Muscle Progenitors Derived from Extraocular Muscles Express Higher Levels of Neurotrophins and their Receptors than other Cranial and Limb Muscles

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Harvesting
2.2. Isolation and Culture of Myogenic Progenitors
2.3. Density of Mononuclear Cells
2.4. RNA Extraction, cDNA Synthesis and Real-Time PCR for the Detection of Neurotrophins
2.5. Immunohistochemistry and Immunofluorescence
2.6. Confocal Microscopy and Image Analysis
2.7. Statistics
3. Results
3.1. Extraocular and Buccinator Muscles Show Higher Mononuclear Cell Densities when Compared with EDL and Tongue Muscles
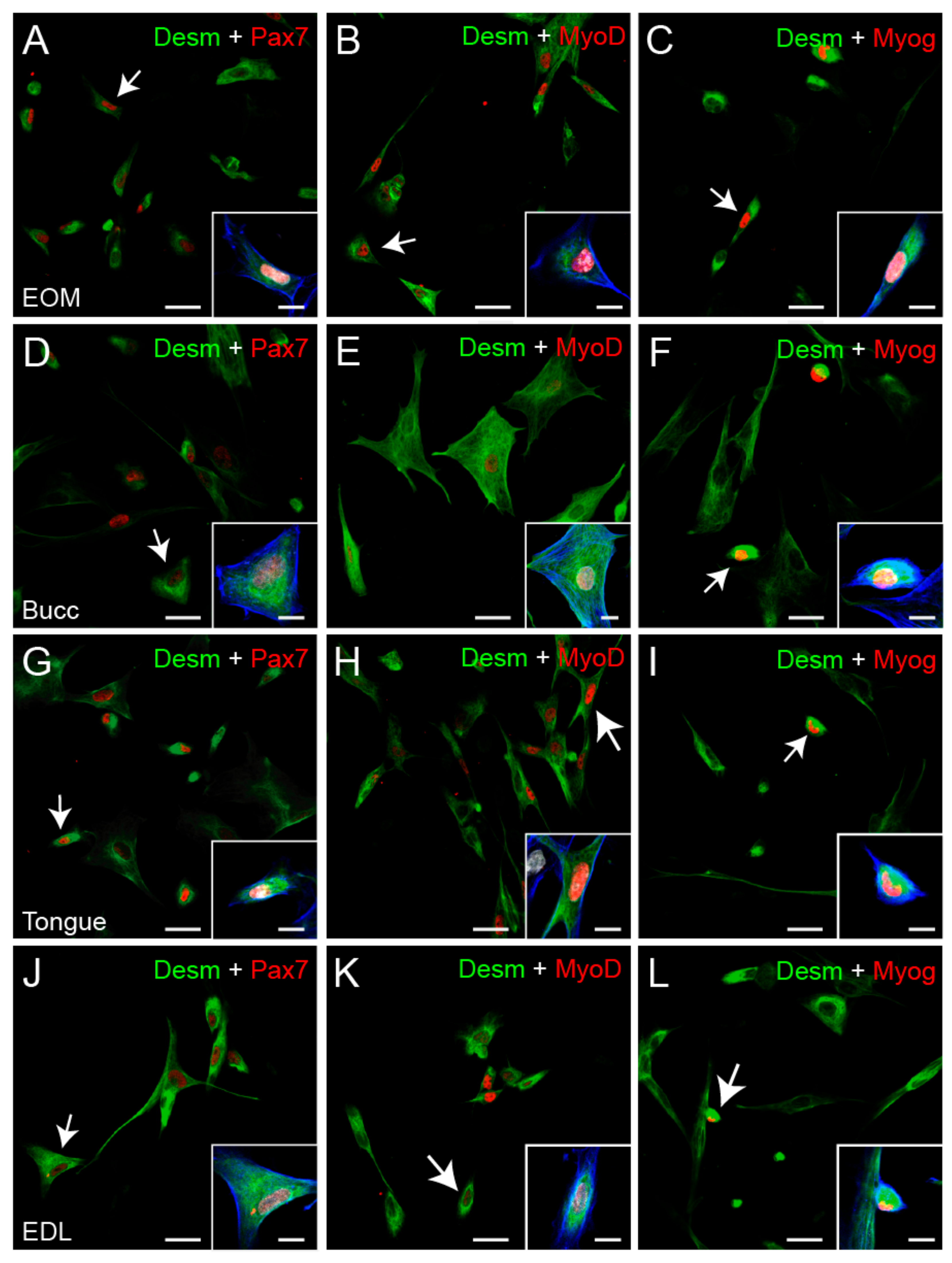
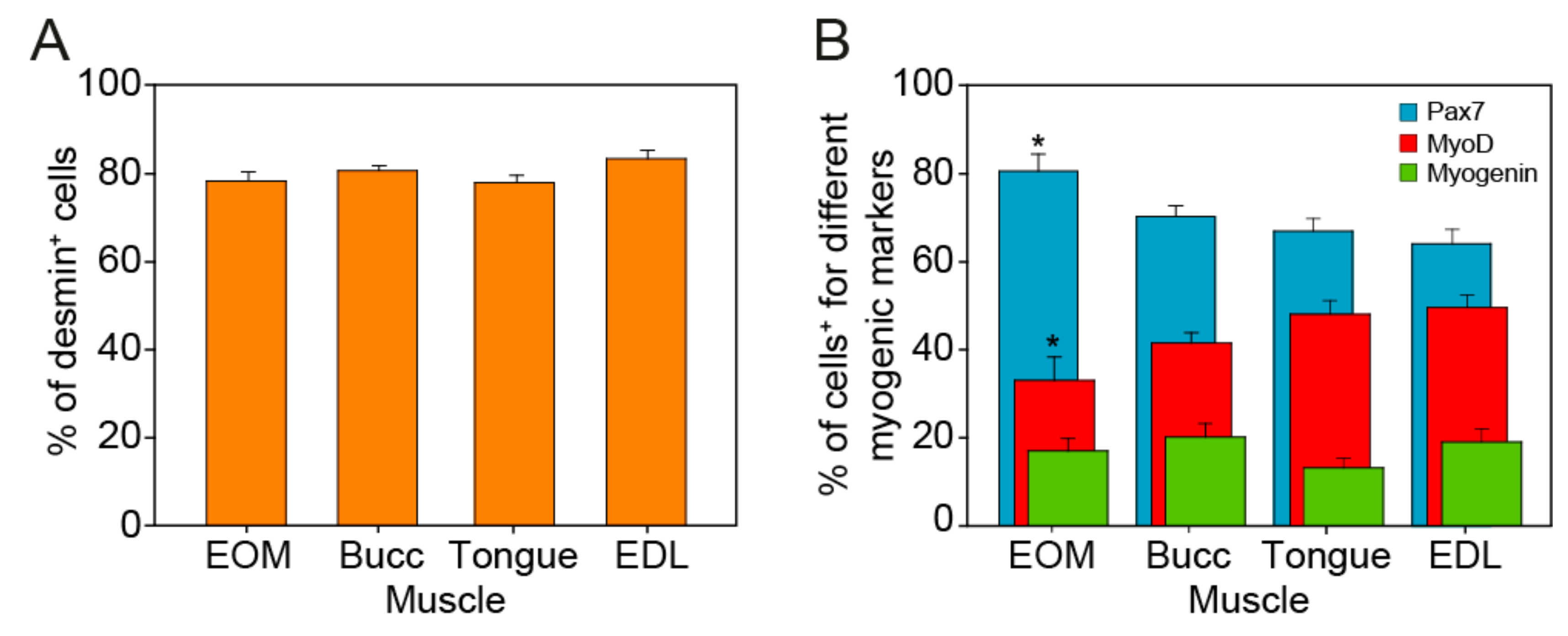
3.2. Myogenicity of the Primary Cultures and State of Differentiation
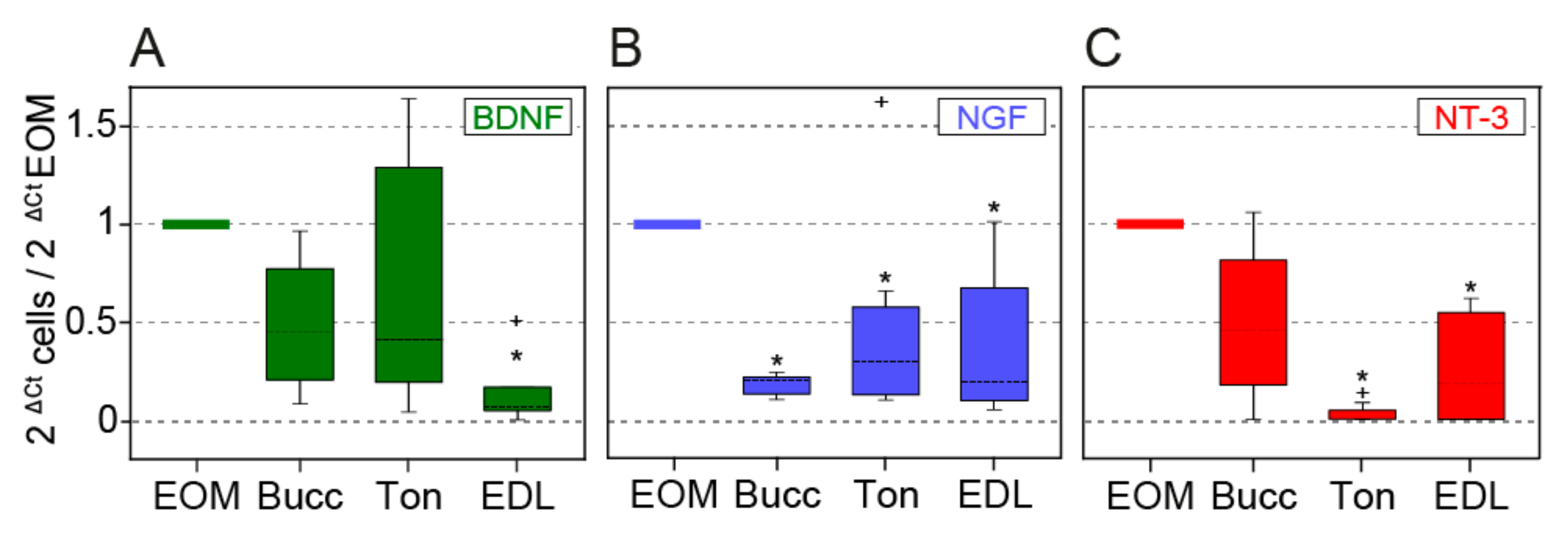
3.3. Myogenic Progenitors Derived from the EOMs Express More Neurotrophins
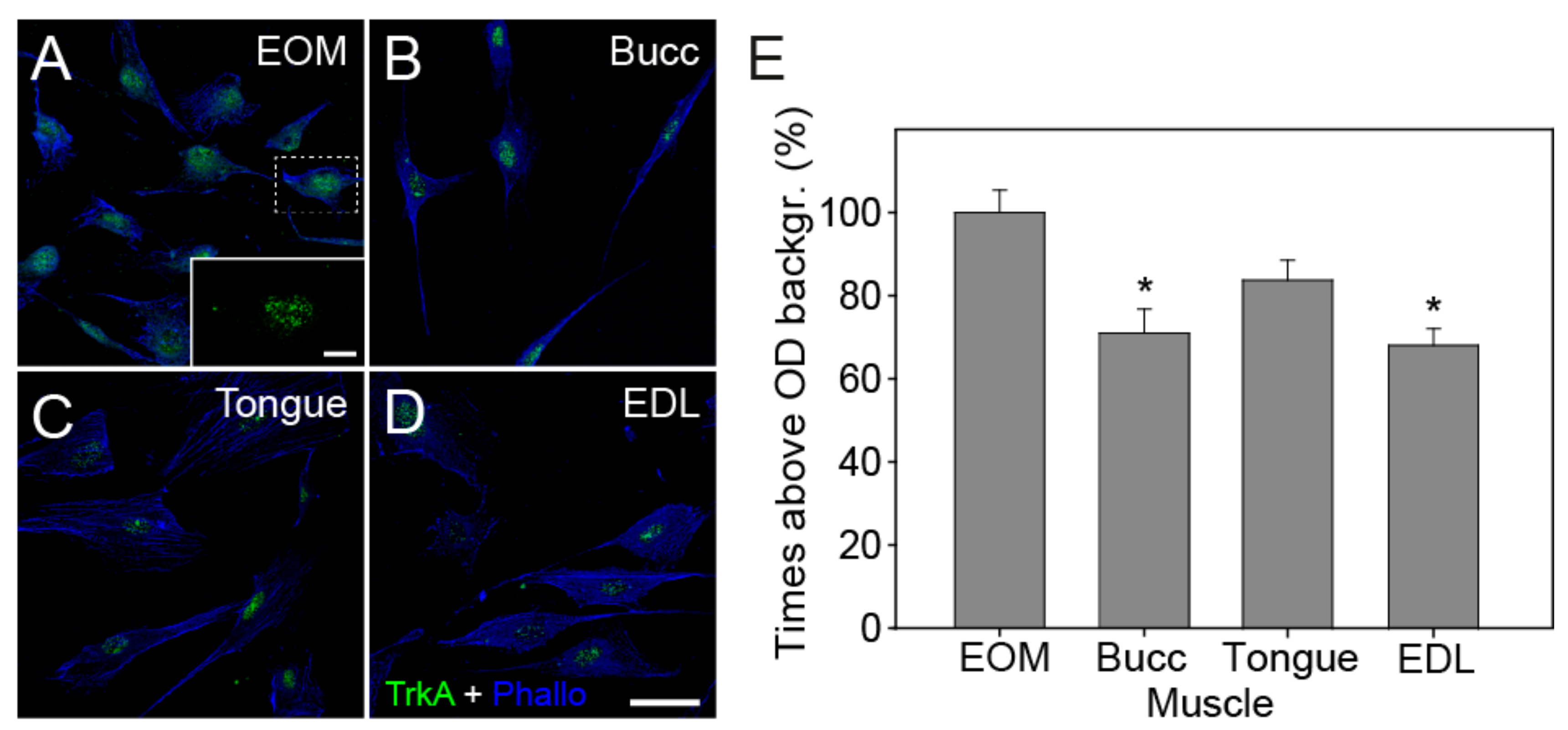
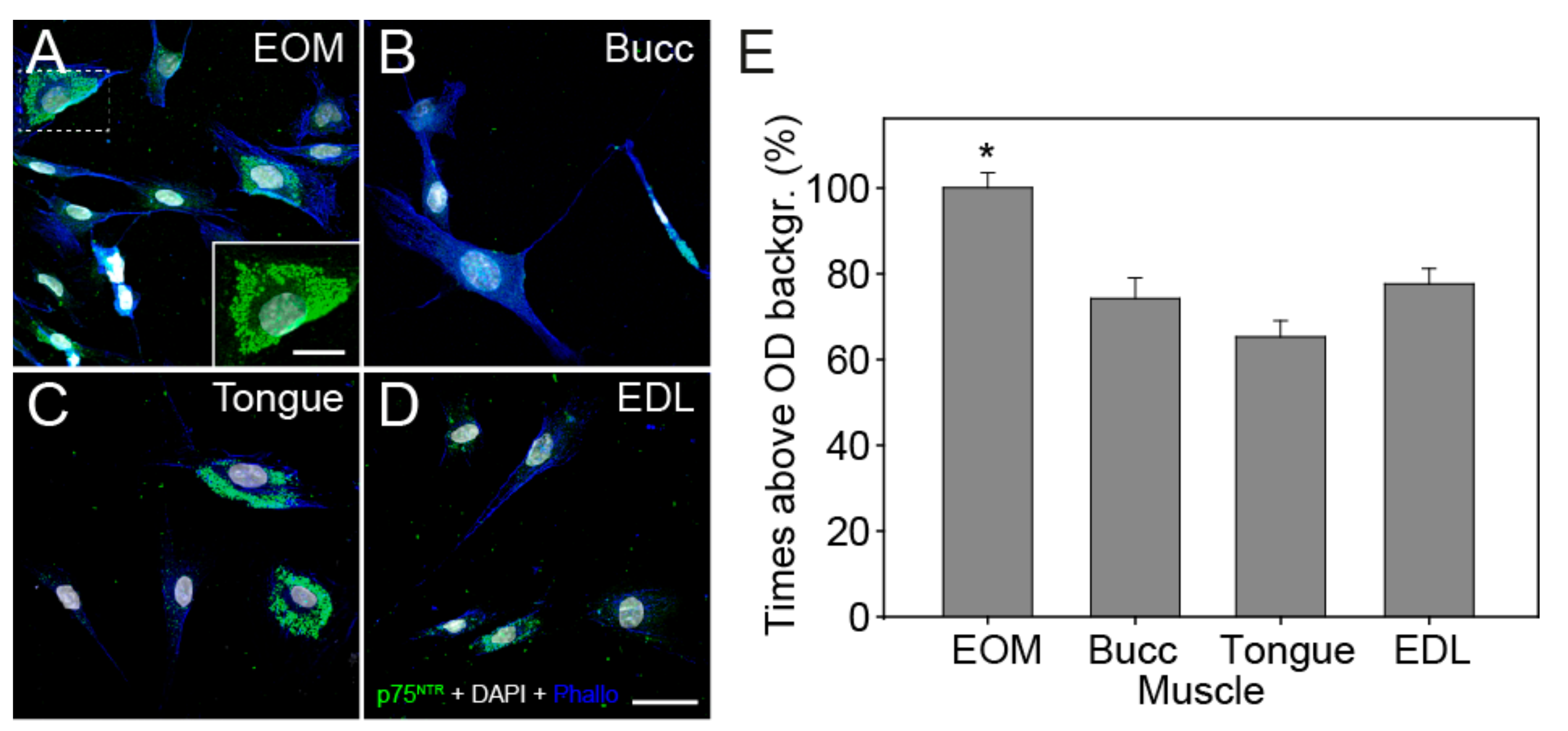
3.4. Expression of Trk and p75NTR Receptors Shown by Immunofluorescence
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Fischer, M.D.; Budak, M.T.; Bakay, M.; Gorospe, J.R.; Kjellgren, D. Definition of the unique human extraocular muscle allotype by expression profiling. Physiol. Genomics 2005, 22, 283–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noden, D.M.; Francis-West, P. The differentiation and morphogenesis of craniofacial muscles. Dev. Dyn. 2006, 235, 1194–1218. [Google Scholar] [CrossRef] [PubMed]
- Büttner-Ennever, J.A. Anatomy of the oculomotor system. Dev. Ophthalmol. 2007, 40, 1–14. [Google Scholar] [PubMed]
- Haslwanter, T. Mechanics of eye movements: implications of the “orbital revolution”. Ann. N. Y. Acad. Sci. 2002, 956, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.D.; Khanna, S.; Kaminski, H.J.; Rao, J.S.; Merriam, A.P. Extraocular muscle is defined by a fundamentally distinct gene expression profile. Proc. Natl. Acad. Sci. 2001, 98, 12062–12067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdez, G.; Tapia, J.C.; Lichtman, J.W.; Fox, M.A.; Sanes, J.R. Shared resistance to aging and ALS in neuromuscular junctions of specific muscles. PloS ONE 2012, 7, e34640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, T.S.; Prendergast, R.A.; Alameddine, H.S.; Tome, F.M.; Fardeau, M. Absence of extraocular muscle pathology in Duchenne’s muscular dystrophy: role for calcium homeostasis in extraocular muscle sparing. J. Exp. Med. 1995, 182, 467–475. [Google Scholar] [CrossRef] [Green Version]
- Ragusa, R.J.; Chow, C.K.; Porter, J.D. Oxidative stress as a potential pathogenic mechanism in an animal model of Duchenne muscular dystrophy. Neuromuscul. Disord. 1997, 7, 379–386. [Google Scholar] [CrossRef]
- Zeiger, U.; Mitchell, C.H.; Khurana, T.S. Superior calcium homeostasis of extraocular muscles. Exp. Eye Res. 2010, 91, 613–622. [Google Scholar] [CrossRef] [Green Version]
- Porter, J.D. Commentary: extraocular muscle sparing in muscular dystrophy: a critical evaluation of potential protective mechanisms. Neuromuscul. Disord. 1998, 8, 198–203. [Google Scholar] [CrossRef]
- Kallestad, K.M.; Hebert, S.L.; McDonald, A.A.; Daniel, M.L.; Cu, S.R.; McLoon, L.K. Sparing of the extraocular muscle in aging and muscular dystrophies: a myogenic precursor cell hypothesis. Exp. Cell Res. 2011, 317, 873–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, A.A.; Kunz, M.D.; McLoon, L.K. Dystrophic changes in extraocular muscles after gamma irradiation in mdx:utrophin (+/-) mice. PLoS ONE 2014, 9, e86424. [Google Scholar] [CrossRef] [PubMed]
- Stuelsatz, P.; Shearer, A.; Li, Y.; Muir, L.A.; Ieronimakis, N.; Shen, Q.W.; Kirillova, I.; Yablonka-Reuveni, Z. Extraocular muscle satellite cells are high performance myo-engines retaining efficient regenerative capacity in dystrophin deficiency. Dev. Biol. 2015, 397, 31–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, P.; Sabourin, L.A.; Girgis-Gabardo, A.; Mansouri, A.; Gruss, P.; Rudnicki, M.A. Pax7 is required for the specification of myogenic satellite cells. Cell 2000, 102, 777–786. [Google Scholar] [CrossRef] [Green Version]
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Pallafacchina, G.; François, S.; Regnault, B.; Czarny, B.; Dive, V.; Cumano, A.; Montarras, D.; Buckingham, M. An adult tissue-specific stem cell in its niche: a gene profiling analysis of in vivo quiescent and activated muscle satellite cells. Stem Cell Res. 2010, 4, 77–91. [Google Scholar] [CrossRef]
- Rocheteau, P.; Vinet, M.; Chretien, F. Dormancy and quiescence of skeletal muscle stem cells. Results Probl. Cell Differ. 2015, 56, 215–235. [Google Scholar]
- Keefe, A.C.; Lawson, J.A.; Flygare, S.D.; Fox, Z.D.; Colasanto, M.P.; Mathew, S.J.; Yandell, M.; Kardon, G. Muscle stem cells contribute to myofibers in sedentary adult mice. Nat. Commun. 2015, 6, 7087. [Google Scholar] [CrossRef] [Green Version]
- Pawlikowski, B.; Pulliam, C.; Betta, N.D.; Kardon, G.; Olwin, B. Pervasive satellite cell contribution to uninjured adult muscle fibers. Skelet. Muscle 2015, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- McLoon, L.K.; Wirtschafter, J. Activated satellite cells are present in uninjured extraocular muscles of mature mice. Trans. Am. Ophthalmol. Soc 2002, 100, 119–123, discussion 123–124. [Google Scholar]
- Oppenheim, R.W. Cell death during development of the nervous system. Annu. Rev. Neurosci. 1991, 14, 453–501. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, A.M. The role of neurotrophins in the developing nervous system. J. Neurobiol. 1994, 25, 1334–1348. [Google Scholar] [CrossRef] [PubMed]
- Giehl, K.M. Trophic dependencies of rodent corticospinal neurons. Rev. Neurosci. 2001, 12, 79–94. [Google Scholar] [CrossRef]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and synaptic plasticity, cognitive function, and dysfunction. Handb. Exp. Pharmacol. 2014, 220, 223–250. [Google Scholar]
- Sakuma, K.; Yamaguchi, A. The recent understanding of the neurotrophin’s role in skeletal muscle adaptation. J. Biomed. Biotechnol. 2011. [Google Scholar] [CrossRef] [Green Version]
- Chevrel, G.; Hohlfeld, R.; Sendtner, M. The role of neurotrophins in muscle under physiological and pathological conditions. Muscle Nerve. 2006, 33, 462–476. [Google Scholar] [CrossRef]
- Mousavi, K.; Parry, D.J.; Jasmin, B.J. BDNF rescues myosin heavy chain IIB muscle fibers after neonatal nerve injury. Am. J. Physiol. Cell Physiol. 2004, 287, C22–C29. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, K.; Jasmin, B.J. BDNF is expressed in skeletal muscle satellite cells and inhibits myogenic differentiation. J. Neurosci. 2006, 26, 5739–5749. [Google Scholar] [CrossRef] [Green Version]
- Clow, C.; Jasmin, B.J. Brain-derived neurotrophic factor regulates satellite cell differentiation and skeletal muscle regeneration. Mol. Biol. Cell. 2010, 21, 2182–2190. [Google Scholar] [CrossRef] [Green Version]
- Colombo, E.; Bedogni, F.; Lorenzetti, I.; Landsberger, N.; Previtali, S.C.; Farina, C. Autocrine and immune cell-derived BDNF in human skeletal muscle: implications for myogenesis and tissue regeneration. J. Pathol. 2013, 231, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Lavasani, M.; Lu, A.; Peng, H.; Cummins, J.; Huard, J. Nerve growth factor improves the muscle regeneration capacity of muscle stem cells in dystrophic muscle. Hum. Gene Ther. 2006, 17, 180–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deponti, D.; Buono, R.; Catanzaro, G.; De Palma, C.; Longhi, R.; Meneveri, R.; Bresolin, N.; Bassi, M.T.; Cossu, G.; Clementi, E.; et al. The low-affinity receptor for neurotrophins p75NTR plays a key role for satellite cell function in muscle repair acting via RhoA. Mol. Biol. Cell. 2009, 20, 3620–3627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellani, L.; Salvati, E.; Alema, S.; Falcone, G. Fine regulation of RhoA and Rock is required for skeletal muscle differentiation. J. Biol. Chem. 2006, 281, 15249–15257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charrasse, S.; Comunale, F.; Grumbach, Y.; Poulat, F.; Blangy, A.; Gauthier-Rouviere, C. RhoAGTPase regulates M-cadherin activity and myoblast fusion. Mol. Biol. Cell. 2006, 17, 749–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruberti, F.; Capsoni, S.; Comparini, A.; Di Daniel, E.; Franzot, J.; Gonfloni, S.; Rossi, G.; Berardi, N.; Cattaneo, A. Phenotypic knockout of nerve growth factor in adult transgenic mice reveals severe deficits in basal forebrain cholinergic neurons, cell death in the spleen, and skeletal muscle dystrophy. J. Neurosci. 2000, 20, 2589–2601. [Google Scholar] [CrossRef]
- Capsoni, S.; Ruberti, F.; Di Daniel, E.; Cattaneo, A. Muscular dystrophy in adult and aged anti-NGF transgenic mice resembles an inclusion body myopathy. J. Neurosci. Res. 2000, 59, 553–560. [Google Scholar] [CrossRef]
- Sterne, G.D.; Coulton, G.R.; Brown, R.A.; Green, C.J.; Terenghi, G. Neurotrophin-3-enhanced nerve regeneration selectively improves recovery of muscle fibers expressing myosin heavy chains 2b. J. Cell Biol. 1997, 139, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, K.; Miranda, W.; Parry, D.J. Neurotrophic factors enhance the survival of muscle fibers in EDL, but not SOL, after neonatal nerve injury. Am. J. Physiol. Cell Physiol. 2002, 283, C950–C959. [Google Scholar] [CrossRef] [Green Version]
- Yalvac, M.E.; Amornvit, J.; Chen, L.; Shontz, K.M.; Lewis, S.; Sahenk, Z. AAV1.NT-3 gene therapy increases muscle fiber diameter through activation of mTOR pathway and metabolic remodeling in a CMT mouse model. Gene Ther. 2018, 25, 129–138. [Google Scholar] [CrossRef]
- Stuelsatz, P.; Shearer, A.; Yablonka-Reuveni, Z. Ancestral Myf5 gene activity in periocular connective tissue identifies a subset of fibro/adipogenic progenitors but does not connote a myogenic origin. Dev. Biol. 2014, 385, 366–379. [Google Scholar] [CrossRef] [Green Version]
- Keire, P.; Shearer, A.; Shefer, G.; Yablonka-Reuveni, Z. Isolation and culture of skeletal muscle myofibers as a means to analyze satellite cells. Methods Mol. Biol. 2013, 946, 431–468. [Google Scholar] [PubMed] [Green Version]
- Rando, T.A.; Blau, H.M. Primary mouse myoblast purification, characterization, and transplantation for cell-mediated gene therapy. J. Cell Biol. 1994, 125, 1275–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, M.; Fitzpatrick, K.; McLoon, L.K. Extraocular Muscle Repair and Regeneration. Curr. Ophthalmol. Rep. 2017, 5, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Sirabella, D.; De Angelis, L.; Berghella, L. Sources for skeletal muscle repair: from satellite cells to reprogramming. J. Cachexia Sarcopenia Muscle 2013, 4, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Gharaibeh, B.; Lu, A.; Tebbets, J. Isolation of a slowly adhering cell fraction containing stem cells from murine skeletal muscle by the preplate technique. Nat. Protoc. 2008, 3, 1501–1509. [Google Scholar] [CrossRef]
- Yablonka-Reuveni, Z.; Rivera, A.J. Temporal expression of regulatory and structural muscle proteins during myogenesis of satellite cells on isolated adult rat fibers. Dev. Biol. 1994, 164, 588–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Baquero-Perez, B.; Kuchipudi, S.V.; Nelli, R.K.; Chang, K.C. A simplified but robust method for the isolation of avian and mammalian muscle satellite cells. BMC Cell Biol. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirieleison, S.M.; Feduska, J.M.; Schugar, R.C.; Askew, Y.; Deasy, B.M. Human muscle-derived cell populations isolated by differential adhesion rates: phenotype and contribution to skeletal muscle regeneration in Mdx/SCID mice. Tissue Eng. Part A. 2012, 18, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, S.L.; Daniel, M.L.; McLoon, L.K. The role of Pitx2 in maintaining the phenotype of myogenic precursor cells in the extraocular muscles. PLoS ONE 2013, 8, e58405. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique, 6th ed.; Methuen & Co Ltd.: London, UK, 1959. [Google Scholar]
- Barbacid, M. The Trk family of neurotrophic receptors. J. Neurobiol. 1994, 25, 1386–1403. [Google Scholar] [CrossRef]
- Seidl, K.; Erck, C.; Buchberger, A. Evidence for participation of nerve growth factor and its low-affinity receptor (p75NTR) in the regulation of the myogenic program. J. Cell Physiol. 1998, 176, 10–21. [Google Scholar] [CrossRef]
- Bonacchi, A.; Taddei, M.L.; Petrai, I. Nuclear localization of TRK-A in liver cells. Histol. Histopathol 2008, 23, 327–340. [Google Scholar] [PubMed]
- Rende, M.; Brizi, E.; Conner, J.; Treves, S.; Censier, K.; Provenzano, C.; Taglialatela, G.; Sanna, P.P.; Donato, R. Nerve growth factor (NGF) influences differentiation and proliferation of myogenic cells in vitro via TrKA. Int. J. Dev. Neurosci. 2000, 18, 869–885. [Google Scholar] [CrossRef]
- Wheeler, E.F.; Gong, H.; Grimes, R.; Benoit, D.; Vazquez, L. p75NTR and Trk receptors are expressed in reciprocal patterns in a wide variety of non-neural tissues during rat embryonic development, indicating independent receptor functions. J. Comp. Neurol. 1998, 391, 407–428. [Google Scholar] [CrossRef]
- Reddypalli, S.; Roll, K.; Lee, H.K.; Lundell, M.; Barea-Rodriguez, E.; Wheeler, E.F. p75NTR-mediated signaling promotes the survival of myoblasts and influences muscle strength. J. Cell Physiol. 2005, 204, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, K.; Lecht, S.; Arien-Zakay, H.; Cohen, G.; Aga-Mizrachi, S.; Yanay, N.; Saragovi, H.U.; Nedev, H.; Marcinkiewicz, C.; Nevo, Y.; et al. Nerve growth factor stimulation of ERK1/2 phosphorylation requires both p75NTR and α9β1 integrin and confers myoprotection towards ischemia in C2C12 skeletal muscle cell model. Cell Signal. 2012, 24, 2378–2388. [Google Scholar] [CrossRef]
- de Perini, A.; Dimauro, I.; Duranti, G. The p75NTR-mediated effect of nerve growth factor in L6C5 myogenic cells. BMC Res. Notes 2017, 10, 686. [Google Scholar] [CrossRef] [Green Version]
- Colombo, E.; Romaggi, S.; Medico, E.; Menon, R.; Mora, M.; Falcone, C. Human neurotrophin receptor p75NTR defines differentiation-oriented skeletal muscle precursor cells: implications for muscle regeneration. J. Neuropathol. Exp. Neurol. 2011, 70, 133–142. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter Measured | EOM | N | Buccinator | N | Tongue | N | EDL | N |
|---|---|---|---|---|---|---|---|---|
| Muscle weight (g) | 0.20 ± 0.014●*‡ | 10 | 0.03 ± 0.004●†‡ | 4 | 0.49 ± 0.017*† | 4 | 0.56 ± 0.045*† | 3 |
| N cells before plating (×105) | 2.88 ± 0.44* | 10 | 0.63 ± 0.17 | 4 | 3.30 ± 0.6* | 4 | 3.25 ± 0.19* | 3 |
| N cells on 1st passage (×105) | 2.92 ± 0.51* | 7 | 0.68 ± 0.19 | 4 | 2.95 ± 0.65 | 3 | 3.17 ± 0.08* | 3 |
| N cells (×105)/g muscle | 1.45 ± 0.16* | 10 | 2.21 ± 0.28 | 4 | 0.67 ± 0.12*† | 4 | 0.59 ± 0.04*† | 3 |
| N rats to obtain 1 × 106 cells | 7 | - | 32 | - | 6 | - | 6 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrero-Rojas, G.; Benítez-Temiño, B.; Pastor, A.M.; Davis López de Carrizosa, M.A. Muscle Progenitors Derived from Extraocular Muscles Express Higher Levels of Neurotrophins and their Receptors than other Cranial and Limb Muscles. Cells 2020, 9, 747. https://doi.org/10.3390/cells9030747
Carrero-Rojas G, Benítez-Temiño B, Pastor AM, Davis López de Carrizosa MA. Muscle Progenitors Derived from Extraocular Muscles Express Higher Levels of Neurotrophins and their Receptors than other Cranial and Limb Muscles. Cells. 2020; 9(3):747. https://doi.org/10.3390/cells9030747
Chicago/Turabian StyleCarrero-Rojas, Génova, Beatriz Benítez-Temiño, Angel M. Pastor, and Mª América Davis López de Carrizosa. 2020. "Muscle Progenitors Derived from Extraocular Muscles Express Higher Levels of Neurotrophins and their Receptors than other Cranial and Limb Muscles" Cells 9, no. 3: 747. https://doi.org/10.3390/cells9030747