | AT2G40370.1 | 4.95 | 1.75 × 10−7 |
| Potri.006G049200 | heat shock transcription factor B3 | AT2G41690.1 | 4.63 | 1.55 × 10−13 |
| Potri.002G015100 | glutathione S-transferase F11 | AT3G03190.1 | 1.38 | 2.08 × 10−5 |
| Potri.001G105200 | glutathione peroxidase 6 | AT4G11600.1 | 1.07 | 2.23 × 10−3 |
| Potri.007G126600 | glutathione peroxidase 2 | AT2G31570.1 | 1.74 | 3.18 × 10−3 |
| Potri.005G020900 | Drought-responsive family protein | AT3G05700.1 | 1.17 | 4.86 × 10−5 |
| Potri.003G134700 | Disease resistance-responsive family protein | AT1G64160.1 | 2.00 | 5.31 × 10−14 |
| Potri.T047500 | disease resistance protein | AT5G17680.1 | 3.86 | 1.41 × 10−45 |
| Potri.013G037300 | Disease resistance protein family | AT5G36930.1 | 1.16 | 3.64 × 10−3 |
| Potri.006G044600 | dehydration-induced protein (ERD15) | AT2G41430.5 | 1.40 | 1.86 × 10−3 |
| Potri.007G072100 | cytochrome P450, family 86, subfamily B, polypeptide 1 | AT5G23190.1 | Inf | 4.64 × 10−3 |
| Potri.004G106600 | cytochrome P450, family 82, subfamily G, polypeptide 1 | AT3G25180.1 | 3.18 | 1.68 × 10−24 |
| Potri.001G334700 | cytochrome P450, family 82, subfamily C, polypeptide 4 | AT4G31940.1 | 1.27 | 6.94 × 10−4 |
| Potri.003G146800 | cytochrome P450, family 78, subfamily A, polypeptide 6 | AT2G46660.1 | 1.50 | 1.31 × 10−4 |
| Potri.003G146800 | cytochrome P450, family 78, subfamily A, polypeptide 6 | AT2G46660.1 | 1.50 | 1.31 × 10−4 |
| Potri.014G072300 | cytochrome P450, family 704, subfamily A, polypeptide 2 | AT2G45510.1 | 1.07 | 1.41 × 10−4 |
| Potri.001G167800 | Cytochrome P450 superfamily protein | AT5G07990.1 | 2.30 | 2.47 × 10−5 |
| Potri.014G019200 | cytochrome B5 isoform D | AT5G48810.1 | 4.41 | 2.60 × 10−5 |
| Potri.017G054300 | cytochrome B5 isoform B | AT2G32720.1 | 2.36 | 1.71 × 10−6 |
| Potri.004G149100 | cold-regulated 413-plasma membrane 2 | AT3G50830.1 | 2.07 | 7.77 × 10−7 |
| Potri.009G116400 | cold, circadian rhythm, and RNA binding 1 | AT4G39260.3 | 1.95 | 2.25 × 10−9 |
| Potri.019G089000 | basic helix-loop-helix (bHLH) DNA-binding superfamily protein | AT1G68810.1 | 2.59 | 9.22 × 10−10 |
| Potri.009G081400 | basic helix-loop-helix (bHLH) DNA-binding superfamily protein | AT4G37850.1 | Inf | 8.35 × 10−6 |
| Potri.009G092900 | Auxin-responsive GH3 family protein | AT2G14960.1 | 5.12 | 2.01 × 10−8 |
| Potri.012G006300 | 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | AT5G24530.1 | 2.19 | 2.62 × 10−13 |
| Potri.005G182700 | 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | AT1G77330.1 | 5.57 | 3.93 × 10−10 |
| Genes have no cri-regulatory element “CCGAC” in promotor regions | ||||
| Potri.019G123500 | WRKY DNA-binding protein 26 | AT2G30490.1 | 1.35 | 5.01 × 10−4 |
| Potri.005G179200 | thylakoidal ascorbate peroxidase | AT1G77490.1 | 1.92 | 4.0 × 10−4 |
| Potri.012G002500 | pyruvate kinase family protein | AT3G49160.1 | 3.763 | 2.50 × 10−4 |
| Potri.007G021300 | S-adenosyl-L-methionine-dependent methyltransferases superfamily protein | AT3G11480.1 | 4.36 | 6.45 × 10−48 |
| Potri.006G222200 | S-adenosyl-L-methionine-dependent methyltransferases superfamily protein | AT5G19530.1 | 1.78 | 8.77 × 10−8 |
| Potri.001G404600 | Peroxisomal membrane 22 kDa family protein | AT4G21380.1 | 1.47 | 5.56 × 10−8 |
| Potri.008G106400 | Peroxidase superfamily protein | AT2G39470.1 | 3.17 | 1.12 × 10−23 |
| Potri.005G195700 | Peroxidase superfamily protein | AT3G11430.1 | 3.64 | 5.63 × 10−31 |
| Potri.006G221800 | myb transcription factor 4 | AT4G00430.1 | 1.07 | 1.40 × 10−3 |
| Potri.002G173900 | myb transcription factor 3 | AT2G43460.1 | 3.60 | 2.47 × 10−24 |
| Potri.005G234500 | salt tolerance homolog2 | AT1G75540.1 | 2.08 | 3.02 × 10−9 |
| Potri.009G037300 | highly ABA-induced PP2C gene 2 | AT2G37170.1 | 4.04 | 9.20 × 10−9 |
| Potri.004G235400 | cytochrome P450, family 707, subfamily A, polypeptide 1 | AT4G18550.1 | 3.07 | 3.43 × 10−7 |
| Potri.003G066400 | Cytochrome P450 superfamily protein | AT1G61720.1 | 3.01 | 4.89 × 10−21 |
| Potri.001G167900 | Cytochrome P450 superfamily protein | AT2G33510.1 | 2.26 | 2.81 × 10−10 |
| Potri.005G113400 | basic helix-loop-helix (bHLH) DNA-binding superfamily protein | AT1G06550.1 | 2.76 | 1.43 × 10−5 |
| Potri.009G107600 | 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | AT5G13930.1 | 1.95 | 1.06 × 10−8 |
| Gene ID | Function description | Tair ID | log2FC | p-Value |
|---|---|---|---|---|
| Potri.005G064300 | ABC-2 type transporter family protein | AT2G13610.1 | 1.60 | 0.00020675 |
| Potri.014G081000 | acetyl Co-enzyme a carboxylase carboxyltransferase alpha subunit | AT2G38040.2 | 1.83 | 5.05 × 10−10 |
| Potri.005G229700 | ADPGLC-PPase large subunit | AT1G27680.1 | −1.38 | 6.79 × 10−9 |
| Potri.001G173700 | alpha/beta-Hydrolases superfamily protein | AT3G15650.2 | −2.80 | 6.58 × 10−8 |
| Potri.004G188100 | arogenate dehydratase 6 | AT1G08250.1 | −1.78 | 1.45 × 10−16 |
| Potri.002G057700 | ATP-dependent caseinolytic (Clp) protease/crotonase family protein | AT1G06550.1 | −1.02 | 1.36 × 10−6 |
| Potri.019G089000 | basic helix-loop-helix (bHLH) DNA-binding superfamily protein | AT1G68810.1 | 3.53 | 2.90 × 10−12 |
| Potri.003G167500 | BCL-2-associated athanogene 5 | AT1G12060.1 | −1.67 | 8.68 × 10−5 |
| Potri.007G055100 | BTB and TAZ domain protein 4 | AT5G67480.2 | −1.53 | 4.51 × 10−10 |
| Potri.008G103900 | Ca2+-binding protein 1 | AT5G49480.1 | −1.38 | 2.76 × 10−5 |
| Potri.013G112500 | calcium-dependent protein kinase 2 | AT1G35670.1 | 1.88 | 4.92 × 10−10 |
| Potri.008G160200 | CBL-interacting protein kinase 4 | AT4G14580.1 | −1.29 | 0.00029303 |
| Potri.001G066400 | CCR-like | AT3G26740.1 | 2.56 | 1.20 × 10−22 |
| Potri.010G113400 | Chaperone DnaJ-domain superfamily protein | AT1G71000.1 | −1.49 | 1.49 × 10−9 |
| Potri.010G084300 | CVP2 like 1 | AT2G32010.2 | −2.15 | 4.23 × 10−13 |
| Potri.001G351400 | cyclophilin 38 | AT3G01480.1 | 1.13 | 5.07 × 10−5 |
| Potri.005G229500 | dihydroflavonol 4-reductase | AT5G42800.1 | −1.38 | 2.02 × 10−10 |
| Potri.005G113700 | flavanone 3-hydroxylase | AT3G51240.1 | −1.10 | 1.53 × 10−6 |
| Potri.012G106500 | Glycosyl hydrolase family 38 protein | AT5G13980.2 | 1.06 | 3.09 × 10−5 |
| Potri.016G138600 | Glycosyl hydrolase superfamily protein | AT5G01930.1 | 1.42 | 0.00014768 |
| Potri.008G151700 | Haloacid dehalogenase-like hydrolase (HAD) superfamily protein | AT2G32150.1 | −1.24 | 1.56 × 10−7 |
| Potri.004G073600 | heat shock protein 90.1 | AT5G52640.1 | −1.18 | 2.27 × 10−7 |
| Potri.008G165200 | HR-like lesion-inducing protein-related | AT4G14420.1 | −1.33 | 6.27 × 10−7 |
| Potri.009G070800 | Lateral root primordium (LRP) protein-related | AT5G12330.4 | 1.27 | 0.00040463 |
| Potri.012G088100 | Leucine-rich receptor-like protein kinase family protein | AT5G56040.2 | 2.70 | 0.00026452 |
| Potri.001G113100 | leucoanthocyanidin dioxygenase | AT4G22880.2 | −1.49 | 1.07 × 10−12 |
| Potri.019G067500 | Major facilitator superfamily protein | AT1G59740.1 | 2.81 | 0.00010361 |
| Potri.002G173900 | myb domain protein 3 | AT3G13540.1 | −1.19 | 2.27 × 10−7 |
| Potri.006G178700 | NAD(P)-binding Rossmann-fold superfamily protein | AT2G23910.1 | −1.11 | 3.45 × 10−5 |
| Potri.008G116500 | NAD(P)-binding Rossmann-fold superfamily protein | AT1G75290.1 | −1.40 | 1.31 × 10−10 |
| Potri.002G234000 | NAD(P)-linked oxidoreductase superfamily protein | AT1G60690.1 | 2.08 | 7.28 × 10−5 |
| Potri.019G093400 | nine-cis-epoxycarotenoid dioxygenase 4 | AT4G19170.1 | 1.44 | 1.14 × 10−10 |
| Potri.008G186500 | Octicosapeptide/Phox/Bem1p family protein | AT3G26510.4 | 1.27 | 8.82 × 10−8 |
| Potri.001G404600 | Peroxisomal membrane 22 kDa (Mpv17/PMP22) family protein | AT1G52870.2 | 2.27 | 6.27 × 10−19 |
| Potri.016G091100 | PHE ammonia lyase 1 | AT2G37040.1 | −1.60 | 6.90 × 10−15 |
| Potri.008G186600 | phosphate transporter 2;1 | AT3G26570.2 | 2.17 | 7.95 × 10−8 |
| Potri.011G142300 | photosystem II subunit R | AT1G79040.1 | 1.70 | 6.07 × 10−10 |
| Potri.006G011200 | Protein of unknown function (DUF594) | AT5G45460.1 | 2.68 | 6.81 × 10−10 |
| Potri.010G210000 | PsbP-like protein 2 | AT2G39470.1 | 2.01 | 1.41 × 10−12 |
| Potri.001G001600 | Pyruvate kinase family protein | AT5G56350.1 | −1.15 | 1.15 × 10−18 |
| Potri.001G098300 | respiratory burst oxidase protein F | AT1G64060.1 | −1.19 | 5.42 × 10−5 |
| Potri.001G055300 | Rubber elongation factor protein (REF) | AT1G67360.2 | −1.07 | 3.27 × 10−19 |
| Potri.017G134900 | transmembrane kinase 1 | AT1G66150.1 | 3.77 | 7.90 × 10−11 |
| Potri.002G195800 | TRICHOME BIREFRINGENCE-LIKE 6 | AT3G62390.1 | −1.81 | 3.95 × 10−6 |
| Potri.006G095000 | tubulin beta 8 | AT5G23860.2 | −1.86 | 2.57 × 10−10 |
| Potri.013G118700 | UDP-glucosyl transferase 78D2 | AT5G17050.1 | −1.73 | 1.81 × 10−10 |
| Potri.014G039000 | Uncharacterised protein family (UPF0114) | AT4G19390.1 | −2.55 | 1.71 × 10−1 |
| Potri.005G064300 | ABC-2 type transporter family protein | AT2G13610.1 | 1.60 | 4.13 × 10−11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Q.; Lu, N.; Sun, Y.; Lv, W.; Luo, Z.; Zhang, H.; Ji, Q.; Yang, Q.; Chen, S.; Zhang, W.; et al. Heterologous Expression of the DREB Transcription Factor AhDREB in Populus tomentosa Carrière Confers Tolerance to Salt without Growth Reduction under Greenhouse Conditions. Forests 2019, 10, 214. https://doi.org/10.3390/f10030214
Guo Q, Lu N, Sun Y, Lv W, Luo Z, Zhang H, Ji Q, Yang Q, Chen S, Zhang W, et al. Heterologous Expression of the DREB Transcription Factor AhDREB in Populus tomentosa Carrière Confers Tolerance to Salt without Growth Reduction under Greenhouse Conditions. Forests. 2019; 10(3):214. https://doi.org/10.3390/f10030214
Chicago/Turabian StyleGuo, Qi, Nan Lu, Yuhan Sun, Wei Lv, Zijing Luo, Huaxin Zhang, Qingju Ji, Qingshan Yang, Shouyi Chen, Wanke Zhang, and et al. 2019. "Heterologous Expression of the DREB Transcription Factor AhDREB in Populus tomentosa Carrière Confers Tolerance to Salt without Growth Reduction under Greenhouse Conditions" Forests 10, no. 3: 214. https://doi.org/10.3390/f10030214