No Longer Underappreciated: The Emerging Concept of Astrocyte Heterogeneity in Neuroscience
by
 , and
, and
Francisco Pestana
1,†, 
Gabriela Edwards-Faret
2,3,†,
T. Grant Belgard
4,
Araks Martirosyan
1,* and
Matthew G. Holt
1,5,*
1
Laboratory of Glia Biology, VIB-KU Leuven Center for Brain and Disease Research, 3000 Leuven, Belgium
2
Laboratory of Neuronal Wiring, VIB-KU Leuven Center for Brain and Disease Research, 3000 Leuven, Belgium
3
Life & Medical Sciences (LIMES) Institute, Faculty of Mathematics and Natural Sciences, University of Bonn, 53115 Bonn, Germany
4
The Bioinformatics CRO, Niceville, FL 32578, USA
5
Leuven Brain Institute, KU Leuven, 3000 Leuven, Belgium
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Brain Sci. 2020, 10(3), 168; https://doi.org/10.3390/brainsci10030168
Submission received: 13 November 2019
/
Revised: 2 March 2020
/
Accepted: 5 March 2020
/
Published: 13 March 2020
(This article belongs to the Special Issue Understanding the Molecular Diversity of Astrocytes)
{kind=link}
{kind=link}
Abstract
:Astrocytes are ubiquitous in the central nervous system (CNS). These cells possess thousands of individual processes, which extend out into the neuropil, interacting with neurons, other glia and blood vessels. Paralleling the wide diversity of their interactions, astrocytes have been reported to play key roles in supporting CNS structure, metabolism, blood-brain-barrier formation and control of vascular blood flow, axon guidance, synapse formation and modulation of synaptic transmission. Traditionally, astrocytes have been studied as a homogenous group of cells. However, recent studies have uncovered a surprising degree of heterogeneity in their development and function, in both the healthy and diseased brain. A better understanding of astrocyte heterogeneity is urgently needed to understand normal brain function, as well as the role of astrocytes in response to injury and disease.
1. Introduction
Astrocytes comprise the largest class of glial cells in the mammalian brain, and are essential for central nervous system (CNS) development and function [1]. Compared to their neuronal counterparts, however, astrocytes remain understudied, largely due to the lack of tools allowing specific astrocyte labeling and manipulation. Hence, while neurons are known to display extensive molecular and functional diversity even within brain regions [2,3], astrocytes have traditionally been discussed as a homogeneous cell type [1], despite the wide range of key CNS functions that astrocytes are thought to participate in. These include processes as diverse as promoting synapse formation, maintaining synaptic homeostasis (through the control of extracellular K+ levels and removal of neurotransmitters), modulation of synaptic transmission, formation and maintenance of the blood-brain-barrier, control of cerebral blood flow and immune response [1]. This diversity of functions raises the obvious question of whether they are performed by all astrocytes, as presupposed, or whether functional subsets of astrocytes exist.
In a key review in 2010 [4], Ye Zhang and Ben Barres argued that astrocyte heterogeneity was an underappreciated topic in neurobiology. These authors made a compelling argument that a better understanding of astrocyte heterogeneity is necessary if we are to fully understand CNS function in both the healthy and injured (or diseased) brain. In the intervening period, substantial methodological improvements have led to an increased focus on the issue of astrocyte heterogeneity. As such, evidence is now accumulating which supports the importance of astrocyte heterogeneity throughout the CNS, both between and within brain regions. We hope that this review acts as a ‘primer’, quickly and clearly illustrating some of the emerging concepts in astrocyte heterogeneity, using selected examples in the areas of anatomy, developmental biology, molecular profiling, cell physiology, function and response to injury and disease. The majority of work described will have been conducted in rodents (mice and rats), unless otherwise stated, because of their wide use in astrocyte research. Where possible, we attempt to build on Zhang and Barres, with a focus on recent studies, particularly those that have used tissue slices, or better still in vivo models, to avoid the caveats associated with reduced complexity systems [5]. Readers are directed to excellent recent reviews to obtain a more complete view of the field [6,7,8].
2. Morphological Heterogeneity
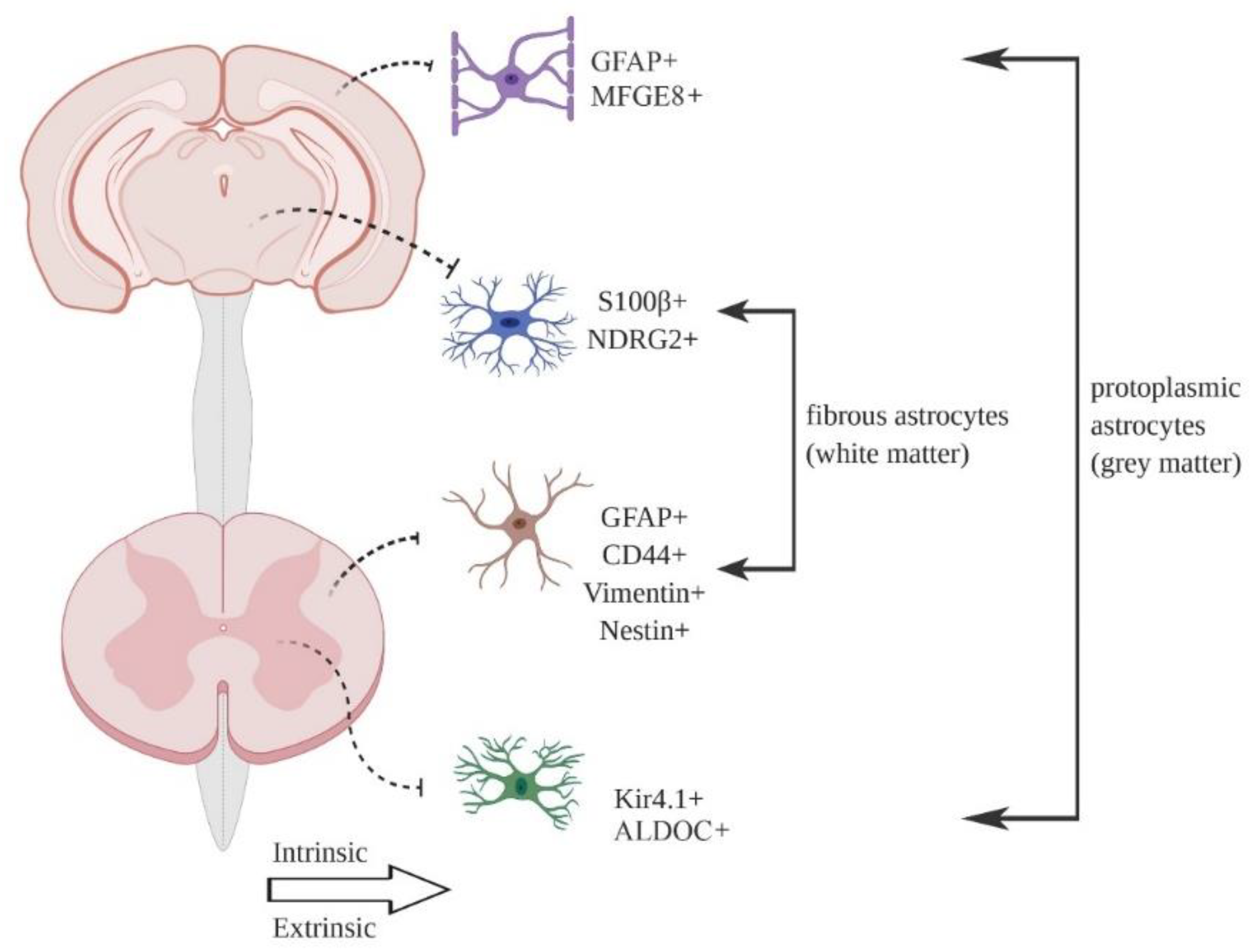
Astrocytes were first identified as a heterogeneous cell type based on gross morphology and differential expression of common marker proteins, such as the intermediate filament GFAP and glutamate transporter GLAST [7,9]. The most common division, based on morphology, is between protoplasmic astrocytes found in the grey matter and fibrous astrocytes found in the white matter (Figure 1). Additional morphological types in the mouse CNS include tanycytes, radial cells, Bergmann glia, velate glia, marginal glia, perivascular glia and ependymal glia [10]. Müller glia are a specialist astrocyte type found in the retina [1]. Multiple astrocyte morphologies may be found in the same brain region [9]. Generally speaking, individual astrocytes are thought to occupy distinct, non-overlapping spatial locations in the tissue (so-called tiling), with only minimal overlap between the ends of astrocyte processes [11], although a recent report using clonal analysis suggests that up to 20% of astrocytes show considerable spatial overlap in cortex [12]. Astrocyte density can vary considerably between regions [10,13], although individual cell counts always depend on the histological methodologies applied, which needs to be considered when comparing studies.
Protoplasmic astrocytes have an instantly recognizable morphology. They exhibit a spongiform profile, consisting of a central soma and four to ten major processes, which in turn branch off into thousands of smaller processes [8]. The tips of astrocyte processes ensheath synapses and blood vessels [1]. At the synapse, there is compelling evidence for bidirectional communication between neurons and astrocytes, with astrocytes thought to modulate synaptic transmission—a concept known as the ‘tripartite synapse’ [14]. Recent careful examination of cells using high-resolution fluorescence imaging has indicated subtle differences between protoplasmic astrocytes occupying distinct brain regions [15,16]. A systematic study of mouse hippocampal and striatal astrocytes, using iontophoresis of Lucifer Yellow dye, revealed that astrocytes in these regions have equivalent somatic volumes, number of main processes and total cell volumes. However, striatal astrocytes generally occupy a larger territory than hippocampal astrocytes and associate with more neuronal cell bodies, while hippocampal astrocytes associate with a larger number of excitatory synapses (an observation which was further confirmed with serial block face scanning electron microscopy) [15]. Such morphological heterogeneity also extends to astrocytes occupying the same brain region. The morphology of cortical protoplasmic astrocytes was investigated using cells outlined using a membrane tagged TdTomato combined with cytoskeletal GFP-labeling. Four prominent morphological subtypes were identified (based on consideration of multiple parameters including cell elongation, flatness and orientation), which distribute through the various cortical layers in differing proportions [16].
However, it is wrong to think that astrocyte morphology is fixed. Historically, there has been ample evidence that astrocyte processes are capable of changing their morphology (summarized by Theodosis and colleagues [17]). However, due to technical limitations associated with classical immunostaining and electron microscopy, such studies have tended to focus on either specialist synapses, such as the suprachiasmatic nucleus during circadian rhythmicity [18], or cortical synapses after prolonged periods of activity [19]. Recent advances in real-time imaging, using the astrocyte-specific dye sulforhodamine 101 (SR101) [20] or astrocyte-specific GFP expression [21], in both tissue slices and in vivo now suggest that modification of astrocytes processes can occur rapidly, in response to neuronal activity in mouse hippocampus and somatosensory cortex. Structural remodeling of this sort will have important consequences for astrocytic regulation of synaptic transmission, for example through the modulation of glutamate transporter proximity to synapses. However, this view of dynamic astrocyte processes has been recently challenged by in vivo experiments using overexpression of genetically encoded FRET sensors, which suggest that interactions between neurons and astrocytes are stable in the striatum [22]. The reasons for this discrepancy are unclear, but may be related to intrinsic differences in the biology of the hippocampus, cortex and striatum, differences in the methodologies used, or a combination of both.
There is now accumulating evidence that astrocyte processes exist as compartmentalized signaling units. In mature astrocytes, overexpression of a FLAG-tagged variant of the glutamate transporter GLT1 in combination with an anti-FLAG antibody conjugated to a quantum dot, allowed the movement of the transporter to be tracked [23]. Experiments show that GLT1 diffuses across the surface of the astrocyte process, becoming ‘trapped’ at synaptic sites, in order to remove glutamate from the synaptic cleft and terminate synaptic transmission. Despite the use of harsh solubilization conditions, GLT1 was isolated from brain extracts in a complex containing glycolytic enzymes and mitochondria [24], indicating that this complex is extremely stable when formed. In theory, such a complex provides a mechanism to spatially match energy production to transporter activity. It is tempting to link these two observations and speculate that capture of GLT1 into such a complex underlies its retarded movement. However, the proof for such a hypothesis remains lacking. Indeed, what could be the trigger for GLT1 capture? It is known that astrocytes respond to the activity of single synapses with compartmentalized Ca2+ signaling [25,26], and that mitochondrial positioning at synapses is dependent on neuronal activity and Ca2+ signaling [27,28]. Hence, the effects of Ca2+ in processes may play a central role in GLT1 capture.
Whether process remodeling is dependent on de novo protein synthesis or the redistribution/reuse of existing proteins is unclear at present. However, astrocyte processes contain mRNAs [29] and glial remodeling is blocked by inhibitors of protein synthesis, such as anisomycin [30]. Therefore, it appears likely that local translation (perhaps in an activity-dependent manner [31]) will emerge as playing a critical role.
Hence, although we now have a good understanding of basic astrocyte morphology throughout the brain, it remains unclear how key signaling machinery is distributed through the cell and how this distribution changes, if at all, upon astrocyte activity. Although technically demanding, experiments aimed at elucidating the composition of astrocyte processes are likely to be particularly revealing, given that these structures are in close proximity to synapses and blood vessels. Future technical improvements, such as (live) super-resolution imaging [32], combined with the ability to monitor mRNA trafficking [33] and local protein synthesis [34] will undoubtedly aid such investigations.
3. Developmental Heterogeneity
Since astrocyte heterogeneity was first recognized, a major question in the field has been how it is established. Is it set at the level of individual progenitor populations, or does local environment play a role? In reality, strong evidence exists for both possibilities.
Embryonic astrocytes derive from radial glia [35,36,37] and several lines of evidence indicate that astrocyte specification in the spinal cord and forebrain originates from a segmental template, induced by the action of known morphogens, such as sonic hedgehog (SHH), bone morphogenetic proteins (BMPs) and Wnt proteins, which induce the expression of specific transcription factors in discrete cell populations (comprehensively summarized by Ben Haim and Rowitch [7]). For example, the basic helix-loop-helix proteins Olig2 and SCL regulate oligodendrocyte versus astrocyte production in the p2 domain of the spinal cord [38], while the transcription factor Emx1 regulates astrocyte production in the cortical plate and corpus callosum [39].
Radial glia are no longer present in the post-natal mouse brain. Despite this, there is a rapid expansion in the number of cortical astrocytes during early post-natal development. Experiments combining retroviral labeling of dividing cells with GFP and immunostaining for astrocyte-specific BLBP (brain lipid binding protein) demonstrated that in cortical layers I -IV this expansion is due to the local proliferation of ‘pioneer’ astrocytes generated from radial glia [36], producing ‘clouds’ of clonally related cells [40]. However, the final distribution of astrocytes in the adult largely matches the original trajectory of their radial glia precursors [37,39], implying that astrocytes do not migrate tangentially and may retain spatial information at later developmental times, which is presumably crucial for the formation of the mature CNS. In fact, neither stab wound injury [39,41] nor toxin-mediated cell ablation [39] appear sufficient to overcome this strict regional allocation and induce mass migration of astrocytes to the injury site.
However, it appears that not all astrocyte functions are hardwired during development. In the cerebellum at least, neurons control astrocyte diversity through the release of SHH [42]. Crucially, Bergmann glia and velate astrocytes show considerable transcriptomic divergence, demonstrating the power of local signaling cues in determining fate. Whether this is a cerebellum specific mechanism remains to be determined. Certainly, the structure of the cerebellum with clearly demarcated cell locations and cell morphologies facilitates such studies. In contrast, cortical astrocytes are grossly similar at the anatomical level and large numbers of cells in the tissue share a common developmental lineage (see above). However, SHH is also produced in adult mouse cortex and SHH receptors are expressed on a proportion of cortical astrocytes [43]. This implies that the influence of local neuronal activity may be more subtle in cortex, with recent single cell data suggesting that local synaptic activity may influence astrocyte identity at the transcriptomic level (see Section 4). Of course, this does not preclude a contribution for other signaling pathways, such as fibroblast growth factor (FGF) signaling [15,44]. A potential role for pathways commonly associated with development is interesting, as evidence suggests that persistent activation of these pathways is necessary to maintain astrocyte diversity [45]. In this respect, a role for glutamatergic neuronal signaling in astrocyte development is not surprising, with genetic ablation of neuronal VGLUT1 adversely affecting post-natal development of cortical astrocytes [46]. It is also required for expression of the potassium channel Kir4.1 in astrocytes of the ventral horn of the spinal cord [47]. Furthermore, genetic manipulations affecting cortical layering, for example neuronal deletion of Dab1 [16] or Satb2 [48], also influence astrocyte positioning.
Hence, it appears cues from neighboring cells in the CNS cooperate with early patterning events to promote astrocyte diversity from a single progenitor.
4. Molecular Heterogeneity
In our opinion, the largest recent insights into astrocyte heterogeneity have been driven by experiments using transcriptome wide sequencing to reveal the underlying molecular machinery of the cell.
In a seminal study of cell type heterogeneity, Doyle and colleagues [49] introduced the TRAP (translating ribosome affinity purification technique) and showed considerable differences between the transcriptomes of cortical astrocytes, cerebellar astrocytes and cerebellar Bergmann glia.
However, it is wrong to think of the transcriptome as static. Using astrocyte-TRAP, Boisvert and colleagues [50] analyzed the transcriptome in cortex, hypothalamus and cerebellum and found region specific, aging-related changes (consistent with previous reports [51,52,53]). The fact that expression of mRNAs associated with synapse function decreased and expression of mRNAs associated with immune response increased suggests that astrocytes become pro-inflammatory during aging (see also a complementary study by Clarke et al. [54]), perhaps predisposing certain individuals to develop neurodegenerative disease [55] (as discussed in Section 7).
Morel and colleagues [56] performed a similar study, including hippocampus, cortex, nucleus accumbens, caudate putamen, thalamus and hypothalamus. They found a high degree of overlap between genes expressed in the cortex and hippocampus, while the nucleus accumbens, caudate putamen and hypothalamus showed expression of unique mRNAs not expressed in astrocytes from any other brain region studied. Amongst the most interesting transcripts identified were those for Hevin (Sparcl1) and SPARC (Sparc). Hevin is a pro-synaptogenic protein [57] and its mRNA was expressed at similar levels across all regions. SPARC, on the other hand, antagonizes Hevin action [57] and its mRNA was enriched in thalamic and hypothalamic astrocytes. Results were interpreted as a gradient of astrocyte gene expression along the dorsal-ventral axis of the brain, with consequences for excitatory synapse formation. Interestingly, the Boisvert study [50] also found evidence for changes in gene expression across the motor, somatosensory and visual cortices, indicative of a rostral-caudal gradient.
A further example of a gene expression gradient can be found in the work comparing hippocampal and striatal astrocytes [15]. These show differences in transcriptome (using TRAP) and proteome (using semi-quantitative liquid chromatography-tandem mass-spectrometry: LC-MS/MS) [15]. Interestingly, validation of the proteomic results with immunohistochemistry demonstrated that the protein µ-crystallin is differentially expressed amongst striatal astrocytes, occupying a gradient along the dorsal-ventral axis [15]. The functional consequences of this expression gradient are unknown at present.
Although the TRAP and MS experiments described here report data from whole astrocyte populations, these studies consistently indicate a complex picture of astrocyte gene expression with multiple axes of variation. Interestingly, even subtle changes in gene expression can have functional consequences: using cultured neurons and astrocytes in a ‘mix-and-match fashion’ revealed that astroglial modulation of synaptic maturation is region-selective [56].
The use of tissue microdissection and intersectional labeling strategies has contributed greatly to our understanding of intra-regional heterogeneity, particularly in cortex. Isolation of astrocytes from upper (2/3 and 4) and deeper (5 and 6) layers of cortex revealed differences in gene expression between the two sets of astrocytes [16,48]. These included genes involved in synapse formation and elimination, such as Sparc and Merkt [16] and Chrdl1 and Il33 [48] amongst others. A partial GLT1/EAAT2 promoter fragment drives TdTomato expression in a subgroup of astrocytes in cortical layer 5 [58], which express the synaptogenic protein Norrin (and see Section 7). A double reporter line (GLT1/EAAT2-TdTomato: Aldh1l1-eGFP) identifies three distinct classes of cortical astrocytes [59], with unique transcriptomes, tissue locations and electrophysiological characteristics. Staining with antibodies specific for CD51, CD63 and C71 in the Aldh1l1-eGFP mouse identifies five astrocyte subpopulations present in cortex, olfactory bulb, brain stem, thalamus, cerebellum and spinal cord [60], albeit in differing proportions. Interestingly, these populations vary in their synaptogenic potential.
In terms of defining the true extent of heterogeneity in a cell population, single cell methods provide, at least in theory, the truest measure. To date, such studies have generally reported a low degree of heterogeneity within specific brain regions. Gokce and colleagues [61] were unable to identify clear, distinct subtypes in the striatum, reporting a continuous transcriptional gradient defined by a unimodal population distribution, albeit with clear separation of genes encoding neurotransmitter transporters and glutamate receptors from those encoding ribosomal proteins and cell polarity regulators. This is in contrast to the proteomics-based findings from Chai and colleagues [15], which defined two distinct populations based on µ-crystallin expression. Zeisel and colleagues [62] identified 2 astrocyte subtypes in the somatosensory cortex and hippocampus, distinguished by differential expression of Gfap (which maps as expected to the pial layer) and Mfge8 (which extends through the remainder of the cortex). A more extensive follow up study by the same group [3] largely recapitulated these findings, with 7 astrocyte subtypes identified across the entire CNS. In another study, Saunders and colleagues [2] identified 8 distinct astrocyte types across 9 major brain regions.
At face value, these studies suggest that the number of discrete astrocyte subtypes in the CNS is rather low, certainly in comparison to reported neuronal diversity. However, the Gokce study [61] does raise the pertinent issue of how cell types are defined computationally, as opposed to identification of cell states. Furthermore, these studies have typically used a ‘one size fits all’ protocol, in which neither the tissue dissociation methods employed or the protocols used for sequencing library preparation have been optimized for a specific cell type, raising questions about whether astrocytes are subject to negative selection, based on issues of low viability after isolation, low mRNA content, etc. To investigate the issue of astrocyte diversity in more depth, our lab recently developed a protocol specifically for use with adult astrocytes and paired it with deep sequencing [63]. This work identified 5 distinct subtypes in cortex and hippocampus; two largely distinct groups identified as putative progenitors and three closely related subtypes likely to represent mature astrocytes. These subtypes contained markers identified in previous single cell studies, albeit over a wider number of astrocyte types, suggesting that the optimized protocol used in our study allows for a more subtle cell characterization. Interestingly, even though two of the ‘mature’ subtypes were largely intermixed in the cortex, they showed differential expression of transcripts involved in synapse formation and modulation of synaptic transmission, suggesting a fine degree of control over local CNS function. In our opinion, more targeted sequencing of astrocytes will also reveal a greater degree of heterogeneity across other brain regions.
Our findings of high levels of heterogeneity arising from subtle differences in transcript expression are further supported by a study from Bayraktar and colleagues [48], who took our single cell data set and by mapping back gene expression profiles to known layer specific markers, using large-scale fluorescence in situ hybridization (ISH), found evidence for both ‘laminar’ and ‘non-laminar’ gene expression in the dataset. Interestingly, laminar gene expression was offset from that classically defined by neuronal wiring patterns [48] and varied along both the dorsal-ventral and rostral-caudal axes.
Hence, the majority of evidence supports the fact that astrocytes are heterogeneous at the transcriptome level. There is evidence both for discrete patterns of gene expression restricted to specific brain regions, as well as for subtle gradients of gene expression spanning regions. Astrocytes may well share common genes, such as those regulating essential metabolic processes, but it appears there is sufficient variation in transcript expression to produce unique astrocyte subtypes, which are specialized to perform certain specific functions. As it stands, however, the majority of current single cell studies show considerable differences in the mouse lines used, age of mouse used, brain regions sampled, isolation method, number of astrocytes isolated, library preparation method employed and sequencing depth (summarized in [63]). These technical factors make it challenging to link subtypes identified by individual studies. This is an obstacle to overcome by systematic studies with the goal of building a comprehensive picture of astrocyte diversity across development and brain regions.
Finally, although the amount of data produced in single cell sequencing experiments can seem overwhelmingly complex, studies on single genes, such as Sema3a [64] and Kcnj10 [47] in the spinal cord (discussed in Section 6), have shown the power of gene expression data to inform hypothesis driven science. The development of new tools utilizing such data (for example ISH probes, antibodies, viral vectors and mouse lines) will undoubtedly be needed to aid future investigations.
5. Physiological Heterogeneity
Historically, astrocytes have been of little interest from an electrophysiological perspective, as they do not show the rapid fluctuations in membrane potential typical of action potential firing in neurons [65]. Generally speaking, mature astrocytes are characterized by a hyperpolarized resting potential that is set close to the equilibrium potential for K+ (EK) (approximately −80 mV), low input resistance (5−20 MΩ) and a linear current to voltage relationship, indicating a relative lack of voltage-dependent ionic conductances in the plasma membrane [65].
Fluctuations in membrane potential are tightly linked to changes in neuronal activity. Indeed, due to their strong, inward K+ conductance, astrocytes are highly sensitive to changes in extracellular K+ levels that are associated with neuronal activity, meaning they play a key role in regulating neuronal firing [66]. Interestingly, a recent comprehensive comparison of hippocampal and striatal astrocytes [15] found that they differ significantly in the size of their Ba2+-sensitive K+ currents. This suggests regional specialization, consistent with the fact that the striatum comprises predominantly GABAergic neurons with hyperpolarized membrane potentials, unlike the hippocampus which contains primarily glutamatergic neurons [15], meaning that striatal astrocytes have a lower requirement for K+ buffering and K+ dissipation. As expected, the difference in K+ conductance was also reflected in differential expression of genes encoding K+ channels, consistent with transcriptome informing function [15].
It has long been recognized that astrocytes can exist in extensive gap junction coupled networks (syncytia), mediated primarily by Connexins 30 and 43 [67]. The extent of gap junction coupling differs widely between brain regions [68,69] and is age dependent [70]. The connectivity of astrocyte networks can also be highly specific, with extensive gap junction coupling in the barrel cortex occurring between astrocytes in the same barrel, but limited coupling occurring between astrocytes from different barrels [71]. Gap junction coupling confers isopotentiality on a syncytium [72], minimizing depolarization due to elevated levels of local extracellular K+ and thereby maintaining a sustained driving force for highly efficient K+ uptake and maintenance of local homeostasis. Interestingly, although hippocampus and striatum show similar levels of gap junction coupling (approximately 100 coupled cells per network), the gap junction blocker carbenoxolone has much more profound effects in striatum [15], arguing for molecular heterogeneity in gap junctions [73] and suggesting caution when using this drug in future experiments. Independent of its function as an ion channel, Connexin 30 has also been reported to play a crucial role in regulated cell adhesion and migration [74], with genetic ablation experiments demonstrating a role for the protein during insertion of astrocyte processes into synaptic clefts. Crucially, Connexin 30 levels in astrocytes are controlled by neuronal activity, which effectively sets up a feedback loop to regulate synaptic transmission [75].
Astrocytes respond to various stimuli with increases in intracellular Ca2+ [6]. Such signaling has been intensively studied, as it is generally thought to represent the predominant form of cellular communication [76]. Generally, studies have been descriptive, with few reports of absolute Ca2+ levels in resting and activated astrocytes. Such measurements would undoubtedly be informative and contribute to our understanding of astrocyte function. Furthermore, technical limitations associated with the use of small organic indicators meant that historically measurements were usually limited to the cell soma [77]. Even at this level, differences in Ca2+ signals were reported, with marginal astrocytes in cortical layer 1 showing frequent asynchronous Ca2+ transients and astrocytes in layers 2/3 showing infrequent Ca2+ transients [78]. Recent studies, however, have taken advantage of the fact that genetically encoded calcium indicators (GECIs) can be expressed in a cell type specific manner and targeted to specific subcellular locations. They have revealed a rich diversity of Ca2+ signals in cells, ranging from the generation of signaling microdomains [79,80,81] through to global waves that encompass entire astrocytes, including their cell bodies [81,82]. Ca2+ responses may be spontaneous [79,83] or evoked by neuronal activity [25,26], with differences between brain regions reported; striatal and hippocampal astrocytes show differences in these two forms of Ca2+ signal [15]. Reported Ca2+ sources include channel-mediated Ca2+ entry [84], release from IP3R2-dependent intracellular stores [81,85] and release from mitochondria [86].
Crucially, a focus on intracellular Ca2+ elevations as activity signals may have artificially constrained our view of astrocyte function, as dopamine appears to both elevate and lower astrocyte Ca2+ in the stratum radiatum, depending on the signaling mechanisms used, Ca2+ source and the activity of extrusion mechanisms [87]. It is also important to note that oscillations in intracellular Ca2+ appear superimposed on a heterogeneous resting Ca2+ landscape within the cell [88]. Rat hippocampal astrocytes in acute tissue slices show a gradient of resting Ca2+ from the cell soma to their processes. This gradient is age dependent and astrocytes with high and low basal Ca2+ occupy contiguous space [88]. The level of resting Ca2+ is influenced by many factors, including Ca2+ leak from stores or through channels, the presence of fixed and mobile Ca2+ buffers, and extrusion mechanisms [89]. Hence, heterogeneity in the basal Ca2+ reflects the Ca2+ homeostasis machinery in cellular subcompartments, which will ‘sculpt’ the dynamics of evoked Ca2+ oscillations. Hence, Ca2+ signaling should no longer be considered a uniform blunt signal, but in all likelihood subtly encodes information in its dynamics. The introduction of next generation signal processing tools, capable of accurately describing the dynamics of (subcellular) Ca2+ transients, will undoubtedly help in revealing the subtleties of this signaling pathway [90].
However, a significant question remains unanswered: what is the function of these differing types of Ca2+ signal in astrocytes? It is tempting to speculate that they play functionally distinct roles. Interestingly, striatal and hippocampal astrocytes show differences in direct GPCR-mediated signaling (activated using DREADDs; Designer Receptors Exclusively Activated by Designer Drugs), with the Gi/o pathway more efficient in promoting Ca2+ transients in striatal astrocytes, suggesting differential expression of intracellular signaling cascades—although differences in DREADD expression levels and/or coupling into downstream signaling pathways cannot be completely excluded, due to the use of viral vectors for DREADD delivery [15]. This is further reinforced by the finding that expression of the intermediate early gene, c-fos, is greater in striatal astrocytes than hippocampal astrocytes following the activation of Gi coupled DREADD [15]. Cortical astrocytes show differential responses to the metabotropic agonist phenylephrine, which reflect the distribution of morphologically and transcriptomically distinct astrocyte populations [63]. Finally, work from our own lab suggests that local synaptic activity as well as the behavioral state of the animal are crucial in shaping intracellular Ca2+ signaling in the mouse visual cortex, suggesting that astrocytes act as signal integrators to produce distinct physiological effects [82]. This is in agreement with previous work showing the effects of the neuromodulator noradrenaline on astrocyte signaling [91].
Ca2+ mediated activities may include controlling cerebral blood flow [92] and modulating synaptic transmission [25], possibly through the Ca2+-mediated release of signaling molecules. Beyond Ca2+, the use of genetically encoded indicators for cAMP [93] and lactate [94] has revealed transient changes in the intracellular concentrations of these molecules under defined conditions, providing another layer of complexity to astrocyte signaling. It is likely that a combination of different signaling modalities, with different spatio-temporal properties, will control the myriad of functions performed by these cells.
6. Functional Heterogeneity
Perhaps the single most important advantage of astrocyte diversity is that it enables the creation of specialized neuron-glia units, which can drive complex behaviors. Multiple examples of distinct astrocyte subpopulations critical to key CNS functions have now been uncovered.
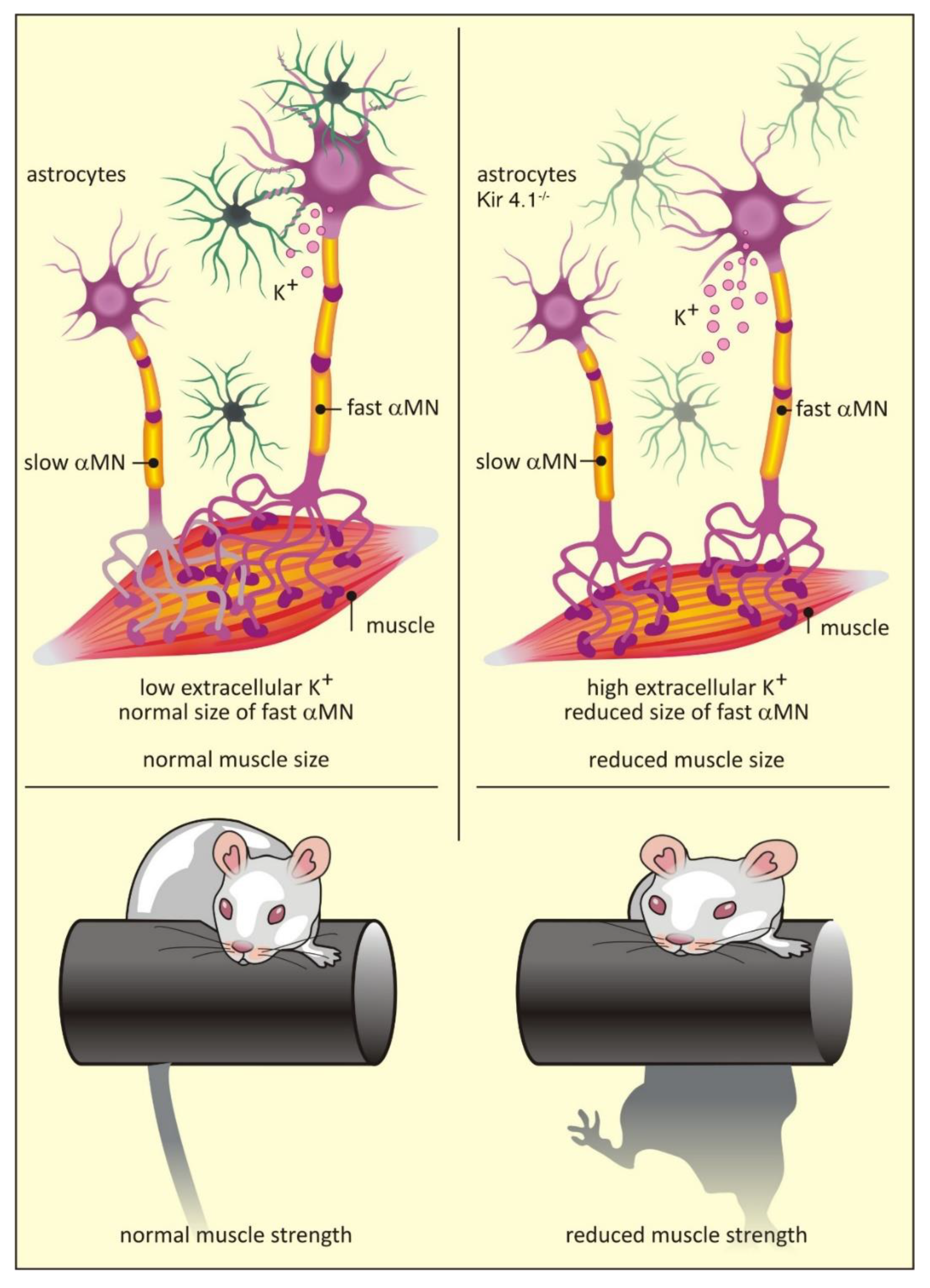
A comprehensive comparison of astrocytes isolated from the dorsal and ventral horns of the mouse spinal cord revealed distinct transcriptomic differences between the two populations [64]. In particular, ventral astrocytes were enriched in Sema3a transcripts, which encode an axon guidance cue that binds to the neuropilin 1 receptor on motor neurons [64]. Gene ablation experiments demonstrated a crucial role for astrocyte Sema3a in orientation and survival of α-motor neurons during development [64]. Similarly, ventral horn astrocytes are also enriched in transcripts for Kcnj10, which encode for the K+ channel Kir4.1 [47]. Unlike Sema3a, expression of Kcnj10 is dispensable for motor neuron survival [47]. However, loss of Kcnj10 expression in astrocytes does have non-cell autonomous effects on local neurons: fast α-motor neurons are smaller than wild type counterparts and show dramatically different electrophysiological profiles. These deficits correlate with reduced muscle size and decreased muscle strength [47] (Figure 2). Crucially, Kcnj10 levels appear reduced in astrocytes generated from amyotrophic lateral sclerosis (ALS) patients [47], providing a potential explanation for the muscle weakness seen in this disease [95] (see also Section 7.1).
Once formed, specialist circuits control a number of critical behaviors in the adult mouse. For example, astrocytes in the suprachiasmatic nucleus (SCN) drive the molecular oscillations underpinning circadian behavior in mammals, controlling clock gene expression in neurons via glutamatergic signaling [96]. Astrocytes also act as rhythm generators in the ventral brain stem area, responding to physiological decreases in pH with the release of ATP, which activates chemoreceptor neurons to induce adaptive changes in breathing [97]. They are also key for generating rhythmic activity in the rat trigeminal sensorimotor circuit responsible for mastication during feeding [98]. Insulin signaling in hypothalamic astrocytes is a key component in glucose metabolism, with the genetic ablation of insulin receptors in mice leading to hyperphagia and impaired regulation of systemic glucose levels [99]. Finally, homeostatic control of dopamine levels by astrocytes in the prefrontal cortex is essential for synaptic transmission and plasticity, playing a crucial role in memory and behavioral flexibility [100]. Likewise, the activation of astrocytes (but not neurons) in hippocampal CA1, using a Gq-coupled DREADD, was shown to enhance memory acquisition in mice [101].
Crucially, however, functional heterogeneity may not be restricted to region-specific astrocytes. Dopamine receptor D1 and dopamine receptor D2 containing medium spiny neuronal subtypes are intermixed in the dorsal striatum [102]. When stimulated, endocannabinoid release from these neurons leads to increases in intracellular Ca2+ in different subsets of striatal astrocytes [102]. Crucially, Ca2+ uncaging experiments in these specific astrocytes evoked a transient synaptic potential only in homotypic pairs of synaptically connected neurons [102].
Taken together, these examples provide clear proof that specialized astrocyte subsets are responsible for the function of specific neuronal circuits, and are capable of synapse-specific regulation. Using the increasingly comprehensive gene expression data now available, it is likely that further examples of astrocyte specialization will be uncovered in the immediate future. However, given the complex transcriptional ‘fingerprints’ which define astrocyte subtypes [48,63], sophisticated intersectional strategies (e.g., split-Cre systems) will be needed for specific labeling and manipulation [103].
7. Heterogeneity in Injury and Disease
Astrocytes are involved in all neuropathologies, ranging from acute injury through to chronic neurodegenerative conditions. However, how astrocytes are involved in these different pathologies remains unclear and needs to be addressed. Even the basic issue of whether astrocytes are primary initiators or modifiers of disease is debatable, and probably depends on the condition in question [6]. As it increasingly appears that all injuries and diseases elicit a unique astrocytic response, a comprehensive description of astrocyte involvement in all conditions is beyond the scope of this work. Rather, in this section, we have chosen specific examples which we think best illustrate key emerging concepts relating to astrocyte heterogeneity in injury and disease. However, for readers interested in specific conditions, in depth reviews can generally be found (for example Alexander’s disease [104], Alzheimer’s disease (AD) [105] and ischemic injury [106]).
7.1. Astrocyte-Specific Diseases
At the time of writing, only Alexander’s disease has been categorically demonstrated to be a primary astrocyte disease, due to mutations in the Gfap gene, which is specifically expressed in astrocytes [107]. More than 50 mutations have been reported and associated with development of the disease. Although it shows variable age of onset, being defined as ‘infant’ (0–2 years), ‘juvenile’ (2–12 years) or ‘adult’ (>12 years), a common phenotype is the formation of Rosenthal fibers in astrocytes, thought to be indicative of cell stress and loss of internal cell homoeostasis [104]. The destruction of myelin characteristic to the disease [108] typically relates to the age of disease onset and matches the regional distribution of GFAP expressing astrocytes in the CNS [109], which are found in large numbers in the white matter and provide metabolic support to myelinating oligodendrocytes [104].
Recent evidence now suggests that Norrie disease, which is caused by mutations in the Ndp (Norrie disease pseudoglioma) gene located on the X-chromosome, may be linked to the secretion of active Norrin protein from distinct astrocyte subsets and subsequent activation of the Wnt signaling pathway in surrounding cells [58]. Consistent with the involvement of Wnt signaling in CNS patterning [110] and synapse formation [111], patients generally present with blindness. Studies in a mouse model carrying a mutation in exon 2 of the Ndp gene [112], which results in the deletion of 56 amino acids from the N-terminus of the protein, suggest that this is due to disorganization of the ganglion cell layer and loss of the outer plexiform layer in the retina. Approximately 30%–50% of males with Norrie disease also present with intellectual disability, behavioral abnormalities, or psychotic-like features [113], consistent with the abnormal dendrite patterning and spine formation, impaired synaptic transmission and behavioral abnormalities seen in the Norrin knockout mouse [58,114].
Expression of Kir4.1 is heavily enriched in astrocytes associated with motor neurons in the ventral spinal cord. Reduced levels of Kir4.1 lead to a specific reduction in the size of fast twitch α-motor neurons and loss of muscle strength [47] (Figure 2; see above). Amyotrophic lateral sclerosis (ALS) is a degenerative disease which mainly affects the lower motor neurons in the brainstem and ventral horn of the spinal cord, with fast twitch α-motor neurons most severely affected [115]. ALS is familial in approximately 10% of cases with mutations in the superoxide dismutase 1 (SOD1) gene found in 20% of affected families [116]. Interestingly, astrocytes derived from ALS patients with a SOD1D90A mutation showed a cell autonomous reduction in Kir4.1 levels [47]. Although there was no evidence for enhanced loss of motor neurons in a SOD mutant mouse model following genetic ablation of Kir4.1, this study suggests that the initial clinical presentation of weakness in ALS may be related to astrocyte-specific down-regulation of Kir4.1.
7.2. Astrocyte Response to External Injury or Disease
Generally speaking, injury or disease is associated with morphological changes and upregulation of classical markers in astrocytes, such as GFAP, a process known as reactive gliosis [117]. In the case of severe insults, such as paralysis-inducing spinal cord injury resulting from spinal cord crush [118] or lesion [119], the consequences are major tissue remodeling and the formation of a prominent glial scar. The glial scar derives almost entirely from newly proliferated astrocytes with elongated shape, whose processes interdigitate forming a barrier (scar) around the injury site [117]. The role of the glial scar is heavily debated and appears double-edged. On the one hand, it is thought the scar acts to seal off the damaged area immediately following injury to prevent the escape of toxic species, such as oxygen free radicals and proteases released from damaged cells, which would otherwise spread through the CNS causing damage [117]. Unfortunately, however, the changes underlying scar formation are effectively irreversible; as regenerating axons do not pass through the glial scar, reinnervation of affected areas and functional recovery is effectively prevented [117]. The basis of this negative effect was long thought to be the expression of inhibitory chondroitin sulfate proteoglycans (CSPGs) by scar forming astrocytes [117]. However, this view has recently been challenged in a spinal crush model [118], where the specific targeting of reactive astrocytes, either by genetic inactivation of the key regulatory protein STAT3 or through cell ablation using diphtheria toxin, failed to stimulate axon regrowth and actually compromised tissue integrity. Surprisingly, in this study, levels of inhibitory CSPGs were maintained in the environment of the scar in the absence of reactive gliosis; in fact, transcriptome sequencing of reactive astrocytes revealed that these cells actually express CSPG types supportive of axonal growth [118]. The reason for the discrepancy between studies is unclear, but may be due to the sustained expression of inhibitory CSPGs from other cells types (e.g., fibroblasts and pericytes) which are minor components of the glial scar or differential response to injury type (as discussed below).
In other injuries, such as stab wound models in the hippocampus [120] and cortex [41], morphological changes are more moderate, with cells generally displaying varying degrees of hypertrophy but maintaining the tiling arrangement seen in healthy tissue [6]. While studies using classical immunohistochemistry-based techniques have revealed important aspects of the injury response, by definition they represent a snap-shot of the tissue at a fixed time point. Advances with in vivo imaging now allow the whole injury response to be evaluated over time. Crucially, the response to cortical stab wound injury appears to be heterogeneous [41], with a subset of astrocytes appearing hypertrophic, another directing processes towards the lesion and a distinct subset undergoing limited proliferation at juxtavascular sites. Given the intimate association of astrocytes with neurons and other CNS cell types, it is highly likely that any morphological rearrangement which affects this exquisite interaction will impact on CNS function [1]. Perhaps unsurprisingly, cellular hypertrophy in reactive astrocytes also leads to the redistribution of integral membrane proteins. In the APP/PS1 mouse model of Alzheimer’s type amyloidosis there is a redistribution of the bestrophin1 (Best1) channel from astrocyte processes to the cell soma, which is concomitant with increased GABA release from astrocytes and reduced firing probability in granule cells [121]. It is likely that aberrant synaptic function is one of the first events in chronic neurodegenerative diseases, such as Alzheimer’s, which are ultimately characterized by synapse loss and neuronal death. Interestingly, there are indications that in humans astrocyte dysfunction occurs first, with uptake of a PET tracer for monoamine oxidase B (MAO-B), a critical enzyme in the GABA synthesis pathway, positively correlating to uptake of the amyloid marker Pittsburgh Compound B [122]. In this case, however, it needs to be remembered that MAO-B is also expressed in serotonergic neurons, which may influence data interpretation, despite serotonergic neurons comprising a comparatively small population of cells in the adult human brain.
Injury and disease also promote massive changes in gene expression levels, with some genes increasing by over 100-fold [123]. Consistent with the well-established concept of inter-regional astrocyte heterogeneity, TRAP experiments in a common mouse model of multiple sclerosis (experimental autoimmune encephalomyelitis) showed profound differences in gene expression between spinal cord, cerebellum, cerebral cortex and hippocampus in the disease state [124]. Whole transcriptome sequencing of FACS isolated astrocytes from mouse models of bacterial infection (lipopolysaccharide, LPS) [123] and stroke (middle cerebral artery occlusion, MCAO) [123] indicated that astrocytes have profoundly different responses depending on insult. Generally speaking, LPS treatment results in the upregulation of genes typically associated with a pro-inflammatory response, including components of the innate immune system (complement cascade) and pro-inflammatory cytokines. In contrast, MCAO generally results in the enhanced expression of genes associated with an anti-inflammatory response, including neurotrophic factors and anti-inflammatory cytokines. This has led to the introduction of two classes for reactive astrocytes, the so-called pro-inflammatory A1 type and neuroprotective A2 type. This classification is based on the concept of M1/M2 polarization in macrophages [125]. Pro-inflammatory A1 astrocytes are toxic to mouse neurons in co-cultures and have been identified in human samples from patients suffering from AD, ALS, Parkinson’s disease (PD) and multiple sclerosis [126], suggesting that neuronal loss in these diseases results, at least in part, due to reactive astrogliosis. Although the A1/A2 framework is conceptually useful, it should be remembered that the original M1/M2 classification has since proved to be problematic, as it represents the ends of a continuum which rarely, if ever, exists in vivo [125]. Consistent with this, a recent study of transcriptional changes in human caudate samples of Huntington’s disease found little evidence for a clear A1 or A2 phenotype [127], with evidence for both inflammatory states suggesting astrocyte response during human disease is a highly complex process.
The degree to which astrocytes can discriminate between insults, however, remains unclear at this point. For example, whether astrocytes can distinguish between subtle conformational differences in toxic protein species [128], including amyloid beta in AD [129], tau in AD [130] or α-synuclein in PD [131] remains to be determined, as well as whether this initiates a specific cellular response. However, it does appear that response is graded to the insult. Astrocyte responses to both LPS and MCAO appear variegated within tissue, with reactive astrocytes highly expressing Lcn2 and Serpina3n interspersed with quiescent astrocytes [123]. In the APP/PS1 mouse model of Alzheimer’s, astrocytes close to amyloid beta plaques show upregulation of MAO-B. Along with redistribution of the Best1 channel (see earlier), MAO-B upregulation is thought to underlie the abnormal circuit firing in this model observed at early stages of amyloidosis, due to the aberrant release of GABA [121]. Responses may also be affected by the age of an animal, as TRAP experiments in aged mice revealed loss of synaptic support functions (such as cholesterol production) and a switch towards a pro-inflammatory state [50,54]. As these changes appear mirrored in the aging human brain [55], and aging is the single biggest risk factor for neurodegenerative disease, this raises the question of how chronic low-level inflammation contributes to these conditions. Whether differences in astrocyte response also explain regional susceptibility to disease, such as seen with the substantia nigra in PD, also remains unclear [132].
Toxins which build up in the CNS need to be efficiently removed, but the CNS lacks a traditional lymphatic system, which usually serves in such a capacity. Recently, however, Nedergaard and colleagues have advanced the concept of the glia-lymphatic (glymphatic) system, a highly polarized transport system for cerebrospinal fluid (CSF) and interstitial fluid (ISF) in the brain that facilitates extracellular waste removal through a network of astrocyte-supported perivascular or perineural channels that drain into the cervical and basal meningeal lymphatic networks or the major dural sinuses [133]. Performance of the glymphatic system is impaired with aging [134] and may be a contributory factor to the development of chronic neurodegenerative disease. In fact, impairment of glymphatic function adversely affects amyloid beta removal in the APP/PS1 mouse model, suggesting that the system is a critical modulator of AD progression [135]. Crucially, impaired glymphatic function has also been shown in animal models of traumatic brain injury [136] and small ischemic lesions [137], suggesting that toxin build up, due to inefficient removal, plays a significant secondary role in tissue damage, following the initial trauma. The impact of astrocyte heterogeneity on the efficiency of this system, and whether it contributes to regional susceptibility to disease, also needs to be explored. However, two observations point towards the glymphatic system being a general system of waste removal. First, efficient toxin removal is central to CNS function and survival. Second, the astrocyte-specific water channel Aquaporin4 (Aqp4) is central to the system and is one of the few markers expressed across all astrocytes in major single cell sequencing studies performed to date [2,3,63].
Whether astrocytes can eventually revert to their pre-injury state is unclear, but likely depends on insult. For example, although most gene expression changes induced by LPS or MCAO are transient, several genes remain elevated one week after the insult [123] and this may have functional consequences. A better understanding of the changes occurring in reactive astrocytes may lead to interesting avenues for brain repair following injury or disease. It appears a limited number of astrocytes in the mouse striatum and medial cortex can reactivate a Notch-dependent latent neurogenic program following a stroke, resulting in the generation of new neurons [138]. Crucially, however, even though this intrinsic repair seems limited, it does appear that the reactive astrocyte state could be harnessed for brain repair. Reactive cortical astrocytes in the 5xFAD Alzheimer’s model appear to be more easily converted into neurons by retroviral-mediated overexpression of the neurogenic transcription factor NeuroD1 [139], as do reactivate astrocytes generated by a cortical stab wound injury [139]. These induced neurons possess Na+ and K+ currents, generate action potentials and show spontaneous and evoked responses, arguing for integration into local circuits. Despite the obvious potential of such a system, it remains unclear why the neurons generated by NeuroD1 reprogramming are mainly glutamatergic and express markers typically found in neurons in deep cortical layers [139]. Whether this will impact full functional recovery needs to be systematically assessed across injuries. Understanding the interconversion process, including whether injury causes partial dedifferentiation of astrocytes into a more ‘stem-like’ state [52], should prove informative and identify specific transcriptional pathways that can be targeted for production of specific neuronal types. A further limitation of the system appears to be the resistance of white matter astrocytes to reprogramming [140], which needs to be considered as primates possess more white matter than rodents.
As evidence for discrete subsets of human astrocytes emerges with specific functions, in both healthy [141] and diseased tissues [60], it seems likely that similar concepts of heterogeneous and graded response, age dependency, as well as waste removal and potential for interconversion, will be found.
8. Conclusions
Based on a number of criteria, astrocytes have emerged as far more heterogeneous than previously thought and are critically importantly for formation and function of the healthy CNS. Perturbed function of specific astrocyte subsets can lead to disease, while astrocyte response to external injury and/or toxic insults is also heterogeneous. Failure to understand and modify astrocyte response to re-establish CNS homeostasis may explain the continued failure of CNS drugs to reach the clinic, and represents a currently undervalued concept in therapeutics development.
Ultimately, continued improvements in tool development, allowing the selective labeling and manipulation of specific astrocyte subsets, will be key to furthering our understanding on how astrocyte subtypes contribute to CNS development and function, both in the normal and pathological CNS.
Author Contributions
F.P., G.E.-F. and A.M. worked individually on subsections with input from M.G.H. Integration of subsections into a final text was done by M.G.H. Input into the various manuscript drafts was provided by T.G.B. All authors have read and agreed to the published version of the manuscript.
Funding
G.E.-F. is a post-doctoral fellow of the Fonds Wetenschappelijk Onderzoek (FWO) (Grant 1214420N). A.M. holds a Stichting Alzheimer Onderzoek (SAO) Pilot Grant (20180024). M.G.H. grateful acknowledges support from the European Research Council (Starting Grant 281961), FWO (Grants G066715N and 1523014N), SAO (S#16025), VIB Institutional Support and VIB Tech Watch funding.
Acknowledgments
Conflicts of Interest
T.G.B. is currently chief executive officer at The Bioinformatics CRO and head of bioinformatics at Bit Bio. The remaining authors have no known possible conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Barres, B.A. The mystery and magic of glia: A perspective on their roles in health and disease. Neuron 2008, 60, 430–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, A.; Macosko, E.Z.; Wysoker, A.; Goldman, M.; Krienen, F.M.; de Rivera, H.; Bien, E.; Baum, M.; Bortolin, L.; Wang, S.; et al. Molecular diversity and specializations among the cells of the adult mouse brain. Cell 2018, 174, 1015–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisel, A.; Hochgerner, H.; Lönnerberg, P.; Johnsson, A.; Memic, F.; van der Zwan, J.; Häring, M.; Braun, E.; Borm, L.E.; La Manno, G.; et al. Molecular architecture of the mouse nervous system. Cell 2018, 174, 999–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Barres, B.A. Astrocyte heterogeneity: An underappreciated topic in neurobiology. Curr. Opin. Neurobiol. 2010, 20, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Foo, L.C.; Allen, N.J.; Bushong, E.A.; Ventura, P.B.; Chung, W.S.; Zhou, L.; Cahoy, J.D.; Daneman, R.; Zong, H.; Ellisman, M.H.; et al. Development of a method for the purification and culture of rodent astrocytes. Neuron 2011, 71, 799–811. [Google Scholar] [CrossRef] [Green Version]
- Khakh, B.S.; Sofroniew, M.V. Diversity of astrocyte functions and phenotypes in neural circuits. Nat. Neurosci. 2015, 18, 942–952. [Google Scholar] [CrossRef]
- Ben Haim, L.; Rowitch, D.H. Functional diversity of astrocytes in neural circuit regulation. Nat. Rev. Neurosci. 2017, 18, 31–41. [Google Scholar] [CrossRef]
- Khakh, B.S.; Deneen, B. The emerging nature of astrocyte diversity. Annu. Rev. Neurosci. 2019, 42, 187–207. [Google Scholar] [CrossRef]
- Matyash, V.; Kettenmann, H. Heterogeneity in astrocyte morphology and physiology. Brain Res. Rev. 2010, 63, 2–10. [Google Scholar] [CrossRef]
- Emsley, J.G.; Macklis, J.D. Astroglial heterogeneity closely reflects the neuronal-defined anatomy of the adult murine CNS. Neuron Glia Biol. 2006, 2, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Ogata, K.; Kosaka, T. Structural and quantitative analysis of astrocytes in the mouse hippocampus. Neuroscience 2002, 113, 221–233. [Google Scholar] [CrossRef]
- Clavreul, S.; Abdeladim, L.; Hernández-Garzón, E.; Niculescu, D.; Durand, J.; Leng, S.H.; Barry, R.; Bonvento, G.; Beaurepaire, E.; Livet, J.; et al. Cortical astrocytes develop in a plastic manner at both clonal and cellular levels. Nat. Commun. 2019, 10, 4884. [Google Scholar] [CrossRef] [PubMed]
- Keller, D.; Erö, C.; Markram, H. Cell densities in the mouse brain: A Systematic Review. Front. Neuroanat. 2018, 12, 83. [Google Scholar] [CrossRef] [Green Version]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite synapses: Astrocytes process and control synaptic information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.; Diaz-Castro, B.; Shigetomi, E.; Monte, E.; Octeau, J.C.; Yu, X.; Cohn, W.; Rajendran, P.S.; Vondriska, T.M.; Whitelegge, J.P.; et al. Neural circuit-specialized astrocytes: Transcriptomic, proteomic, morphological, and functional evidence. Neuron 2017, 95, 531–549. [Google Scholar] [CrossRef]
- Lanjakornsiripan, D.; Pior, B.J.; Kawaguchi, D.; Furutachi, S.; Tahara, T.; Katsuyama, Y.; Suzuki, Y.; Fukazawa, Y.; Gotoh, Y. Layer-specific morphological and molecular differences in neocortical astrocytes and their dependence on neuronal layers. Nat. Commun. 2018, 9, 1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodosis, D.T.; Poulain, D.A.; Oliet, S.H. Activity-dependent structural and functional plasticity of astrocyte-neuron interactions. Physiol. Rev. 2008, 88, 983–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becquet, D.; Girardet, C.; Guillaumond, F.; François-Bellan, A.M.; Bosler, O. Ultrastructural plasticity in the rat suprachiasmatic nucleus. Possible involvement in clock entrainment. Glia 2008, 56, 294–305. [Google Scholar] [CrossRef]
- Genoud, C.; Quairiaux, C.; Steiner, P.; Hirling, H.; Welker, E.; Knott, G.W. Plasticity of astrocytic coverage and glutamate transporter expression in adult mouse cortex. PLoS Biol. 2006, 4, e343. [Google Scholar] [CrossRef] [Green Version]
- Perez-Alvarez, A.; Navarrete, M.; Covelo, A.; Martin, E.D.; Araque, A. Structural and functional plasticity of astrocyte processes and dendritic spine interactions. J. Neurosci. 2014, 34, 12738–12744. [Google Scholar] [CrossRef]
- Bernardinelli, Y.; Randall, J.; Janett, E.; Nikonenko, I.; König, S.; Jones, E.V.; Flores, C.E.; Murai, K.K.; Bochet, C.G.; Holtmaat, A.; et al. Activity-dependent structural plasticity of perisynaptic astrocytic domains promotes excitatory synapse stability. Curr. Biol. 2014, 24, 1679–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Octeau, J.C.; Chai, H.; Jiang, R.; Bonanno, S.L.; Martin, K.C.; Khakh, B.S. An optical neuron-astrocyte proximity assay at synaptic distance scales. Neuron 2018, 98, 49–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy-Royal, C.; Dupuis, J.P.; Varela, J.A.; Panatier, A.; Pinson, B.; Baufreton, J.; Groc, L.; Oliet, S.H. Surface diffusion of astrocytic glutamate transporters shapes synaptic transmission. Nat. Neurosci. 2015, 18, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Genda, E.N.; Jackson, J.G.; Sheldon, A.L.; Locke, S.F.; Greco, T.M.; O’Donnell, J.C.; Spruce, L.A.; Xiao, R.; Guo, W.; Putt, M.; et al. Co-compartmentalization of the astroglial glutamate transporter, GLT-1, with glycolytic enzymes and mitochondria. J. Neurosci. 2011, 31, 18275–18288. [Google Scholar] [CrossRef] [Green Version]
- Di Castro, M.A.; Chuquet, J.; Liaudet, N.; Bhaukaurally, K.; Santello, M.; Bouvier, D.; Tiret, P.; Volterra, A. Local Ca2+ detection and modulation of synaptic release by astrocytes. Nat. Neurosci. 2011, 14, 1276–1284. [Google Scholar] [CrossRef]
- Panatier, A.; Vallée, J.; Haber, M.; Murai, K.K.; Lacaille, J.C.; Robitaille, R. Astrocytes are endogenous regulators of basal transmission at central synapses. Cell 2011, 146, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.G.; O’Donnell, J.C.; Takano, H.; Coulter, D.A.; Robinson, M.B. Neuronal activity and glutamate uptake decrease mitochondrial mobility in astrocytes and position mitochondria near glutamate transporters. J. Neurosci. 2014, 34, 1613–1624. [Google Scholar] [CrossRef] [Green Version]
- Stephen, T.L.; Higgs, N.F.; Sheehan, D.F.; Al Awabdh, S.; López-Doménech, G.; Arancibia-Carcamo, I.L.; Kittler, J.T. Miro1 regulates activity-driven positioning of mitochondria within astrocytic processes apposed to synapses to regulate intracellular calcium signaling. J. Neurosci. 2015, 35, 15996–16011. [Google Scholar] [CrossRef]
- Sakers, K.; Lake, A.M.; Khazanchi, R.; Ouwenga, R.; Vasek, M.J.; Dani, A.; Dougherty, J.D. Astrocytes locally translate transcripts in their peripheral processes. Proc. Natl. Acad. Sci. USA 2017, 114, E3830–E3838. [Google Scholar] [CrossRef] [Green Version]
- Langle, S.L.; Poulain, D.A.; Theodosis, D.T. Induction of rapid, activity-dependent neuronal-glial remodelling in the adult rat hypothalamus in vitro. Eur. J. Neurosci. 2003, 18, 206–214. [Google Scholar] [CrossRef]
- Hasel, P.; Dando, O.; Jiwaji, Z.; Baxter, P.; Todd, A.C.; Heron, S.; Márkus, N.M.; McQueen, J.; Hampton, D.W.; Torvell, M.; et al. Neurons and neuronal activity control gene expression in astrocytes to regulate their development and metabolism. Nat. Commun. 2017, 8, 15132. [Google Scholar] [CrossRef] [PubMed]
- Berning, S.; Willig, K.I.; Steffens, H.; Dibaj, P.; Hell, S.W. Nanoscopy in a living mouse brain. Science 2012, 335, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelles, D.A.; Fang, M.Y.; O’Connell, M.R.; Xu, J.L.; Markmiller, S.J.; Doudna, J.A.; Yeo, G.W. Programmable RNA tracking in live cells with CRISPR/Cas9. Cell 2016, 165, 488–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tom Dieck, S.; Kochen, L.; Hanus, C.; Heumüller, M.; Bartnik, I.; Nassim-Assir, B.; Merk, K.; Mosler, T.; Garg, S.; Bunse, S.; et al. Direct visualization of newly synthesized target proteins in situ. Nat. Methods 2015, 12, 411–414. [Google Scholar] [CrossRef] [Green Version]
- Malatesta, P.; Hartfuss, E.; Götz, M. Isolation of radial glial cells by fluorescent-activated cell sorting reveals a neuronal lineage. Development 2000, 127, 5253–5263. [Google Scholar] [PubMed]
- Ge, W.P.; Miyawaki, A.; Gage, F.H.; Jan, Y.N.; Jan, L.Y. Local generation of glia is a major astrocyte source in postnatal cortex. Nature 2012, 484, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magavi, S.; Friedmann, D.; Banks, G.; Stolfi, A.; Lois, C. Coincident generation of pyramidal neurons and protoplasmic astrocytes in neocortical columns. J. Neurosci. 2012, 32, 4762–4772. [Google Scholar] [CrossRef]
- Muroyama, Y.; Fujiwara, Y.; Orkin, S.H.; Rowitch, D.H. Specification of astrocytes by bHLH protein SCL in a restricted region of the neural tube. Nature 2005, 438, 360–363. [Google Scholar] [CrossRef]
- Tsai, H.H.; Li, H.; Fuentealba, L.C.; Molofsky, A.V.; Taveira-Marques, R.; Zhuang, H.; Tenney, A.; Murnen, A.T.; Fancy, S.P.; Merkle, F.; et al. Regional astrocyte allocation regulates CNS synaptogenesis and repair. Science 2012, 337, 358–362. [Google Scholar] [CrossRef] [Green Version]
- García-Marqués, J.; López-Mascaraque, L. Clonal identity determines astrocyte cortical heterogeneity. Cereb. Cortex 2013, 23, 1463–1472. [Google Scholar] [CrossRef] [Green Version]
- Bardehle, S.; Krüger, M.; Buggenthin, F.; Schwausch, J.; Ninkovic, J.; Clevers, H.; Snippert, H.J.; Theis, F.J.; Meyer-Luehmann, M.; Bechmann, I.; et al. Live imaging of astrocyte responses to acute injury reveals selective juxtavascular proliferation. Nat. Neurosci. 2013, 16, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Farmer, W.T.; Abrahamsson, T.; Chierzi, S.; Lui, C.; Zaelzer, C.; Jones, E.V.; Bally, B.P.; Chen, G.G.; Théroux, J.F.; Peng, J.; et al. Neurons diversify astrocytes in the adult brain through sonic hedgehog signaling. Science 2016, 351, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.D.; Petrova, R.; Eng, L.; Joyner, A.L. Sonic hedgehog regulates discrete populations of astrocytes in the adult mouse forebrain. J. Neurosci. 2010, 30, 13597–13608. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Goldman, J.E. An FGF-responsive astrocyte precursor isolated from the neonatal forebrain. Glia 2009, 57, 592–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmer, W.T.; Murai, K. Resolving astrocyte heterogeneity in the CNS. Front. Cell Neurosci. 2017, 11, 300. [Google Scholar] [CrossRef]
- Morel, L.; Higashimori, H.; Tolman, M.; Yang, Y. VGluT1+ neuronal glutamatergic signaling regulates postnatal developmental maturation of cortical protoplasmic astroglia. J. Neurosci. 2014, 34, 10950–10962. [Google Scholar] [CrossRef]
- Kelley, K.W.; Ben Haim, L.; Schirmer, L.; Tyzack, G.E.; Tolman, M.; Miller, J.G.; Tsai, H.H.; Chang, S.M.; Molofsky, A.V.; Yang, Y.; et al. Kir4.1-dependent astrocyte-fast motor neuron interactions are required for peak strength. Neuron 2018, 98, 306–319. [Google Scholar] [CrossRef]
- Bayraktar, O.A.; Bartels, T.; Holmqvist, S.; Kleshchevnikov, V.; Martirosyan, A.; Polioudakis, D.; Ben Haim, L.; Young, A.M.H.; Batiuk, M.Y.; Prakash, K.; et al. Astrocyte layers in the mammalian cerebral cortex revealed by a single-cell in situ transcriptomic map. Nat. Neurosci. 2020, in press. [Google Scholar] [CrossRef]
- Doyle, J.P.; Dougherty, J.D.; Heiman, M.; Schmidt, E.F.; Stevens, T.R.; Ma, G.; Bupp, S.; Shrestha, P.; Shah, R.D.; Doughty, M.L.; et al. Application of a translational profiling approach for the comparative analysis of CNS cell types. Cell 2008, 135, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Boisvert, M.M.; Erikson, G.A.; Shokhirev, M.N.; Allen, N.J. The aging astrocyte transcriptome from multiple regions of the mouse brain. Cell Rep. 2018, 22, 269–285. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; McConnell, E.; Pare, J.F.; Xu, Q.; Chen, M.; Peng, W.; Lovatt, D.; Han, X.; Smith, Y.; Nedergaard, M. Glutamate-dependent neuroglial calcium signaling differs between young and adult brain. Science 2013, 339, 197–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusnakova, V.; Honsa, P.; Dzamba, D.; Ståhlberg, A.; Kubista, M.; Anderova, M. Heterogeneity of astrocytes: From development to injury-single cell gene expression. PLoS ONE 2013, 8, e69734. [Google Scholar] [CrossRef] [PubMed]
- Orre, M.; Kamphuis, W.; Osborn, L.M.; Melief, J.; Kooijman, L.; Huitinga, I.; Klooster, J.; Bossers, K.; Hol, E.M. Acute isolation and transcriptome characterization of cortical astrocytes and microglia from young and aged mice. Neurobiol. Aging 2014, 35, 1–14. [Google Scholar] [CrossRef]
- Clarke, L.E.; Liddelow, S.A.; Chakraborty, C.; Münch, A.E.; Heiman, M.; Barres, B.A. Normal aging induces A1-like astrocyte reactivity. Proc. Natl. Acad. Sci. USA 2018, 115, E1896–E1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soreq, L.; Consortium, U.K.B.E.; North American Brain Expression, C.; Rose, J.; Soreq, E.; Hardy, J.; Trabzuni, D.; Cookson, M.R.; Smith, C.; Ryten, M.; et al. Major shifts in glial regional identity are a transcriptional hallmark of human brain aging. Cell Rep. 2017, 18, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Morel, L.; Chiang, M.S.R.; Higashimori, H.; Shoneye, T.; Iyer, L.K.; Yelick, J.; Tai, A.; Yang, Y. Molecular and functional properties of regional astrocytes in the adult brain. J. Neurosci. 2017, 37, 8706–8717. [Google Scholar] [CrossRef] [Green Version]
- Kucukdereli, H.; Allen, N.J.; Lee, A.T.; Feng, A.; Ozlu, M.I.; Conatser, L.M.; Chakraborty, C.; Workman, G.; Weaver, M.; Sage, E.H.; et al. Control of excitatory CNS synaptogenesis by astrocyte-secreted proteins Hevin and SPARC. Proc. Natl. Acad. Sci. USA 2011, 108, E440–E449. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.J.; Philips, T.; Kim, N.; Dastgheyb, R.; Chen, Z.; Hsieh, Y.C.; Daigle, J.G.; Datta, M.; Chew, J.; Vidensky, S.; et al. Molecularly defined cortical astroglia subpopulation modulates neurons via secretion of Norrin. Nat. Neurosci. 2019, 22, 741–752. [Google Scholar] [CrossRef]
- Morel, L.; Men, Y.; Chiang, M.S.R.; Tian, Y.; Jin, S.; Yelick, J.; Higashimori, H.; Yang, Y. Intracortical astrocyte subpopulations defined by astrocyte reporter mice in the adult brain. Glia 2019, 67, 171–181. [Google Scholar] [CrossRef] [Green Version]
- John Lin, C.C.; Yu, K.; Hatcher, A.; Huang, T.W.; Lee, H.K.; Carlson, J.; Weston, M.C.; Chen, F.; Zhang, Y.; Zhu, W.; et al. Identification of diverse astrocyte populations and their malignant analogs. Nat. Neurosci. 2017, 20, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Gokce, O.; Stanley, G.M.; Treutlein, B.; Neff, N.F.; Camp, J.G.; Malenka, R.C.; Rothwell, P.E.; Fuccillo, M.V.; Südhof, T.C.; Quake, S.R. Cellular taxonomy of the mouse striatum as revealed by single-cell RNA-seq. Cell Rep. 2016, 16, 1126–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisel, A.; Muñoz-Manchado, A.B.; Codeluppi, S.; Lönnerberg, P.; La Manno, G.; Juréus, A.; Marques, S.; Munguba, H.; He, L.; Betsholtz, C.; et al. Brain structure. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 2015, 347, 1138–1142. [Google Scholar] [CrossRef]
- Batiuk, M.Y.; Martirosyan, A.; Wahis, J.; de Vin, F.; Marneffe, C.; Kusserow, C.; Koeppen, J.; Viana, J.F.; Oliveira, J.F.; Voet, T.; et al. Identification of region-specific astrocyte subtypes at single cell resolution. Nat. Commun. 2020, 11, 1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molofsky, A.V.; Kelley, K.W.; Tsai, H.H.; Redmond, S.A.; Chang, S.M.; Madireddy, L.; Chan, J.R.; Baranzini, S.E.; Ullian, E.M.; Rowitch, D.H. Astrocyte-encoded positional cues maintain sensorimotor circuit integrity. Nature 2014, 509, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Wang, W.; Kimelberg, H.K.; Zhou, M. Electrical coupling of astrocytes in rat hippocampal slices under physiological and simulated ischemic conditions. Glia 2010, 58, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Amzica, F.; Massimini, M. Glial and neuronal interactions during slow wave and paroxysmal activities in the neocortex. Cereb. Cortex 2002, 12, 1101–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rash, J.E.; Yasumura, T.; Dudek, F.E.; Nagy, J.I. Cell-specific expression of connexins and evidence of restricted gap junctional coupling between glial cells and between neurons. J. Neurosci. 2001, 21, 1983–2000. [Google Scholar] [CrossRef] [Green Version]
- Blomstrand, F.; Aberg, N.D.; Eriksson, P.S.; Hansson, E.; Rönnbäck, L. Extent of intercellular calcium wave propagation is related to gap junction permeability and level of connexin-43 expression in astrocytes in primary cultures from four brain regions. Neuroscience 1999, 92, 255–265. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, W.T.; Cornell-Bell, A.H.; Sontheimer, H. Astrocytes exhibit regional specificity in gap-junction coupling. Glia 1994, 11, 315–325. [Google Scholar] [CrossRef]
- Cotrina, M.L.; Gao, Q.; Lin, J.H.; Nedergaard, M. Expression and function of astrocytic gap junctions in aging. Brain Res. 2001, 901, 55–61. [Google Scholar] [CrossRef]
- Houades, V.; Koulakoff, A.; Ezan, P.; Seif, I.; Giaume, C. Gap junction-mediated astrocytic networks in the mouse barrel cortex. J. Neurosci. 2008, 28, 5207–5217. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Buckalew, R.; Du, Y.; Kiyoshi, C.M.; Alford, C.C.; Wang, W.; McTigue, D.M.; Enyeart, J.J.; Terman, D.; Zhou, M. Gap junction coupling confers isopotentiality on astrocyte syncytium. Glia 2016, 64, 214–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottrell, G.T.; Burt, J.M. Functional consequences of heterogeneous gap junction channel formation and its influence in health and disease. Biochim. Biophys. Acta 2005, 1711, 126–141. [Google Scholar] [CrossRef] [Green Version]
- Pannasch, U.; Freche, D.; Dallérac, G.; Ghézali, G.; Escartin, C.; Ezan, P.; Cohen-Salmon, M.; Benchenane, K.; Abudara, V.; Dufour, A.; et al. Connexin 30 sets synaptic strength by controlling astroglial synapse invasion. Nat. Neurosci. 2014, 17, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Ghézali, G.; Vasile, F.; Curry, N.; Fantham, M.; Cheung, G.; Ezan, P.; Cohen-Salmon, M.; Kaminski, C.; Rouach, N. Neuronal activity drives astroglial Connexin 30 in perisynaptic processes and shapes its functions. Cereb. Cortex 2019. [Google Scholar] [CrossRef] [PubMed]
- Rusakov, D.A. Disentangling calcium-driven astrocyte physiology. Nat. Rev. Neurosci. 2015, 16, 226–233. [Google Scholar] [CrossRef]
- Losi, G.; Mariotti, L.; Sessolo, M.; Carmignoto, G. New tools to study astrocyte Ca2+ signal dynamics in brain networks in vivo. Front. Cell Neurosci. 2017, 11, 134. [Google Scholar] [CrossRef] [Green Version]
- Takata, N.; Hirase, H. Cortical layer 1 and layer 2/3 astrocytes exhibit distinct calcium dynamics in vivo. PLoS ONE 2008, 3, e2525. [Google Scholar] [CrossRef] [Green Version]
- Shigetomi, E.; Bushong, E.A.; Haustein, M.D.; Tong, X.; Jackson-Weaver, O.; Kracun, S.; Xu, J.; Sofroniew, M.V.; Ellisman, M.H.; Khakh, B.S. Imaging calcium microdomains within entire astrocyte territories and endfeet with GCaMPs expressed using adeno-associated viruses. J. Gen. Physiol. 2013, 141, 633–647. [Google Scholar] [CrossRef]
- Shigetomi, E.; Kracun, S.; Sofroniew, M.V.; Khakh, B.S. A genetically targeted optical sensor to monitor calcium signals in astrocyte processes. Nat. Neurosci. 2010, 13, 759–766. [Google Scholar] [CrossRef] [Green Version]
- Haustein, M.D.; Kracun, S.; Lu, X.H.; Shih, T.; Jackson-Weaver, O.; Tong, X.; Xu, J.; Yang, X.W.; O’Dell, T.J.; Marvin, J.S.; et al. Conditions and constraints for astrocyte calcium signaling in the hippocampal mossy fiber pathway. Neuron 2014, 82, 413–429. [Google Scholar] [CrossRef] [Green Version]
- Slezak, M.; Kandler, S.; Van Veldhoven, P.P.; Van den Haute, C.; Bonin, V.; Holt, M.G. Distinct mechanisms for visual and motor-related astrocyte responses in mouse visual cortex. Curr. Biol. 2019, 29, 3120–3127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigetomi, E.; Tong, X.; Kwan, K.Y.; Corey, D.P.; Khakh, B.S. TRPA1 channels regulate astrocyte resting calcium and inhibitory synapse efficacy through GAT-3. Nat. Neurosci. 2011, 15, 70–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, K.M.; Hill-Eubanks, D.C.; Liedtke, W.B.; Nelson, M.T. TRPV4 channels stimulate Ca2+-induced Ca2+ release in astrocytic endfeet and amplify neurovascular coupling responses. Proc. Natl. Acad. Sci. USA 2013, 110, 6157–6162. [Google Scholar] [CrossRef] [Green Version]
- Straub, S.V.; Bonev, A.D.; Wilkerson, M.K.; Nelson, M.T. Dynamic inositol trisphosphate-mediated calcium signals within astrocytic endfeet underlie vasodilation of cerebral arterioles. J. Gen. Physiol. 2006, 128, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Wu, P.H.; Hughes, E.G.; Fukaya, M.; Tischfield, M.A.; Langseth, A.J.; Wirtz, D.; Bergles, D.E. Transient opening of the mitochondrial permeability transition pore induces microdomain calcium transients in astrocyte processes. Neuron 2017, 93, 587–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennings, A.; Tyurikova, O.; Bard, L.; Zheng, K.; Semyanov, A.; Henneberger, C.; Rusakov, D.A. Dopamine elevates and lowers astroglial Ca2+ through distinct pathways depending on local synaptic circuitry. Glia 2017, 65, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Bard, L.; Reynolds, J.P.; King, C.; Jensen, T.P.; Gourine, A.V.; Rusakov, D.A. Time-resolved imaging reveals heterogeneous landscapes of nanomolar Ca2+ in neurons and astroglia. Neuron 2015, 88, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Schwaller, B. Cytosolic Ca2+ buffers. Cold Spring Harb. Perspect. Biol. 2010, 2, a004051. [Google Scholar] [CrossRef]
- Wang, Y.; DelRosso, N.V.; Vaidyanathan, T.V.; Cahill, M.K.; Reitman, M.E.; Pittolo, S.; Mi, X.; Yu, G.; Poskanzer, K.E. Accurate quantification of astrocyte and neurotransmitter fluorescence dynamics for single-cell and population-level physiology. Nat. Neurosci. 2019. [Google Scholar] [CrossRef]
- Paukert, M.; Agarwal, A.; Cha, J.; Doze, V.A.; Kang, J.U.; Bergles, D.E. Norepinephrine controls astroglial responsiveness to local circuit activity. Neuron 2014, 82, 1263–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsu, Y.; Couchman, K.; Lyons, D.G.; Collot, M.; Agarwal, A.; Mallet, J.M.; Pfrieger, F.W.; Bergles, D.E.; Charpak, S. Calcium dynamics in astrocyte processes during neurovascular coupling. Nat. Neurosci. 2015, 18, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Vardjan, N.; Chowdhury, H.H.; Horvat, A.; Velebit, J.; Malnar, M.; Muhič, M.; Kreft, M.; Krivec, Š.G.; Bobnar, S.T.; Miš, K.; et al. Enhancement of astroglial aerobic glycolysis by extracellular lactate-mediated increase in cAMP. Front. Mol. Neurosci. 2018, 11, 148. [Google Scholar] [CrossRef] [PubMed]
- Mächler, P.; Wyss, M.T.; Elsayed, M.; Stobart, J.; Gutierrez, R.; von Faber-Castell, A.; Kaelin, V.; Zuend, M.; San Martín, A.; Romero-Gómez, I.; et al. In vivo evidence for a lactate gradient from astrocytes to neurons. Cell Metab. 2016, 23, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Robberecht, W.; Philips, T. The changing scene of amyotrophic lateral sclerosis. Nat. Rev. Neurosci. 2013, 14, 248–264. [Google Scholar] [CrossRef]
- Brancaccio, M.; Edwards, M.D.; Patton, A.P.; Smyllie, N.J.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Cell-autonomous clock of astrocytes drives circadian behavior in mammals. Science 2019, 363, 187–192. [Google Scholar] [CrossRef]
- Gourine, A.V.; Kasymov, V.; Marina, N.; Tang, F.; Figueiredo, M.F.; Lane, S.; Teschemacher, A.G.; Spyer, K.M.; Deisseroth, K.; Kasparov, S. Astrocytes control breathing through pH-dependent release of ATP. Science 2010, 329, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Morquette, P.; Verdier, D.; Kadala, A.; Féthière, J.; Philippe, A.G.; Robitaille, R.; Kolta, A. An astrocyte-dependent mechanism for neuronal rhythmogenesis. Nat. Neurosci. 2015, 18, 844–854. [Google Scholar] [CrossRef]
- Garcia-Cáceres, C.; Quarta, C.; Varela, L.; Gao, Y.; Gruber, T.; Legutko, B.; Jastroch, M.; Johansson, P.; Ninkovic, J.; Yi, C.X.; et al. Astrocytic insulin signaling couples brain glucose uptake with nutrient availability. Cell 2016, 166, 867–880. [Google Scholar] [CrossRef] [Green Version]
- Petrelli, F.; Dallérac, G.; Pucci, L.; Calì, C.; Zehnder, T.; Sultan, S.; Lecca, S.; Chicca, A.; Ivanov, A.; Asensio, C.S.; et al. Dysfunction of homeostatic control of dopamine by astrocytes in the developing prefrontal cortex leads to cognitive impairments. Mol. Psychiatry 2018. [Google Scholar] [CrossRef] [Green Version]
- Adamsky, A.; Kol, A.; Kreisel, T.; Doron, A.; Ozeri-Engelhard, N.; Melcer, T.; Refaeli, R.; Horn, H.; Regev, L.; Groysman, M.; et al. Astrocytic activation generates de novo neuronal potentiation and memory enhancement. Cell 2018, 174, 59–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, R.; Bajo-Grañeras, R.; Moratalla, R.; Perea, G.; Araque, A. Circuit-specific signaling in astrocyte-neuron networks in basal ganglia pathways. Science 2015, 349, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckervordersandforth, R.; Tripathi, P.; Ninkovic, J.; Bayam, E.; Lepier, A.; Stempfhuber, B.; Kirchhoff, F.; Hirrlinger, J.; Haslinger, A.; Lie, D.C.; et al. In vivo fate mapping and expression analysis reveals molecular hallmarks of prospectively isolated adult neural stem cells. Cell Stem Cell 2010, 7, 744–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messing, A.; Brenner, M.; Feany, M.B.; Nedergaard, M.; Goldman, J.E. Alexander disease. J. Neurosci. 2012, 32, 5017–5023. [Google Scholar] [CrossRef] [PubMed]
- Perez-Nievas, B.G.; Serrano-Pozo, A. Deciphering the astrocyte reaction in Alzheimer’s disease. Front. Aging Neurosci. 2018, 10, 114. [Google Scholar] [CrossRef] [Green Version]
- Takano, T.; Oberheim, N.; Cotrina, M.L.; Nedergaard, M. Astrocytes and ischemic injury. Stroke 2009, 40, S8–S12. [Google Scholar] [CrossRef] [Green Version]
- Brenner, M.; Johnson, A.B.; Boespflug-Tanguy, O.; Rodriguez, D.; Goldman, J.E.; Messing, A. Mutations in GFAP, encoding glial fibrillary acidic protein, are associated with Alexander disease. Nat. Genet. 2001, 27, 117–120. [Google Scholar] [CrossRef]
- Messing, A.; Goldman, J.E.; Johnson, A.B.; Brenner, M. Alexander disease: New insights from genetics. J. Neuropathol. Exp. Neurol. 2001, 60, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Lundgaard, I.; Osório, M.J.; Kress, B.T.; Sanggaard, S.; Nedergaard, M. White matter astrocytes in health and disease. Neuroscience 2014, 276, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Jing, L.; Lefebvre, J.L.; Gordon, L.R.; Granato, M. Wnt signals organize synaptic prepattern and axon guidance through the zebrafish unplugged/MuSK receptor. Neuron 2009, 61, 721–733. [Google Scholar] [CrossRef] [Green Version]
- He, C.W.; Liao, C.P.; Pan, C.L. Wnt signalling in the development of axon, dendrites and synapses. Open Biol. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Berger, W.; van de Pol, D.; Bächner, D.; Oerlemans, F.; Winkens, H.; Hameister, H.; Wieringa, B.; Hendriks, W.; Ropers, H.H. An animal model for Norrie disease (ND): Gene targeting of the mouse ND gene. Hum. Mol. Genet. 1996, 5, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, K.B. NDP-Related Retinopathies. In GeneReviews; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Stephens, K., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Fiala, J.C.; Spacek, J.; Harris, K.M. Dendritic spine pathology: Cause or consequence of neurological disorders? Brain Res. Brain Res. Rev. 2002, 39, 29–54. [Google Scholar] [CrossRef]
- Nijssen, J.; Comley, L.H.; Hedlund, E. Motor neuron vulnerability and resistance in amyotrophic lateral sclerosis. Acta Neuropathol. 2017, 133, 863–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bento-Abreu, A.; Van Damme, P.; Van Den Bosch, L.; Robberecht, W. The neurobiology of amyotrophic lateral sclerosis. Eur. J. Neurosci. 2010, 31, 2247–2265. [Google Scholar] [CrossRef] [PubMed]
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.A.; Burda, J.E.; Ren, Y.; Ao, Y.; O’Shea, T.M.; Kawaguchi, R.; Coppola, G.; Khakh, B.S.; Deming, T.J.; Sofroniew, M.V. Astrocyte scar formation aids central nervous system axon regeneration. Nature 2016, 532, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Houle, J.D.; Jin, Y. Chronically injured supraspinal neurons exhibit only modest axonal dieback in response to a cervical hemisection lesion. Exp. Neurol. 2001, 169, 208–217. [Google Scholar] [CrossRef]
- Stone, D.J.; Rozovsky, I.; Morgan, T.E.; Anderson, C.P.; Finch, C.E. Increased synaptic sprouting in response to estrogen via an apolipoprotein E-dependent mechanism: Implications for Alzheimer’s disease. J. Neurosci. 1998, 18, 3180–3185. [Google Scholar] [CrossRef] [Green Version]
- Jo, S.; Yarishkin, O.; Hwang, Y.J.; Chun, Y.E.; Park, M.; Woo, D.H.; Bae, J.Y.; Kim, T.; Lee, J.; Chun, H.; et al. GABA from reactive astrocytes impairs memory in mouse models of Alzheimer’s disease. Nat. Med. 2014, 20, 886–896. [Google Scholar] [CrossRef]
- Carter, S.F.; Schöll, M.; Almkvist, O.; Wall, A.; Engler, H.; Längström, B.; Nordberg, A. Evidence for astrocytosis in prodromal Alzheimer disease provided by 11C-deuterium-L-deprenyl: A multitracer PET paradigm combining 11C-Pittsburgh compound B and 18F-FDG. J. Nucl. Med. 2012, 53, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic analysis of reactive astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, N.; Itoh, Y.; Tassoni, A.; Ren, E.; Kaito, M.; Ohno, A.; Ao, Y.; Farkhondeh, V.; Johnsonbaugh, H.; Burda, J.; et al. Cell-specific and region-specific transcriptomics in the multiple sclerosis model: Focus on astrocytes. Proc. Natl. Acad. Sci. USA 2018, 115, E302–E309. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Castro, B.; Gangwani, M.R.; Yu, X.; Coppola, G.; Khakh, B.S. Astrocyte molecular signatures in Huntington’s disease. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatami, A.; Albay, R., 3rd; Monjazeb, S.; Milton, S.; Glabe, C. Monoclonal antibodies against Abeta42 fibrils distinguish multiple aggregation state polymorphisms in vitro and in Alzheimer disease brain. J. Biol. Chem. 2014, 289, 32131–32143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, H.M.; Mulder, S.D.; Beliën, J.A.; Musters, R.J.; Eikelenboom, P.; Veerhuis, R. Astrocytic Abeta1-42 uptake is determined by Abeta-aggregation state and the presence of amyloid-associated proteins. Glia 2010, 58, 1235–1246. [Google Scholar] [CrossRef]
- Karikari, T.K.; Nagel, D.A.; Grainger, A.; Clarke-Bland, C.; Crowe, J.; Hill, E.J.; Moffat, K.G. Distinct conformations, aggregation and cellular internalization of different tau strains. Front. Cell Neurosci. 2019, 13, 296. [Google Scholar] [CrossRef]
- Chavarría, C.; Rodríguez-Bottero, S.; Quijano, C.; Cassina, P.; Souza, J.M. Impact of monomeric, oligomeric and fibrillar alpha-synuclein on astrocyte reactivity and toxicity to neurons. Biochem. J. 2018, 475, 3153–3169. [Google Scholar] [CrossRef]
- Jackson, W.S. Selective vulnerability to neurodegenerative disease: The curious case of Prion Protein. Dis. Model. Mech. 2014, 7, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, B.C.; Karimy, J.K.; Kundishora, A.J.; Mestre, H.; Cerci, H.M.; Matouk, C.; Alper, S.L.; Lundgaard, I.; Nedergaard, M.; Kahle, K.T. Glymphatic system impairment in Alzheimer’s disease and idiopathic normal pressure hydrocephalus. Trends Mol. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kress, B.T.; Iliff, J.J.; Xia, M.; Wang, M.; Wei, H.S.; Zeppenfeld, D.; Xie, L.; Kang, H.; Xu, Q.; Liew, J.A.; et al. Impairment of paravascular clearance pathways in the aging brain. Ann. Neurol. 2014, 76, 845–861. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xiao, N.; Chen, Y.; Huang, H.; Marshall, C.; Gao, J.; Cai, Z.; Wu, T.; Hu, G.; Xiao, M. Deletion of aquaporin-4 in APP/PS1 mice exacerbates brain Abeta accumulation and memory deficits. Mol. Neurodegener. 2015, 10, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliff, J.J.; Chen, M.J.; Plog, B.A.; Zeppenfeld, D.M.; Soltero, M.; Yang, L.; Singh, I.; Deane, R.; Nedergaard, M. Impairment of glymphatic pathway function promotes tau pathology after traumatic brain injury. J. Neurosci. 2014, 34, 16180–16193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Ding, F.; Deng, S.; Guo, X.; Wang, W.; Iliff, J.J.; Nedergaard, M. Focal solute trapping and global glymphatic pathway impairment in a murine model of multiple microinfarcts. J. Neurosci. 2017, 37, 2870–2877. [Google Scholar] [CrossRef]
- Magnusson, J.P.; Göritz, C.; Tatarishvili, J.; Dias, D.O.; Smith, E.M.; Lindvall, O.; Kokaia, Z.; Frisén, J. A latent neurogenic program in astrocytes regulated by Notch signaling in the mouse. Science 2014, 346, 237–241. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, L.; Wu, Z.; Chen, Y.; Wang, F.; Chen, G. In vivo direct reprogramming of reactive glial cells into functional neurons after brain injury and in an Alzheimer’s disease model. Cell Stem Cell 2014, 14, 188–202. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.H.; Li, W.; Zheng, J.J.; Xu, Y.G.; He, Q.; Chen, G. Differential neuronal reprogramming induced by NeuroD1 from astrocytes in grey matter versus white matter. Neural Regen. Res. 2020, 15, 342–351. [Google Scholar] [CrossRef]
- Kelley, K.W.; Nakao-Inoue, H.; Molofsky, A.V.; Oldham, M.C. Variation among intact tissue samples reveals the core transcriptional features of human CNS cell classes. Nat. Neurosci. 2018, 21, 1171–1184. [Google Scholar] [CrossRef]
Figure 1.
Astrocyte heterogeneity in the central nervous system (CNS). Different populations of astrocytes have been identified in the grey and white matter of the rodent brain and spinal cord, based on differences in morphology and marker protein expression. Astrocyte heterogeneity may arise in two separate ways. During development, astrocytes are generated from distinct pools of progenitor cells, which express unique combinations of transcription factors that drive cell identity (intrinsic). Alternatively, astrocyte diversity may be driven by cues from neighboring cells (such as sonic hedgehog release) (extrinsic) in adult tissue. In reality, it is likely that a combination of intrinsic and extrinsic factors drives astrocyte heterogeneity (see the main text for details).
Figure 1.
Astrocyte heterogeneity in the central nervous system (CNS). Different populations of astrocytes have been identified in the grey and white matter of the rodent brain and spinal cord, based on differences in morphology and marker protein expression. Astrocyte heterogeneity may arise in two separate ways. During development, astrocytes are generated from distinct pools of progenitor cells, which express unique combinations of transcription factors that drive cell identity (intrinsic). Alternatively, astrocyte diversity may be driven by cues from neighboring cells (such as sonic hedgehog release) (extrinsic) in adult tissue. In reality, it is likely that a combination of intrinsic and extrinsic factors drives astrocyte heterogeneity (see the main text for details).

Figure 2.
Transcriptome impacts function: the consequences of astrocyte heterogeneity. Differences in transcript expression between astrocytes can affect the function of local neuronal circuits, ultimately leading to functional impairments. For example, astrocytes in the ventral horn of the spinal cord are enriched in expression of Kcnj10, which encodes the K+ channel Kir4.1. Astrocyte-specific deletion of Kir4.1 leads to impaired local K+ homeostasis, a selective decrease in fast α-motor neuron (MN) size and impaired function, a decrease in the size of the innervated muscle and consequent loss of peak strength. Figure is adapted from Kelly et al. [47].
Figure 2.
Transcriptome impacts function: the consequences of astrocyte heterogeneity. Differences in transcript expression between astrocytes can affect the function of local neuronal circuits, ultimately leading to functional impairments. For example, astrocytes in the ventral horn of the spinal cord are enriched in expression of Kcnj10, which encodes the K+ channel Kir4.1. Astrocyte-specific deletion of Kir4.1 leads to impaired local K+ homeostasis, a selective decrease in fast α-motor neuron (MN) size and impaired function, a decrease in the size of the innervated muscle and consequent loss of peak strength. Figure is adapted from Kelly et al. [47].

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pestana, F.; Edwards-Faret, G.; Belgard, T.G.; Martirosyan, A.; Holt, M.G. No Longer Underappreciated: The Emerging Concept of Astrocyte Heterogeneity in Neuroscience. Brain Sci. 2020, 10, 168. https://doi.org/10.3390/brainsci10030168
AMA Style
Pestana F, Edwards-Faret G, Belgard TG, Martirosyan A, Holt MG. No Longer Underappreciated: The Emerging Concept of Astrocyte Heterogeneity in Neuroscience. Brain Sciences. 2020; 10(3):168. https://doi.org/10.3390/brainsci10030168
Chicago/Turabian StylePestana, Francisco, Gabriela Edwards-Faret, T. Grant Belgard, Araks Martirosyan, and Matthew G. Holt. 2020. "No Longer Underappreciated: The Emerging Concept of Astrocyte Heterogeneity in Neuroscience" Brain Sciences 10, no. 3: 168. https://doi.org/10.3390/brainsci10030168
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.