Purine Metabolites in Tumor-Derived Exosomes May Facilitate Immune Escape of Head and Neck Squamous Cell Carcinoma

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Cancer Genome Atlas (TCGA) Analysis
2.2. Cell Lines
2.3. Patients
2.4. Exosome Isolation by mini-SEC
2.5. Exosome Characterization
2.6. UPLC-MS/MS for Purine Metabolites
2.7. Statistical Analysis
3. Results
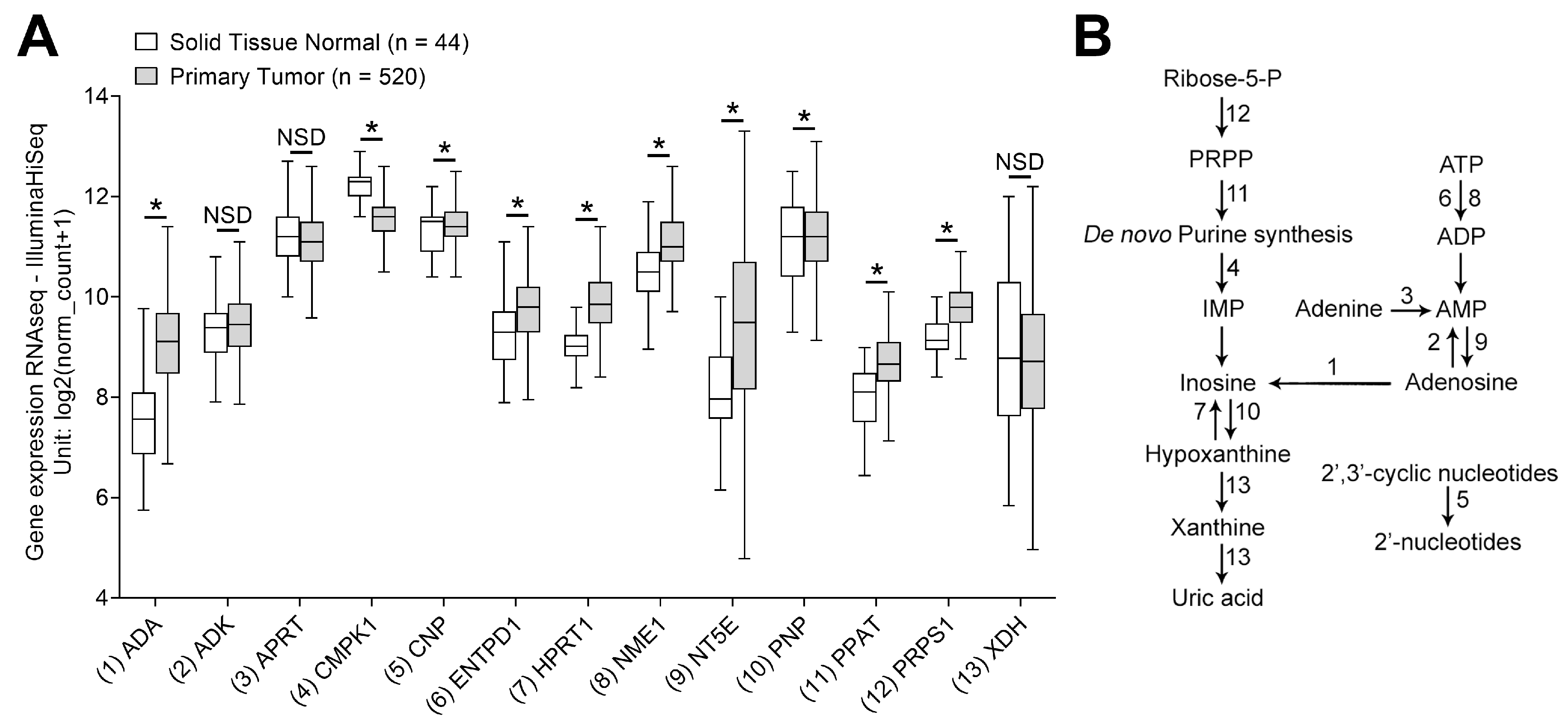
3.1. mRNA Transcripts for Purine Metabolites Are Upregulated in HNSCC
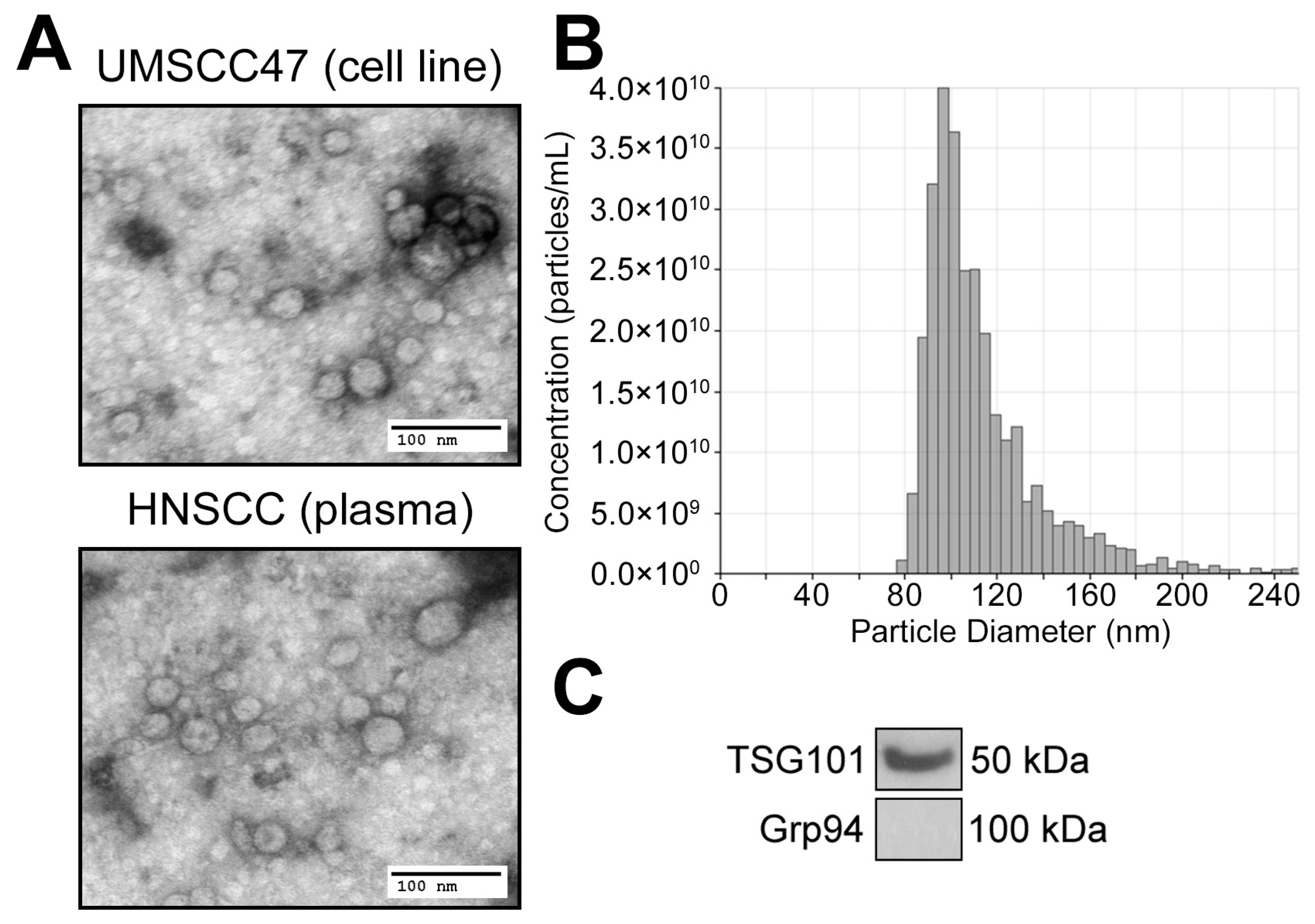
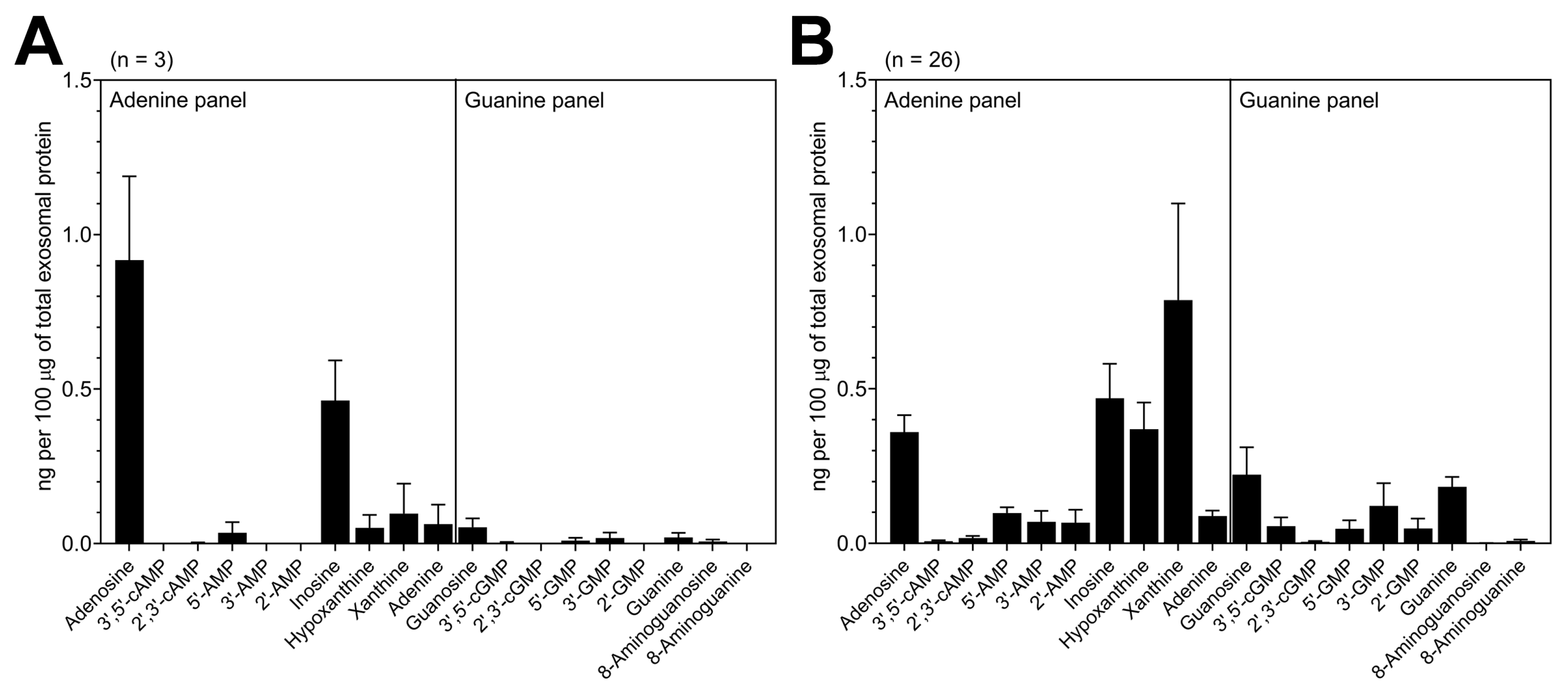
3.2. TEX Produced by HNSCC Cells Encapsulate Purine Metabolites
3.3. Exosomes Derived from HNSCC Patients’ Plasma Contain Purine Metabolites
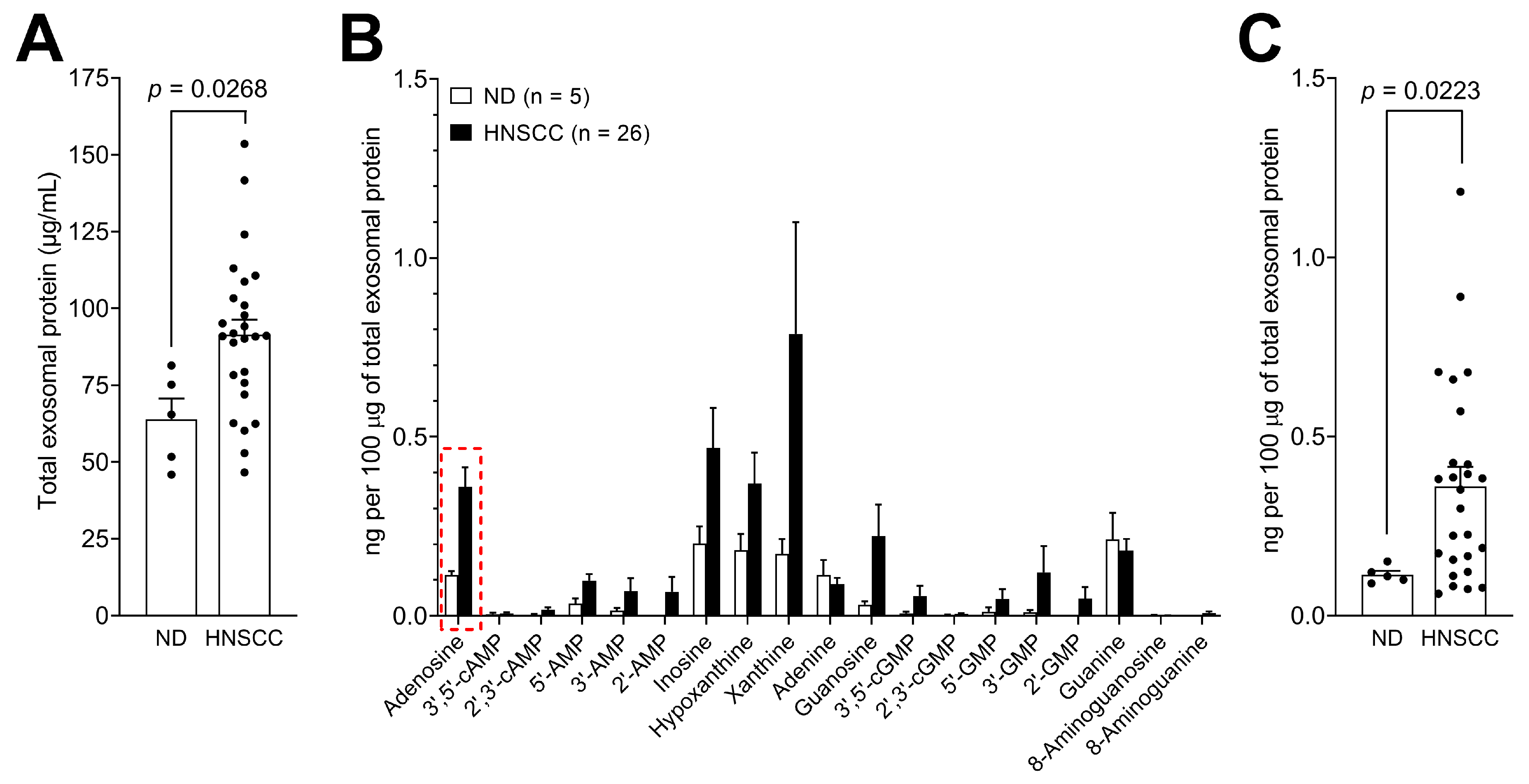
3.4. Levels of Purine Metabolites are Upregulated in Plasma-Derived Exosomes from Cancer Patients Compared to Normal Donors
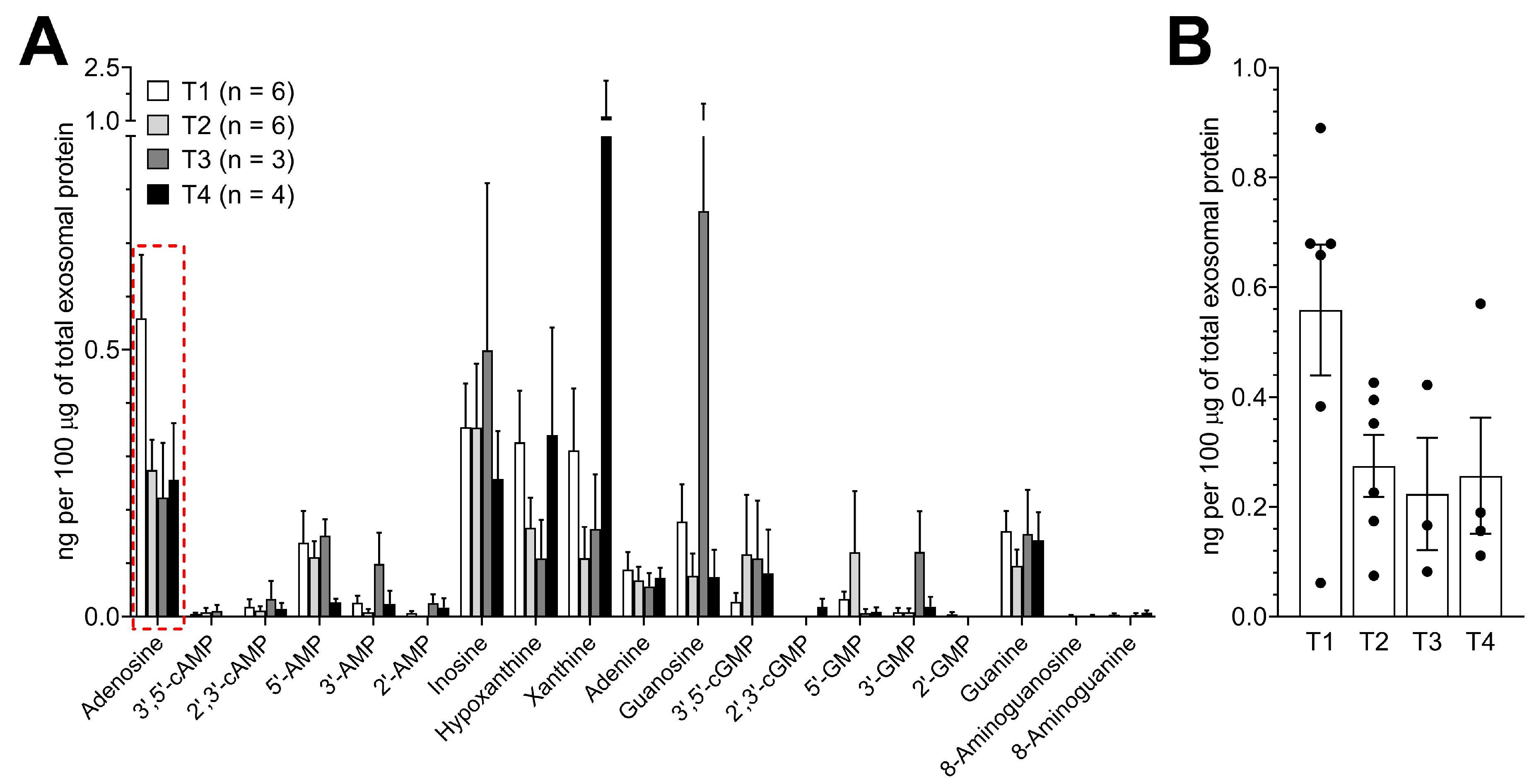
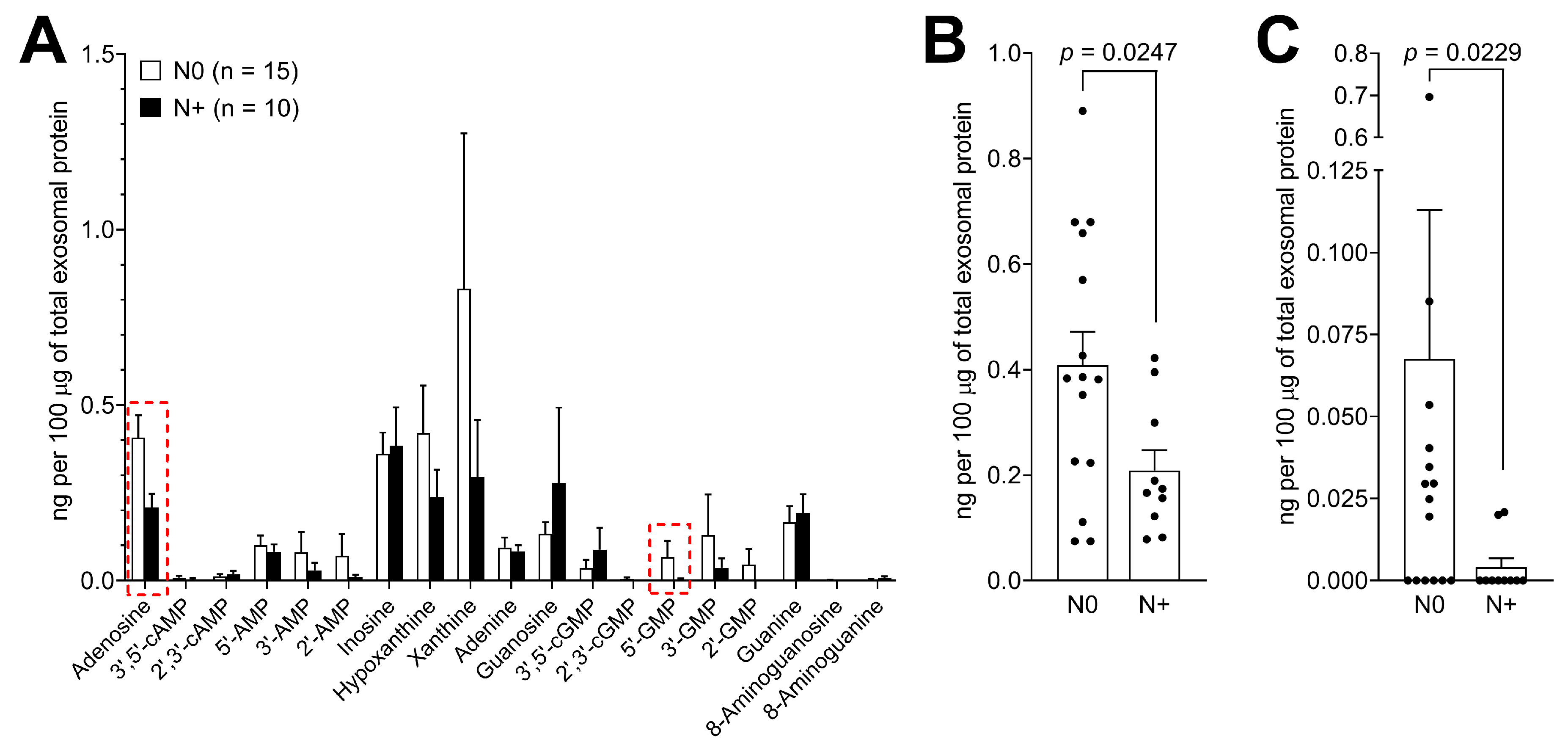
3.5. Purine Metabolite Levels in Plasma-Derived Exosomes are Elevated in Patients with Early-Stage Tumors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Leemans, C.R.; Snijders, P.J.F.; Brakenhoff, R.H. The molecular landscape of head and neck cancer. Nat. Rev. Cancer 2018, 18, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Leemans, C.R.; Braakhuis, B.J.M.; Brakenhoff, R.H. The molecular biology of head and neck cancer. Nat. Rev. Cancer 2011, 11, 9–22. [Google Scholar] [CrossRef]
- Bose, P.; Brockton, N.T.; Dort, J.C. Head and neck cancer: From anatomy to biology. Int. J. Cancer 2013, 133, 2013–2023. [Google Scholar] [CrossRef] [PubMed]
- Mandapathil, M.; Szczepanski, M.J.; Szajnik, M.; Ren, J.; Jackson, E.K.; Johnson, J.T.; Gorelik, E.; Lang, S.; Whiteside, T.L. Adenosine and prostaglandin E2 cooperate in the suppression of immune responses mediated by adaptive regulatory T cells. J. Biol. Chem. 2010, 285, 27571–27580. [Google Scholar] [CrossRef] [Green Version]
- Azambuja, J.H.; Ludwig, N.; Braganhol, E.; Whiteside, T.L. Inhibition of the adenosinergic pathway in cancer rejuvenates innate and adaptive immunity. Int. J. Mol. Sci. 2019, 20, 5698. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Diergaarde, B.; Ferrone, S.; Kirkwood, J.M.; Whiteside, T.L. Melanoma cell-derived exosomes in plasma of melanoma patients suppress functions of immune effector cells. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Whiteside, T.L. Tumor-Derived Exosomes and Their Role in Cancer Progression. Adv. Clin. Chem. 2016, 74, 103–141. [Google Scholar] [PubMed] [Green Version]
- Rak, J. Extracellular vesicles biomarkers and effectors of the cellular interactome in cancer. Front. Pharm. 2013, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Szczepanski, M.J.; Szajnik, M.; Welsh, A.; Whiteside, T.L.; Boyiadzis, M. Blast-derived microvesicles in sera from patients with acute myeloid leukemia suppress natural killer cell function via membrane-associated transforming growth factor-beta1. Haematologica 2011, 96, 1302–1309. [Google Scholar] [CrossRef]
- Skog, J.; Wurdinger, T.; Van Rijn, S.; Meijer, D.; Gainche, L.; Sena-esteves, M.; Curry, W.T., Jr.; Carter, R.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and protein that promote tumor growth and provide diagnostic biomarkers. Nat Cell Biol. 2012, 10, 1470–1476. [Google Scholar] [CrossRef]
- Ludwig, N.; Yerneni, S.; Azambuja, J.; Gillespie, D.; Menshikova, E.; Jackson, E.; Whiteside, T. Tumor-derived exosomes promote angiogenesis via adenosine A2B receptor signaling. Angiogenesis 2020. [Google Scholar] [CrossRef] [PubMed]
- Schuler, P.J.; Saze, Z.; Hong, C.S.; Muller, L.; Gillespie, D.G.; Cheng, D.; Harasymczuk, M.; Mandapathil, M.; Lang, S.; Jackson, E.K.; et al. Human CD4+ CD39+ regulatory T cells produce adenosine upon co-expression of surface CD73 or contact with CD73+ exosomes or CD73+ cells. Clin. Exp. Immunol. 2014, 177, 531–543. [Google Scholar] [CrossRef]
- Clayton, A.; Al-Taei, S.; Webber, J.; Mason, M.D.; Tabi, Z. Cancer Exosomes Express CD39 and CD73, Which Suppress T Cells through Adenosine Production. J. Immunol. 2011, 187, 676–683. [Google Scholar] [CrossRef]
- Sayner, S.; Choi, C.; Maulucci, M.; KC, R.; Zhou, C.; Scruggs, A.; Yarbrough, T.; Blair, L.; King, J.; Seifert, R.; et al. Extracellular vesicles: Another compartment for the second messenger, cyclic adenosine monophosphate (cAMP). Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 316, L691–L700. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M.; Craft, B.; Kamath, A.; Brooks, A.; Zhu, J.; Haussler, D. The UCSC Xena Platform for cancer genomics data visualization and interpretation. BioRxiv 2018, 326470. [Google Scholar]
- Heo, D.S.; Snyderman, C.; Gollin, S.M.; Pan, S.; Walker, E.; Deka, R.; Barnes, E.L.; Johnson, J.T.; Herberman, R.B. Biology, Cytogenetics, and Sensitivity to Immunological Effector Cells of New Head and Neck Squamous Cell Carcinoma Lines. Cancer Res. 1989, 49, 5167–5175. [Google Scholar] [PubMed]
- Ludwig, N.; Razzo, B.M.; Yerneni, S.S.; Whiteside, T.L. Optimization of cell culture conditions for exosome isolation using mini-size exclusion chromatography (mini-SEC). Exp. Cell Res. 2019, 378, 149–157. [Google Scholar] [CrossRef]
- Ludwig, S.; Sharma, P.; Theodoraki, M.-N.; Pietrowska, M.; Yerneni, S.S.; Lang, S.; Ferrone, S.; Whiteside, T.L. Molecular and Functional Profiles of Exosomes From HPV(+) and HPV(−) Head and Neck Cancer Cell Lines. Front. Oncol. 2018, 12, 445. [Google Scholar] [CrossRef]
- Ludwig, S.; Marczak, L.; Sharma, P.; Abramowicz, A.; Gawin, M.; Widlak, P.; Whiteside, T.L.; Pietrowska, M. Proteomes of exosomes from HPV(+) or HPV(-) head and neck cancer cells: Differential enrichment in immunoregulatory proteins. Oncoimmunology 2019, 8, 1–11. [Google Scholar] [CrossRef]
- Ludwig, N.; Hong, C.-S.; Ludwig, S.; Azambuja, J.H.; Sharma, P.; Theodoraki, M.-N.; Whiteside, T.L. Isolation and Analysis of Tumor-Derived Exosomes. Curr. Protoc. Immunol. 2019, 127, e91. [Google Scholar] [CrossRef]
- Ludwig, N.; Yerneni, S.S.; Razzo, B.M.; Whiteside, T.L. Exosomes from HNSCC Promote Angiogenesis through Reprogramming of Endothelial Cells. Mol. Cancer Res. 2018, 16, 1798–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2019, 8, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, N.; Whiteside, T.L. Potential Roles of Tumor-derived Exosomes in Angiogenesis. Expert Opin. Targets 2018, 22, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, T.L. Exosomes and tumor-mediated immune suppression. J. Clin. Invest 2016, 126, 1216–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Lv, T.; Zhang, Q.; Zhu, Q.; Zhan, P.; Zhu, S.; Zhang, J.; Song, Y. The biology, function and clinical implications of exosomes in lung cancer. Cancer Lett. 2017, 407, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Zebrowska, A.; Skowronek, A.; Wojakowska, A.; Widlak, P.; Pietrowska, M. Metabolome of exosomes: Focus on vesicles released by cancer cells and present in human body fluids. Int. J. Mol. Sci. 2019, 20, 3461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puhka, M.; Takatalo, M.; Nordberg, M.E.; Valkonen, S.; Nandania, J.; Aatonen, M.; Yliperttula, M.; Laitinen, S.; Velagapudi, V.; Mirtti, T.; et al. Metabolomic profiling of extracellular vesicles and alternative normalization methods reveal enriched metabolites and strategies to study prostate cancer-related changes. Theranostics 2017, 7, 3824–3841. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, S.C.; Dos Santos Coelho, I.; Lieberknecht, V.; Cunha, M.P.; Calixto, J.B.; Rodrigues, A.L.S.; Santos, A.R.S.; Dutra, R.C. Inosine, an Endogenous Purine Nucleoside, Suppresses Immune Responses and Protects Mice from Experimental Autoimmune Encephalomyelitis: A Role for A2A Adenosine Receptor. Mol. Neurobiol. 2017, 54, 3271–3285. [Google Scholar] [CrossRef] [PubMed]
- Razzo, B.; Ludwig, N.; Hong, C.; Sharma, P.; Fabian, K.; Fecek, R.; Storkus, W.; Whiteside, T. Tumor-derived exosomes promote carcinogenesis of murine oral squamous cell carcinoma. Carcinogenesis 2019, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.-S.; Muller, L.; Whiteside, T.L.; Boyiadzis, M. Plasma exosomes as markers of therapeutic response in patients with acute myeloid leukemia. Front. Immunol. 2014, 5, 160. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, S.; Floros, T.; Theodoraki, M.-N.; Hong, C.-S.; Jackson, E.; Lang, S.; Whiteside, T. Suppression of lymphocyte functions by plasma exosomes correlates with disease activity in patients with head and neck cancer. Clin. Cancer Res. 2017, 23, 4843–4854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, E.K.; Ren, J.; Mi, Z. Extracellular 2′,3′-cAMP is a source of adenosine. J. Biol. Chem. 2009, 284, 33097–33106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, E.K.; Cheng, D.; Mi, Z.; Gillespie, D.G. Guanosine regulates adenosine levels in the kidney. Physiol. Rep. 2014, 2, 1–9. [Google Scholar] [CrossRef]
- Jackson, E.; Mi, Z.; Janesko-Feldman, K.; Jackson, T.; Kochanek, P. 2′,3′-cGMP Exists In Vivo and Comprises a 2′,3′-cGMP-Guanosine Pathway. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 316, R783–R790. [Google Scholar] [CrossRef]
- Schneider, E.H.; Hofmeister, O.; Kälble, S.; Seifert, R. Apoptotic and anti-proliferative effect of guanosine and guanosine derivatives in HuT-78 T lymphoma cells. Naunyn. Schmiedebergs Arch. Pharm. 2020, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, N.; Azambuja, J.H.; Rao, A.; Gillespie, D.G.; Jackson, E.K.; Whiteside, T.L. Adenosine receptors regulate exosome production. Purinergic Signal. 2020. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Internal Standard or Target Purine | Parent Ion (m/z) | Collision Energy (volts) | Daughter Ion (m/z) | Approximate Retention Time (min) |
|---|---|---|---|---|
| Analysis of Adenosine | ||||
| 13C10-Adenosine | 278 | 19 | 141 | 3.29 |
| Adenosine | 268 | 19 | 136 | 3.29 |
| Analysis of 3′,5′-cAMP | ||||
| 13C5-3′,5′-cAMP | 335 | 28 | 136 | 3.8 |
| 3′,5′-cAMP | 330 | 28 | 136 | 3.8 |
| Analysis of 2′,3′-cAMP | ||||
| 13C5-2′,3′-cAMP | 335 | 28 | 136 | 2.42 |
| 2′,3′-cAMP | 330 | 28 | 136 | 2.42 |
| Analysis of 5′-AMP | ||||
| 13C10-5′-AMP | 358 | 19 | 141 | 1.72 |
| 5′-AMP | 348 | 19 | 136 | 1.72 |
| Analysis of 3′-AMP | ||||
| 13C5-3′-AMP | 353 | 19 | 136 | 2.16 |
| 3′-AMP | 348 | 19 | 136 | 2.16 |
| Analysis of 2′-AMP | ||||
| 13C5-2′-AMP | 353 | 19 | 136 | 3.10 |
| 2′-AMP | 348 | 19 | 136 | 3.10 |
| Analysis of Inosine | ||||
| 15N4-Inosine | 273 | 20 | 141 | 3.10 |
| Inosine | 269 | 20 | 137 | 3.10 |
| Analysis of Hypoxanthine | ||||
| 13C5-Hypoxanthine | 141.8 | 22 | 124 | 1.86 |
| Hypoxanthine | 136.8 | 22 | 119 | 1.86 |
| Analysis of Xanthine | ||||
| 15N2-Xanthine | 154.9 | 20 | 137.8 | 2.00 |
| Xanthine | 152.9 | 20 | 135.8 | 2.00 |
| Analysis of Adenine | ||||
| 13C515N5-Adenine | 146 | 21 | 128 | 1.50 |
| Adenine | 136 | 21 | 119 | 1.50 |
| Internal Standard or Target Purine | Parent Ion (m/z) | Collision Energy (volts) | Daughter Ion (m/z) | Approximate Retention Time (min) |
|---|---|---|---|---|
| Analysis of Guanosine | ||||
| 13C10,15N5-Guanosine | 299 | 20 | 162 | 3.10 |
| Guanosine | 284 | 20 | 152 | 3.10 |
| Analysis of 3′,5′-cGMP | ||||
| 13C5-3′,5′-cGMP | 351 | 16 | 152 | 4.28 |
| 3′,5′-cGMP | 346 | 16 | 152 | 4.28 |
| Analysis of 2′,3′-cGMP | ||||
| 13C5-3′,5′-cGMP | 351 | 152 | 4.28 | |
| 2′,3′-cGMP | 346 | 16 | 152 | 2.96 |
| Analysis of 5′-GMP | ||||
| 13C10-5′-GMP | 374 | 15 | 157 | 1.76 |
| 5′-GMP | 364 | 15 | 152 | 1.76 |
| Analysis of 3′-GMP | ||||
| 13C10-5′-GMP | 374 | 15 | 157 | 1.76 |
| 3′-GMP | 364 | 15 | 152 | 2.43 |
| Analysis of 2′-GMP | ||||
| 13C10-5′-GMP | 374 | 15 | 157 | 1.76 |
| 2′-GMP | 364 | 15 | 152 | 3.80 |
| Analysis of Guanine | ||||
| 13C2,15N-Guanine | 155 | 20 | 138 | 1.56 |
| Guanine | 152 | 20 | 135 | 1.56 |
| Analysis of 8-Aminoguanosine | ||||
| 13C2,15N-8-Aminoguanosine | 302 | 17 | 170 | 3.64 |
| 8-Aminoguanosine | 299 | 17 | 167 | 3.64 |
| Analysis of 8-Aminoguanine | ||||
| 13C2,15N-8-Aminoguanine | 170 | 18 | 153 | 1.50 |
| 8-Aminoguanine | 167 | 18 | 150 | 1.50 |
| Characteristics | HNSCC Patients (n = 26) | |
|---|---|---|
| n | % | |
| Gender | ||
| Male | 18 | 69 |
| Female | 8 | 31 |
| Age at diagnosis (y) | ||
| Average ± SD | 64.2 ± 13.27 | |
| Range | 39–99 | |
| Primary tumor site | ||
| Larynx and pharynx | 8 | 31 |
| Oral cavity | 18 | 69 |
| Tumor extension | ||
| T1 | 6 | 23 |
| T2 | 6 | 23 |
| T3 | 3 | 12 |
| T4 | 4 | 15 |
| TX | 7 | 27 |
| Nodal involvement | ||
| N0 | 15 | 58 |
| N+ | 10 | 38 |
| NX | 1 | 4 |
| Nodal involvement | ||
| N1 | 4 | 15 |
| N2a | 0 | 0 |
| N2b | 3 | 12 |
| N2c | 3 | 12 |
| Distant metastases | ||
| M0 | 26 | 100 |
| M1 | 0 | 0 |
| Alcohol consumption | ||
| Yes | 12 | 46 |
| No | 14 | 54 |
| Tobacco consumption | ||
| Yes | 18 | 69 |
| No | 8 | 31 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ludwig, N.; Gillespie, D.G.; Reichert, T.E.; Jackson, E.K.; Whiteside, T.L. Purine Metabolites in Tumor-Derived Exosomes May Facilitate Immune Escape of Head and Neck Squamous Cell Carcinoma. Cancers 2020, 12, 1602. https://doi.org/10.3390/cancers12061602
Ludwig N, Gillespie DG, Reichert TE, Jackson EK, Whiteside TL. Purine Metabolites in Tumor-Derived Exosomes May Facilitate Immune Escape of Head and Neck Squamous Cell Carcinoma. Cancers. 2020; 12(6):1602. https://doi.org/10.3390/cancers12061602
Chicago/Turabian StyleLudwig, Nils, Delbert G. Gillespie, Torsten E. Reichert, Edwin K. Jackson, and Theresa L. Whiteside. 2020. "Purine Metabolites in Tumor-Derived Exosomes May Facilitate Immune Escape of Head and Neck Squamous Cell Carcinoma" Cancers 12, no. 6: 1602. https://doi.org/10.3390/cancers12061602